
Abstract
Abstract
The effects of partial root-zone drying (PRD) on tomato fruit growth and proteome in the pericarp of cultivar Ailsa Craig were investigated. The PRD treatment was 70% of water applied to fully irrigated (FI) plants. PRD reduced the fruit number and slightly increased the fruit diameter, whereas the total fruit fresh weight (FW) and dry weight (DW) per plant did not change. Although the growth rate was higher in FI than in PRD fruits, the longer period of cell expansion resulted in bigger PRD fruits. Proteins were extracted from pericarp tissue at two fruit growth stages (15 and 30 days post-anthesis [dpa]), and submitted to proteomic analysis including two-dimensional gel electrophoresis and mass spectrometry for identification. Proteins related to carbon and amino acid metabolism indicated that slower metabolic flux in PRD fruits may be the cause of a slower growth rate compared to FI fruits. The increase in expression of the proteins related to cell wall, energy, and stress defense could allow PRD fruits to increase the duration of fruit growth compared to FI fruits. Upregulation of some of the antioxidative enzymes during the cell expansion phase of PRD fruits appears to be related to their role in protecting fruits against the mild stress induced by PRD.
Introduction
In many countries, as a consequence of global climate change and environmental pollution, water use in agriculture is reduced. Water supplies are also under pressure from non-agricultural users, so that saving water resources and increasing agricultural productivity per unit of water (“more crop per drop”) are becoming of strategic importance for many countries. Considerable emphasis is placed on crop physiology and crop management under dry conditions with the aim of increasing crop water use efficiency (WUE; Costa et al., 2007).
Partial root-zone drying technique (PRD) is one of the irrigation methods that have been suggested to increase WUE of many agricultural crops (FAO, 2002). Under PRD, only half of the root zone is irrigated, while the other half is allowed to dry out. The treatment is then reversed, allowing the previously well-watered side of the root system to dry down, while fully irrigating the previously dry side. The principle behind PRD is that irrigating part of the root system keeps the leaves hydrated, although exposing the remaining part of the roots to soil drying triggers synthesis and transport of chemical signals, particularly the plant hormone abscisic acid (ABA), from roots to the shoots via the xylem (Loveys et al., 2000). The frequency of the switch is determined by the soil type, genotypes, or other factors such as rainfall and temperature. In most published data, the PRD cycle runs 10–15 days (Stikić et al., 2010).
Many studies have demonstrated significant and beneficial effects of PRD on increasing WUE and conserving water for irrigation. However, there are discrepancies in the published results concerning the feasibility of these techniques to maintain yield. There are reports in the literature that demonstrate that under PRD, tomato fruit yield was maintained and improved in both processed tomatoes (Zegbe-Domínguez et al., 2003, 2004; Zegbe et al., 2006a, 2006b) and fresh market tomatoes (Kirda et al., 2004). On the contrary, there are also results showing a reduction in tomato yield of plants grown under PRD (Tahi et al., 2007; Topcu et al., 2007).
Apart from genotypic effects, tomato yield depends on fruit growth, especially on the growth of pericarp cells. According to Ho and Hewitt (1986), the development of pericarp tissue determines more than two-thirds of the potential fruit weight. After fruit set, tomato fruit development consists of three phases, including cell division, cell expansion, and ripening (Bertin, 2005; Gillaspy et al., 1993). When cell division ends, the final size of the fruit is determined by the rate and duration of cell expansion (Grange, 1995). The majority of tomato fruit growth by cell expansion occurs between 15 and 35 days after anthesis (Pearce et al., 1993), with some genotypic variations (Bertin, 2005).
Fruit growth and productivity are complex processes depending on the interaction of different biochemical and physiological processes with genetic and environmental factors. From previously-published data concerning the biochemical mechanisms controlling growth of tomato cells, it is evident that many cell wall enzymes have been associated with tomato fruit growth during the development towards maturation. Cell wall enzymes such as XET-xyloglucan-endotransglycosylase (Thompson et al., 1998), and expansins (Rose et al., 1997), have been shown to be closely associated with cell wall expansion and tomato fruit growth during development, whereas the appearance of different peroxidase isozymes and increases in peroxidase activity within the pericarp during maturation have been associated with cell wall stiffening and the cessation of growth before ripening (Andrews et al., 2000, 2001; Thompson et al., 1998). Recently, proteomics was used for the investigation of the biochemical mechanisms of tomato growth (Faurobert et al., 2007). Extensive proteome analyses done during tomato fruit development and ripening demonstrated changes in a large number of proteins during fruit development: 15 proteins associated with carbohydrate metabolism, 5 with photosynthesis and respiration, 9 with amino acid metabolism, 5 with secondary metabolism, and 1 for vitamin and lipid metabolism (Faurobert et al., 2007).
Surprisingly, despite the intensive research on PRD's effects on tomato growth and productivity, little work has been done regarding the biochemical mechanisms of tomato fruit growth and ripening under PRD. Very recently, Shu and associates (2010) obtained proteomic profiles of rice leaves from plants under partial root osmotic stress (PROS), whole root osmotic stress (WROS), and well-watered (WW) conditions. By their proteomic analyses, a total of 58 proteins showed differential expression after one or both treatments, and functional classification of these proteins suggests that stress signals regulate photosynthesis, and carbohydrate and energy metabolism. Two other proteins (anthranilate synthase and submergence-induced nickel-binding protein) were upregulated only in the PROS plants, indicating their important roles in stress resistance.
Our earlier results showed that increases in the activity of enzyme cell wall-associated peroxidase in tomato fruits under PRD coincided with the ripening phase (Savic et al., 2008). These data indicated that cell wall-associated peroxidase may promote tomato fruit maturation by inducing a more rapid process. They also emphasize the importance of increasing our understanding of the biochemical events involved in the response of tomato fruit to PRD. The aims of this study were to: (1) compare the fruit growth processes between PRD and fully irrigated (FI) plants in the period from 15–30 days post-anthesis (dpa), for which 15 dpa corresponds to the stage of full expansion and rapid fruit growth, whereas 30 dpa is close to the beginning of ripening processes; (2) identify the pericarp proteins that vary in these stages of cell expansion to examine their functions; and (3) link proteins with both fruit developmental stages and PRD and FI effects.
