
Abstract
Abstract
Different abiotic stresses inhibit or delay the development and growth of plants. The most crucial step of plant life cycle, which ensures the survival of the next generation, is seed germination. Plants are sessile organisms that need to integrate internal and external signals in order to produce the correct response. Plants have evolved mechanisms that enable seed germination to be arrested under stress conditions and then resumed when conditions are favorable. The complexity of this mechanism was explored in Arabidopsis thaliana using mutants that had defects in their phytohormone metabolism and signaling pathways. These analyses led to the identification of many important components that are involved in these pathways and shed light on the complex crosstalk between phytohormones under abiotic stress. Combined “omics” techniques such as functional genomics, transcriptomics, and proteomics with the support of bioinformatics, physiology, and molecular genetics have greatly expanded the present understanding of the seed germination process. This minireview focuses on the current status of knowledge about seed germination under abiotic stress with a particular emphasis on genetic interactions, hormonal balance, and epigenetic regulation that occur in Arabidopsis thaliana during this process.
The Three Phases of Water Uptake During Seed Germination Process