Materials and Methods
Plant material and growing conditions
Tomato plants (Solanum lycopersicum L.), cv. Ailsa Craig were raised from seed and transplanted into 20-L pots (one plant per pot) filled with 11 kg of commercial compost in a growth chamber (photoperiod was 14-h; light intensity at plant level was 300 μmolm−2s−1, with temperature 25/18°C, and relative humidity 70%). The pots were specially designed for PRD experiments, such that they were separated with plastic sheets into two equal-sized compartments. Washed roots of the seedlings were divided into approximate halves and repotted into these two hydraulically separated compartments. The compartments were classified as PRD-L (left side) and PRD-R (right side). After transplantation, all plants were irrigated daily to full pot-holding capacity, with a volumetric soil water content (θ) of 35%. The θ of both compartments of each pot was measured daily using TDR probes (time domain reflectometer, TRASE; Soilmoisture Equipment Corp., Santa Barbara, CA, USA). Ten days after transplantation, the plants were subjected to following irrigation treatments: (1) full irrigation (FI), in which the whole root system was irrigated daily at 9:00 h, to reach a field capacity around 35%; and (2) partial root-zone drying (PRD), in which the same amount of water as for FI was applied to one-half of the roots while the other half was allowed to dry. The irrigation from wet to dry side was shifted when θ of the dry side had decreased to 15–20%. In total PRD, the plants received about 70% of the water that was applied for irrigation of the FI plants.
Measurement of physiological parameters
The growth rate for the fruits from the first five flower trusses was calculated for the whole fruit growth period. This calculation was based on the results of daily measurements of the fruit's equatorial diameter, from the third flower trusses, with a digital caliper. The yield per plant was calculated by multiplying the average fresh weight (FW) of red ripe fruits by their total number. Then, for dry weight (DW) measurements, the fresh fruits were cut into halves and oven-dried at 85°C. Maturation of fruits was followed by visual daily estimation. At the end of the experiments all leaves were collected and measurements of their fresh and dry weights were done.
Fruit sampling and protein extraction for proteomic analysis
Fruit samples for proteome analyses were harvested from each of the first five flower trusses of a single plant. Samples for analyses were collected at two stages of tomato fruit development: 15 dpa, corresponding to the stage of full expansion and rapid growth of fruit cells, and 30 dpa, at the stage close to the beginning of the ripening process. Pericarps were collected from at least 15 different fruits, immediately frozen in liquid nitrogen, and stored at −80°C. Prior to protein extraction the samples were ground to a fine powder in pre-chilled steel cylinders in a mixer mill.
Protein extraction was done by a phenol method, as described by Faurobert and colleagues (2006). Before two-dimensional gel electrophoresis (2-DE), the proteins were solubilized in lysis buffer (9 M urea, 4% [w/v] CHAPS, 0.5% [v/v] Triton X-100, 20 mM dithiothreitol [DTT], and 1.2% [v/v] pharmalyte, pH 3–10). Protein concentration was measured according to a modified Bradford assay (Ramagli and Rodriguez, 1985).
2-DE
All procedures for separation of proteins by 2-DE, image analysis, and mass spectrometry, were done as previously described by Manaa and associates (2011). Proteins (500 μg) were first separated according to their pI value. After passive rehydration of 24-cm-long Immobiline dry strips (pH 4–7; GE Healthcare Bio-Sciences, Piscataway Township, NJ, USA), isoelectric focusing was performed with the electrophoresis power supply EPS 3500 XL (Pharmacia Biotech, Piscataway, NJ, USA). SDS-PAGE was carried out with 13% acrylamide gels in the Hoefer Isodalt electrophoresis chamber. Four gels per sample were stained with colloidal Coomassie blue (Neuhoff et al., 1988).
Protein spot detection and quantification were done using Progenesis SameSpot software version 3.0, which allows tracking differences in protein patterns between irrigation treatments. Based on statistical tests, spots were picked for mass spectrometry (MS) analysis.
Protein identification by MS
The varying protein spots (52 in number) from the gels were sliced, and trypsin-digested proteins were analyzed by liquid chromatography-mass spectrometry/mass spectrometry (LC-MS/MS). Identification was done using the Sol Genomics Network (SGN) tomato unigene database. High-performance liquid chromatography (HPLC) was performed with the Ultimate LC system combined with Famos autosampler and Switchos II microcolumn switching for pre-concentration (Dionex Corp., Sunnyvale, CA, USA). Identification was performed with Xtandem software. The main search parameters were trypsin digestion, cys carboxyamidomethylation and Met oxidation; protein N-ter acetylation and deamination were set to enzymatic cleavage, static or possible modifications. For interrogation the SGN tomato database (EST database db65, version 20100114, ftp.sgn.cornell.edu/unigene_builds//, 42257 sequences) was translated in the six reading frames.
Identified tryptic peptides were filtered according (1) to their cross-correlation score (Xcorr), superior to 1.7, 2.2, and 3.3 for mono-, di-, and tricharged peptides, respectively; and (2) to their probability lower then 0.05. A minimum of two different peptides was required. In the case of identification with only two or three MS/MS spectra, similarity between the experimental and the theoretical MS/MS spectra was visually confirmed.
Statistical analysis
The statistical analyses for all physiological parameters were performed with SigmaPlot software (version 11.0) using Student's t-test, with significance level less than 0.05. For protein quantification, Progenesis SameSpot software version 3.0 was used to detect varying spots using one-way analysis of variance (ANOVA) on normalized spot volume from the gel repeats with p<0.01 and q<0.015. In order to assess the effect of stage, treatment, and interaction, a two-factor ANOVA model was applied using R software version 2.11.1 (The R Project for Statistical Computing; http://www.r-project.org/), with significance levels of *p<0.01, **p<0.001, and ***p<0.0001.
Results
Soil water content
Changes in soil water content for FI and PRD treatments during the experimental period are shown in Figure 1. Soil water content values (θ) with FI treatment were kept close to the field capacity (35%), while θ in the PRD groups depended on whether the side was irrigated or not (Fig. 1). Withholding water from one compartment of the pots allowed the soil to dry and to reach a θ of 18% after 5–8 days. After the re-watering of the dry compartment, θ returned to about 35% within 3 days. At the same time the previously dry compartment was irrigated. This pattern of alternating soil drying and re-watering of the PRD compartments was sustained during the entire experimental period. In all, in the PRD treatment group there were 9 shifts between the dry- and wet-side watering.
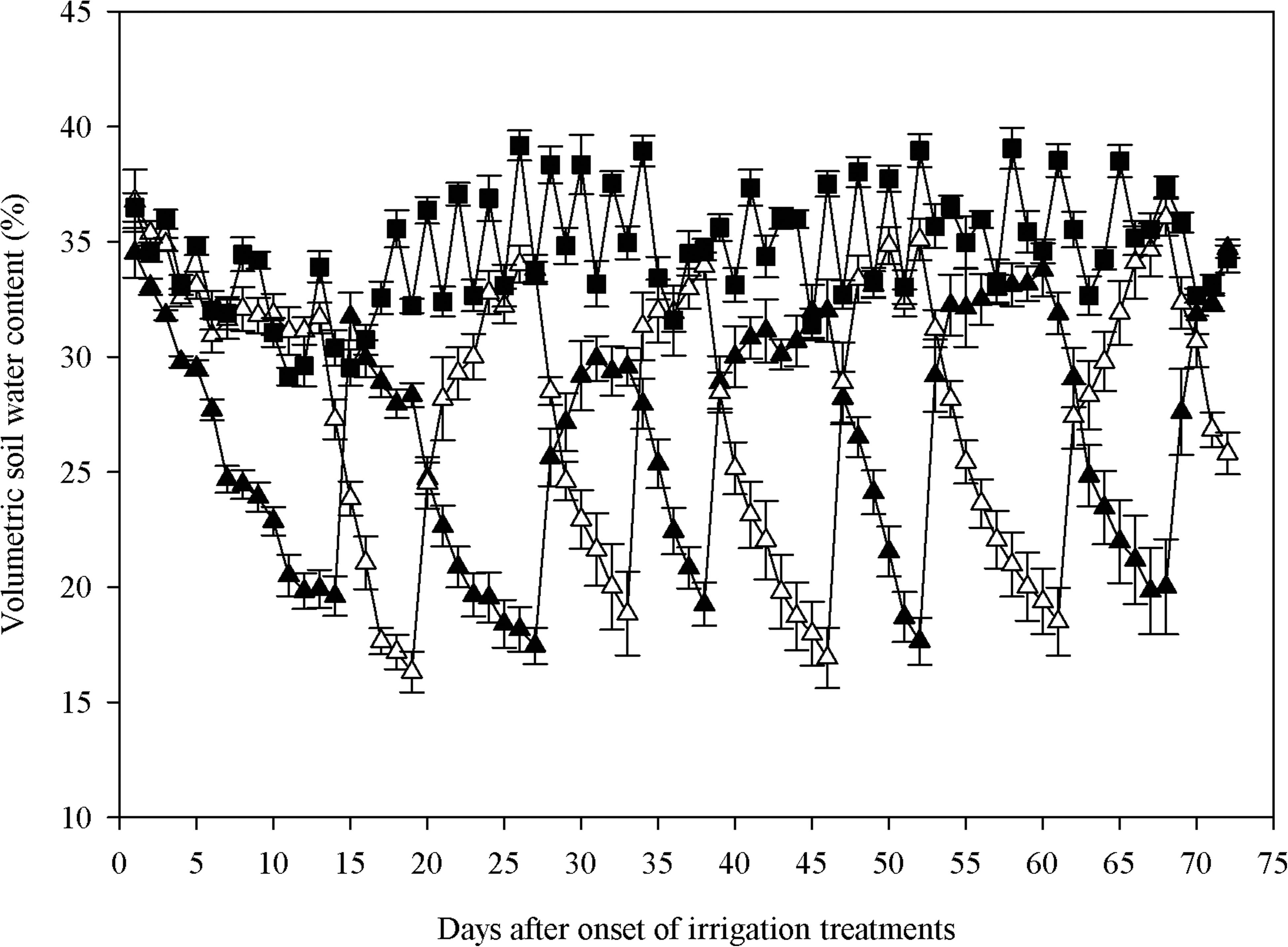
Changes in volumetric soil water content (%) for full irrigation (FI, solid squares) and partial root-zone drying (PRD-L, solid triangles and PRD-R, open triangles) treatments of tomato plants.
Plant growth and biomass
Compared to the FI treatment, the PRD treatment significantly reduced the number of fruits yielded per plant (20%), while the final fruit diameter was 4.3 mm bigger in PRD than in FI plants (Table 1). FW data showed that the PRD fruits were heavier (61.8 g) compared to FI fruits (50.1 g). Dry mass of the fruits was not affected by the treatments. Although the number of fruits was reduced under PRD, due to the higher fresh weight of the individual fruits, the yield was similar in both treatment groups (about 2.150 kg per plant). PRD had a reducing effect on leaf biomass. FW and DW results were significantly higher in FI compared to PRD plants (15% and 25% for FW and DW, respectively). Differences were also found in fruit:leaf ratios, and these values were higher in PRD than in FI plants. These data suggest that the increase of resource partitioning between leaves and fruits under PRD might influence the FW and DW of PRD fruits.
p≤0.05; **p≤0.01.
Means±standard error for at least six plants per treatment group are shown.
ns, not significant; FW, fresh weight; DW, dry weight; PRD, partial root-zone drying; FI, fully irrigated.
The kinetics of tomato fruit growth are presented in Figure 2, as the time course of fruit diameter changes (Fig. 2A), and fruit growth rate (FGR, Fig. 2B). Tomato fruit diameters were fitted by fourth-order regression, and the curves followed a sigmoid pattern of growth. The FGR bell-shaped curve was obtained by fitting the changes in FGR by the third-order regression line (Fig. 2B).

Kinetics of fruit growth under partial root-zone drying PRD (solid triangles) and fully irrigated (FI) (solid squares) treatments. (
The fruit diameter results (Fig. 2A) showed that in both PRD and FI treatments the intensive fruit growth rate started 7–10 dpa, but reached a plateau earlier in FI fruits (between 40 and 50 dpa) than in PRD fruits (between 50 and 60 dpa). During nearly the entire experimental period the fruit diameter was smaller in PRD fruits than in FI fruits. The FGR results (Fig. 2B) showed that the maximal growth rate was significantly higher in FI fruits (1.7 mm/day−1), than under PRD treatment (1.4 mm/day−1). However, because of the prolonged period of fruit growth in PRD than in FI plants (about 10 days), the final diameter of the fruits was slightly higher in PRD than in FI fruits.
Proteome maps of tomato pericarp proteins
Proteomic results were compared between two stages of pericarp fruit cell expansion (15 and 30 dpa), and between the two types of irrigation (PRD and FI). The comparison started with 2-DE gels. In total, 1679 spots were detected in fruit pericarp (Fig. 3). Among the identified spots, a total of 52 protein spots displayed differential abundance variations (p<0.05). These spots were then excised from the gels for MS and protein identification. In total, it was possible to identify 46 proteins from 52 protein spots. All the searches were done against the SGN database. For the 6 remaining spots, it was not possible to determine the protein spot identity. Two of the spots showed a different expression pattern (both up- and downregulation), although their identification confirmed that it was the same protein (enolase). Therefore, they were marked differently, as spots 9 and 33 (Table 2).
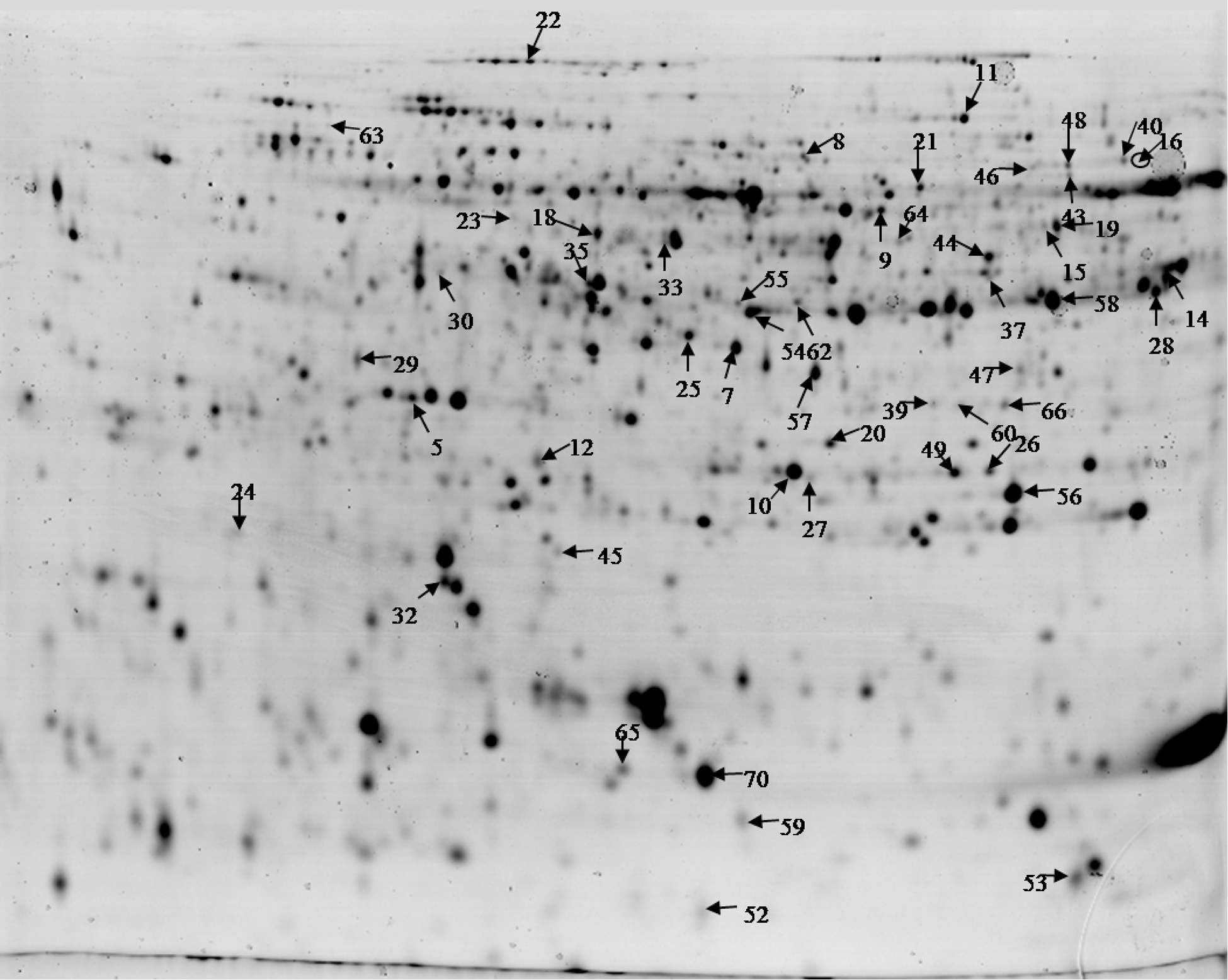
Two-dimensional electrophoresis gel of tomato pericarp proteins.
Spot number corresponds to the position of the spot in the gel shown in Figure 3.
Database of the Sol Genomic Network/UniProt accession numbers.
Peptide signal (PS) according to TargetP software: cTP, a chloroplast transit peptide; mTP, a mitochondrial targeting peptide; cyt, cytoplasm; SP, secreted protein.
Theoretical pI and mass (kDa) of the identified proteins.
Calculated pI and mass (kDa) of identified proteins by Progenesis Software.
ANOVA results for treatment, stage, and interaction effects: ns, non-significant effect; *p<0.01, **p<0.001, ***p<0.0001.
Normalized volume of the identified proteins: FI treatment/increased (+) or decreased (−) in average volume in PRD treatment.
ANOVA, analysis of variance; PRD, partial root-zone drying; FI, fully irrigated.
Distribution of proteins into functional classes
The identified proteins were classified into 8 functional categories to provide a comparison between developmental effects and the effects of PRD and FI on the investigated fruits. The categories that included protein spots were: carbon metabolism (13 proteins); amino acid metabolism (5 proteins); protein translation, processing, and degradation (8 proteins); energy metabolism (3 proteins); cell-wall-related (5 proteins); oxidative stress (8 proteins); stress defense and heat shock (4 proteins); and 6 proteins whose function was unknown. The list of identified proteins that were classified according to their function is presented in Table 2. They are shown with the protein spot number, the identification parameters, annotation and predicted localization, theoretical and calculated pI/Mr, treatment and stage effects, and their interaction by ANOVA testing.
Proteome changes mediated by stage effect
By proteomic analyses a total of 29 proteins showed different expression during fruit cell expansion at 15 dpa and 30 dpa in both the FI and PRD treatment groups (Table 2).
Specific expression of proteins under PRD showed stage effects, which were both up- and downregulated. Upregulated proteins included 9 proteins, and among them the highest number was in the oxidative stress category (4). Others were classified into the following categories: protein translation processing and degradation (2), carbon metabolism (1), cell wall (1), and stress defense and heat shock (1). The number of downregulated proteins was smaller and included only three proteins from the category of carbon metabolism.
In FI fruits only 3 proteins exhibited upregulation during the fruit cell expansion process, and they belonged to the carbon metabolism (1), oxidative stress (1), and stress defense and heat shock categories (1). At the same time only 1 protein (amino acid metabolism) was downregulated.
The results also showed that 12 proteins from different categories followed the same trends of expression during fruit growth in both the PRD and FI treatment groups. Among them, 8 proteins were upregulated, including 2 in the category of carbon metabolism, 2 in protein translation processing and degradation, 2 were cell-wall-related, and 2 were in the category of miscellaneous proteins. The expression of 4 proteins belonging to the categories of protein translation processing and degradation, and to cell wall and oxidative stress, were downregulated.
Proteome changes mediated by treatment effect
Comparison between the FI and PRD treatments (the treatment effects shown in Table 2) at 15 dpa showed differences for 31 proteins. The proteins upregulated under PRD compared to the same stage of FI fruits included 6 proteins from five categories: carbon metabolism (2), amino acid metabolism (1), energy metabolism (1), oxidative stress (1), and stress defense and heat shock (1).
At 15 dpa the number of proteins downregulated in PRD fruits was higher, and included 21 proteins from the following categories: carbon metabolism (6); amino acid metabolism (1); protein translation, processing, and degradation (4); energy metabolism (2); oxidative stress (5); and stress defense and heat shock (1). Additionally 2 proteins were found in the category of miscellaneous proteins.
In the phase close to the beginning of the ripening processes (30 dpa), 30 proteins showed different expression. The upregulation in PRD fruits compared to FI fruits was noted for only 3 proteins: 1 whose function was connected to energy metabolism, and 2 belonging to the stress defense and heat shock category. The number of downregulated proteins was higher (27) than of upregulated proteins. They belonged to six categories, including: carbon metabolism (10); amino acid metabolism (3); protein translation, processing, and degradation (5); oxidative stress (2); stress defense and heat shock (1); and miscellaneous proteins (4).
Our results also showed significant interaction effects between stage and treatment. This was noted for 9 proteins: 2 in carbon metabolism (spots 37 and 44); 3 in protein translation, processing, and degradation (spots 12, 15, and 30); two in the cell-wall-related category (spots 14 and 60); 1 from the oxidative stress category (spot 59); and 1 in miscellaneous proteins (spot 29). Therefore, the different expression levels of these proteins that were registered could not solely be a consequence of the stage effect, or of the applied treatment.
Discussion
Plant fruit growth and yield
The soil water content measurements displayed a characteristic pattern for the PRD experiments, clearly showing alternate soil drying and re-watering (Fig. 1). A similar pattern of soil water content and shifting intervals has also been observed in other PRD experiments (Sobeih et al., 2004; Wang et al., 2010).
The PRD irrigation must be switched regularly from one side of the roots to the other to keep roots in dry soil alive and fully functional, and to sustain the supply of root signals (Davies et al., 2000). Root signals can thus be transmitted to the shoots if there is sufficient sap flow (Dodd et al., 2008). However, in the literature there are many differences in applied irrigation scheduling and in the amounts of water applied to the wet side or dry side. These differences, as well as differences in soil heterogeneity (Dodd, 2007), may explain the conflicting results of PRD's effects on growth or yield of different plants, including the tomato.
Furthermore, there is a discrepancy in published results concerning the feasibility of PRD techniques to maintain tomato fruit growth or yield. Some reports in the literature demonstrate that with the use of PRD, the yield of tomato plants is maintained or improved for both processing tomatoes (Zegbe-Domínguez et al., 2003, 2004; Zegbe et al., 2006a, 2006b), and fresh market tomatoes (Kirda et al., 2004). On the contrary, there are also results showing a decrease in tomato yield of plants grown under PRD (Tahi et al., 2007; Topcu et al., 2007).
Our results show that plants under PRD produced a smaller number of the fruits than those grown under FI (Table 1). Although in our experiment total flower numbers and abortion/fruit setting success were not measured, visual estimation of the investigated plants indicated that a smaller number of flowers was produced in PRD than in FI plants. However, due to the higher fresh weight of individual PRD fruits compared to FI fruits, the yield was similar for both treatments. On the other hand, Mingo and associates (2003) showed that PRD treatment significantly limited the fruit growth rate in an experiment with the same cultivar. The discrepancy between our findings and those of Mingo's group may be explained by differences in PRD experiment duration. Mingo's group explored the short-term effects of PRD, which were assessed only 5 days after PRD and FI treatments. Our fruit growth and yield results were also inconsistent with the results of Davies and colleagues (2000), who used tomato cultivar Ailsa Craig in a similar a long-term PRD experimental system. They found that although the total number of fruits per plant was not significantly different between the FI and PRD groups, the fruit's fresh weight and total plant yield were significantly reduced with PRD treatment. Also, their results did not show any differences in fruit dry weight between treatments. However, it must be noted that their group applied 50% of FI irrigation to PRD plants, whereas in our experiment PRD plants received 70% of FI water irrigation. Our previous PRD results with irrigation scheduling similar to the 50% level for PRD used by the Davies group, which were done with another tomato cultivar (Sunpak), showed that under these conditions both fresh fruit weight and yields per plant were reduced (Savic et al., 2008).
The fruit DW and FW seen under PRD in our experiments could be explained by a source/sink relationship. Tomato fruit is the strongest sink for water, and assimilates it more than the rest of plant's organs, whereas leaves are the main source (Ho, 1992). We might speculate, as did Davies and associates (2000), that the reduction of leaf biomass caused by PRD resulted in a relative increase in the source/sink relationship. Consequently, assimilates and water were redirected from leaves towards the fruits.
In the literature there is much information concerning the final tomato fruit diameter, and the effects of different environmental factors, especially temperature, on both the cell division rate and the rate or duration of cell expansion (Adams et al., 2001; Bertin, 2005). However, the number of similar approaches concerning the effects of PRD on fruit growth is limited. In a previous study we compared cell growth rates and the duration of cell expansion in pericarp tissue of PRD- versus FI-irrigated fruits (Savic et al., 2008). The results demonstrated that the maximal growth rate was significantly higher in FI treatment compared to PRD, although the duration of cell expansion was longer in PRD than in FI fruits. As a consequence, no significant differences in fruit diameter between PRD and FI fruits were found. In our present experiments similar patterns of fruit diameter curves and of fruit growth rate curves were obtained (Fig. 2A and B). The first detectable differences in PRD and FI fruit diameter started to occur 7–10 dpa, corresponding to the beginning of the cell expansion phase (Monselise et al., 1978). During most of the expansion period, fruit growth rate data were higher in FI fruits than in PRD fruits, and maximal differences were seen between 25 and 30 dpa. The breaker stage (when most of the cells had stopped expanding) was reached 10 days later in PRD than in FI fruits. Finally, the mean diameter of PRD fruits was larger than that of FI fruits. These results are in accord with our previous results (Savic et al., 2008), and indicate that in the long-term, the duration of cell expansion is more important for obtaining larger fruits than maximal FGR values.
On the basis of the available data it is difficult to explain the differences between our PRD experiment and the experiments for which smaller total amounts of water were used for irrigating the PRD wet side. Although we did not measure the chemical signals generated by PRD plants, we might speculate that using more water in a PRD system leads to better detection of the soil water content, and increased synthesis and transport of these chemical signals. This assumption is in accord with that of Tardieu and Davies (1992), who suggested that a plant exposed to drought in the field “measures” the access that roots have to soil water, rather than soil water potential per se. Furthermore, Dodd and colleagues (2008) demonstrated that to sustain effective chemical signalling under PRD conditions, it is necessary not only to produce ABA in dry roots (as a key signal molecule), but also to maintain water uptake from the roots, sap flow, and signal transport to the shoots. Thus, larger soil water quantities in the wet part of the root system may maximize ABA export from the entire root system and transport to the shoots.
The potential size of tomato fruit also depends on the rate of water accumulation, because water may account for 95% of the total fresh weight (Ho, 1992). The results of Freitas and associates (2011) demonstrated higher fruit fresh weight of two tomato cultivars in response to ABA treatment. This was the result of the ABA-induced increase in the abundance of vascular bundles in tomato fruit. We might further speculate that both increased ABA accumulation under PRD, and longer growth periods of PRD fruits compared to FI fruits, may result in more accumulation of water and increased fresh weight in these fruits, and therefore increased yields.
PRD changed carbon and amino acid metabolism during fruit growth
With the aim of better understanding the effects of PRD on fruit growth, we focused our proteomic research on 15 dpa, which corresponds to the stage of full expansion and rapid growth of fruit cells, and on 30 dpa, which is the stage close to the beginning of the ripening process. At these stages, we assessed both the stage effects (comparisons between 15 and 30 dpa), and treatment effects (comparison between PRD and FI plants), on proteome variations.
In general, the results show that proteins linked to Carbon metabolism (C) were mostly downregulated during fruit cell expansion in PRD compared to FI fruits at the same growth stage. More protein spots were downregulated at the older growth stage. On the other hand, when the stage effect was compared between FI and PRD plants, a similar pattern of upregulation of protein spots for both PRD and FI fruits was seen. The stage effect results are in accordance with the results of Faurobert and associates (2007), who showed that many of the spots related to C compounds and carbohydrate metabolism were upregulated during tomato fruit development, indicating maximal values in the mature fruits.
Most of the proteins involved in the first category (C metabolism) included those related to primary carbohydrate metabolism (glycolysis and the tricarboxylic acid cycle). Upregulation of several enzymes was seen at 30 dpa compared to 15 dpa in both treatment groups (PRD and FI). Dihydrolipoamide dehydrogenase and pyruvate dehydrogenase are partners in the pyruvate dehydrogenase complex, which transforms pyruvate into acetyl-CoA, and links cytosolic glycolytic metabolism with the tricarboxylic acid cycle. Both enzymes were downregulated in PRD compared to FI treatment. However, pyruvate dehydrogenase patterns of variation in PRD and FI fruits were similar the overall upregulation seen during the development of salt stressed tomato roots (Manaa et al., 2011). Therefore no effect of treatment could be seen, and this was confirmed by the highly significant interaction between stage and treatment effects.
Our results highlight the depressing effect of PRD on glycolysis-related proteins. Two spots, one of them showing upregulation and the other showing significant downregulation in PRD plants, were identified as enolase. This enzyme is also a signal for many environmental stressors, including salt stress, drought, cold, and anaerobic stress, in different plant species (Riccardi et al., 1998; Umeda et al., 1994; Yan et al., 2005), and may act as a positive regulator of some stress-responsive genes (Lee et al., 2002).
The detailed analyses of metabolic regulation underlying tomato fruit development done by Carrari and Fernie (2006) revealed an upregulation of glycolysis prior to the onset of ripening, together with an increase in ethylene biosynthesis rates. The significant expression of triosephosphate isomerase and glyceraldehyde 3-phosphate dehydrogenase, both carbon metabolism proteins, seen at 15 dpa, and the increase seen at 30 dpa in both PRD and FI fruits, supports this metabolic trend.
The other protein in the carbon metabolism category was acid beta-fructofuranosidase. It cleaves sucrose into glucose and fructose, which are sources of energy and carbon for growing tissues, and they increase osmotic pressure and aid in cell elongation and plant growth (Sturm, 1999). In our experiments, acid beta-fructofuranosidase had decreased expression at 30 dpa in PRD fruits compared to FI fruits. This might explain the slower increase in PRD fruit size. Four more proteins that play key roles in the process of glycolysis showed decreased expression in PRD plants at 30 dpa compared to FI plants: pyruvate kinase isozyme G, two isoforms of glyceraldehyde 3-phosphate dehydrogenase, and triosephosphate isomerase.
Similarly, lower levels of two enzymes catalyzing fatty acid synthesis, acetoacetyl-CoA thiolase and 50-kDa ketoacyl-ACP synthase, were found in PRD compared to FI fruits at 15 dpa and 30 dpa. These results indicate lower metabolic flux in PRD fruits compared to FI fruits, and may explain why PRD fruits grew more slowly than FI fruits.
It is also difficult to compare our long-term PRD results with others in the literature because they mostly examined classic drought or salt stress. Central carbohydrate metabolism is known to rapidly readjust after salt stress, and most of the studies dealing with short-term salt or osmotic stress reported a downregulation of carbohydrate metabolism genes (Jiang and Deyholos, 2006; Kerepesi and Galiba, 2000; Ndimba et al., 2005; Wang et al., 1999). The effect of PRD on proteins of carbon metabolism in our experiment shows a similar pattern as the results of studies dealing with short-term osmotic or salt stress.
In the category of amino acid metabolism, significant treatment effects were noted for several enzymes known to be involved in amino acid biosynthesis. Glutamine synthetase, which catalyzes incorporation of inorganic ammonium into glutamic amino acid to form glutamine (Perez-Garcia et al., 1998), showed increased expression in the earlier stage in PRD plants. S-adenosylmethionine synthetase catalyzes the biosynthesis of S-adenosyl-L-methionine. It was downregulated, suggesting decreased synthesis of lignin and other secondary metabolites in PRD fruits. Cytosolic cysteine synthase catalyzes the last step of L-cysteine biosynthesis, which can be used for synthesis of protein or glutathione, and functions as an S-donor for methionine and secondary metabolite biosynthesis (Barroso et al., 1999). Downregulation of this and other amino acid metabolism-related enzymes at 30 dpa under PRD followed the pattern of decreasing primary carbohydrate metabolism.
In the protein translation, processing, and degradation category, several proteinases were identified. Among them, cysteine proteinase 3 showed a more than twofold positive stage effect in both treatment groups. This enzyme is known to participate in programmed cell death and stress responses. Faurobert and associates (2007) observed its regular increase during development until the red ripe fruit phase. Lower expression of cysteine proteinase in PRD fruits is consistent with the slower development seen of these fruits compared to FI fruits. Downregulation of the subunit of the proteasome that is part of the ATP-dependent multicatalytic proteinase complex, both during the cell expansion phase and in PRD plants, might be the result of directing catabolism to save energy.
Energy and cell-wall-related proteins
In the category of energy metabolism, two enzymes that lead to ATP synthesis showed different expression levels only in PRD plants at 15 dpa. ATP synthase delta chain from chloroplast (spot 24) showed increased expression during treatment, while ATP synthase subunit alpha (spot 21), a secreted protein, showed decreased expression. On the basis of the available data it is difficult to explain these opposite changes. We might speculate that there was enhanced demand for energy by chloroplasts, which may have been compensated for by different mechanisms in other cell compartments.
Lower expression of magnesium-dependent soluble inorganic pyrophosphatase, an energy-providing enzyme (Jardin et al., 1995), was found in PRD compared with FI plants at the earlier growth stage. This reduction might be consistent with this enzyme's role in anabolism, as it followed the general trend seen for other carbon metabolism proteins, which were downregulated due to PRD treatment.
In the period between 15 and 30 dpa fruit growth is the result of cell expansion, which aside from energy also requires the biosynthesis of cell wall metabolites. Our proteomic analyses categorized 5 proteins as cell wall-related. Cinnamyl alcohol dehydrogenase (CAD) catalyzes the reduction of various phenylpropenyl aldehyde derivatives and components of the cell wall (Molina et al., 2008). Prolyl 4-hydroxylase catalyzes the formation of 4-hydroxyproline. UDP-glucose:protein transglucosylase-like protein is involved in glucan synthesis, which plays a role in the synthesis of cell-wall polysaccharides (Li et al., 2008). SGRP-1 protein, a glycine-rich protein, belongs to a class of structural cell-wall proteins, and UDP-L-rhamnose synthase plays a role in the synthesis of cell-wall pectic polysaccharides. The higher number of upregulated protein spots at 30 dpa compared to 15 dpa in PRD fruits suggests that metabolic activity in PRD fruits during cell enlargement is oriented toward cell-wall metabolism. When we compared the treatment effect at 15 dpa, almost all cell-wall-related proteins of FI fruits were overexpressed compared to PRD, but at 30 dpa there were no significant differences except for CAD. Our proteomic analysis did not include ripe fruit, because we noted that cell-wall and energy-related proteins generally increase with ripening. We could speculate that higher biosynthetic flux orientation to these pathways, which are both necessary for cell enlargement, allowed PRD fruits to increase the rate of fruit growth, and led to larger diameters of PRD fruits in the final stages of development.
Antioxidative and defense proteins
Among antioxidative enzymes the superoxide dismutases (SODs) act as the first line of defense against oxidative stress by converting superoxide to less-toxic hydrogen peroxide molecules. Ascorbate peroxidases (APX), glutathione peroxidase, and catalase, then detoxify hydrogen peroxide (Foyer and Noctor, 2005). Our previous results demonstrated an increase in cell-wall peroxidase activity during the final stage of PRD tomato fruit ripening (Savic et al., 2008). These results suggest that this enzyme may control fruit maturation by inducing a more rapid process in PRD fruits compared to FI fruits.
There are also results indicating that PRD might cause oxidative stress in the leaves or roots of different plants. In tomato leaves, Tahi and colleagues (2008) demonstrated an increase in SOD, soluble peroxidase (POX), and polyphenol oxidase (PPO) activities in the early phase of water deficit, and then the activity decreased in the remaining phase of the drying cycle. In the same experimental system, the activities of SOD, POX, and PPO showed an alternating increase/decrease, paralleling the alternating irrigation pattern in PRD-treated roots. Upregulation of the activity of antioxidative enzymes was also confirmed by Lei and associates (2009) and Hu and co-workers (2010). However, their results were obtained in a PRD system in which plants were watered with much less water (50% of FI) than we used in our experiment (70% of FI). Furthermore, in our previous experiments we assessed the non-enzymatic antioxidant activity in tomato fruits and potato tubers grown under PRD in polytunnel and field conditions. These results demonstrated that by increasing antioxidant activity, PRD irrigation had a beneficial effect on the nutrient characteristics of these crops (Jensen et al., 2010; Jovanović et al., 2004).
During growth of both PRD and FI fruits our proteomic results demonstrated significant changes in several antioxidant enzymes. Significant upregulation under PRD was found for cTP superoxide dismutase, while cytosolic Cu/Zn-superoxide dismutase was unchanged. Our results also showed a significant increased stage effect for cyt ascorbate peroxidase in PRD fruits, which together with increased SOD protein expression, could significantly improve resistance to PRD-induced drought stress. This would allow the PRD fruits to continue to grow, and after 30 dpa to produce larger FI fruits.
The increase in the expression of antioxidant enzymes could also be related to fruit developmental stage. The results of Faurobert and associates (2007) confirmed an overexpression of H2O2-scavenging enzymes from the breaker to the ripening phases of tomato fruits. Although our analyses did not include the ripening phase, increased expression of 3 APX enzymes located in different compartments, and of cyt SOD, could be related to developmental changes in both PRD and FI fruits. However, in PRD fruits APX enzymes were following similar temporal expression patterns as cell-wall-related proteins. APXs were downregulated at 15 dpa, but their biosynthesis was then intensified, and at 30 dpa they reached the expression levels of the APXs in FI fruits. This suggests that PRD treatment induced mild drought stress, which induced a cascade of resistance reactions in fruits, starting with the increased biosynthesis of SODs seen at the beginning of the cell expansion phase.
In our proteomic analysis 4 proteins were classified as stress defense proteins, and they included 1 heat-shock protein. Both oxygen-evolving enhancer proteins are important for maintaining water and high rates of oxygen evolution during photosynthesis (Hoganson and Babcock, 1997). There are findings that confirm a significant role for these proteins in increasing stress resistance (Ali and Komatsu, 2006). We did not find any significant stage effect on the expression of these proteins in either treatment group. However, comparisons between treatments showed an overexpression of both proteins in PRD compared to FI fruits.
Among this category of proteins a different expression pattern was also found for heat-shock protein 70 (HSP 70). This protein has a role in a variety of cellular processes by keeping proteins in a competent state (Sung et al., 2001), and has been found to be upregulated in tomato fruit after cold stress (Page et al., 2010). The results of Manaa and colleagues (2011) showed that among the 4 HSPs identified in their salt study, HSP 70 was the only one that was more abundant in sensitive varieties, and was upregulated in all tomato genotypes by salt stress. Our results showed that HSP 70 was upregulated during the phase of rapid growth of PRD fruits. Comparisons between treatments demonstrated downregulation in PRD plants at 15 dpa, but in the next phase of fruit growth the differences between treatment groups were small. The expression patterns of HSP 70 followed the expression patterns of ascorbate peroxidases and cell-wall-related proteins in PRD fruits. The higher expression of stress defense proteins at 30 dpa in PRD fruits, and the similar pattern of HSP 70 compared to FI plants, also indicated a possible time-course activation of stress defense mechanisms that was also seen for antioxidative enzymes during the growth process of PRD fruits.
Conclusion
This study showed that tomato plants under 70% PRD could produce fruits with a larger diameter compared to FI fruits, whereas the total fruit fresh and dry weights were similar in both treatment groups. Although the growth rate was higher in FI than in PRD fruits, the longer period of cell expansion resulted in larger PRD fruits.
At the proteome level, the expression of proteins related to carbon and amino acid metabolism mirrored fruit growth rates, and indicated that the slower metabolic flux seen in PRD fruits may be the cause of their slower growth rate compared to FI fruits. On the basis of the increase in expression seen of the cell-wall proteins, energy-related proteins, and stress defense proteins, one might also speculate that metabolic flux orientation toward the cell-wall proteins, and the production of energy necessary for cell enlargement, allowed PRD fruits to increase their rate of growth, which at the end of the developmental phase led to a larger diameter compared to FI fruits. The upregulation of some antioxidative enzymes during the cell expansion phase of PRD fruits appeared to be related to their role in protecting fruits against the mild stress induced by this treatment.
Overall, our data provide novel insights into the understanding of fruit growth and protein function in the pericarp cell expansion process. Future proteome investigations are needed to further analyze later phases of fruit development, especially during ripening, which could be of great interest, and help us understand the effects of PRD not only on fruit growth, but also on fruit quality.
Footnotes
Acknowledgements
This study was supported by The “Pavle Savić” Program of Bilateral Cooperation between Serbia and France 2010–2011, COST Action (COST-STSM-FA0603-03178), and the Serbian Ministry of Education and Science project TR 31005. Many thanks are due to Esther Pelpoir, Caroline Callot, Karine Leyre, Arafet Manaa, and Jean-Paul Bouchet Benoît Valot, for their help in proteome analysis, and to our other colleagues at INRA in Avignon for their hospitality.
Author Disclosure Statement
No competing financial interests exist.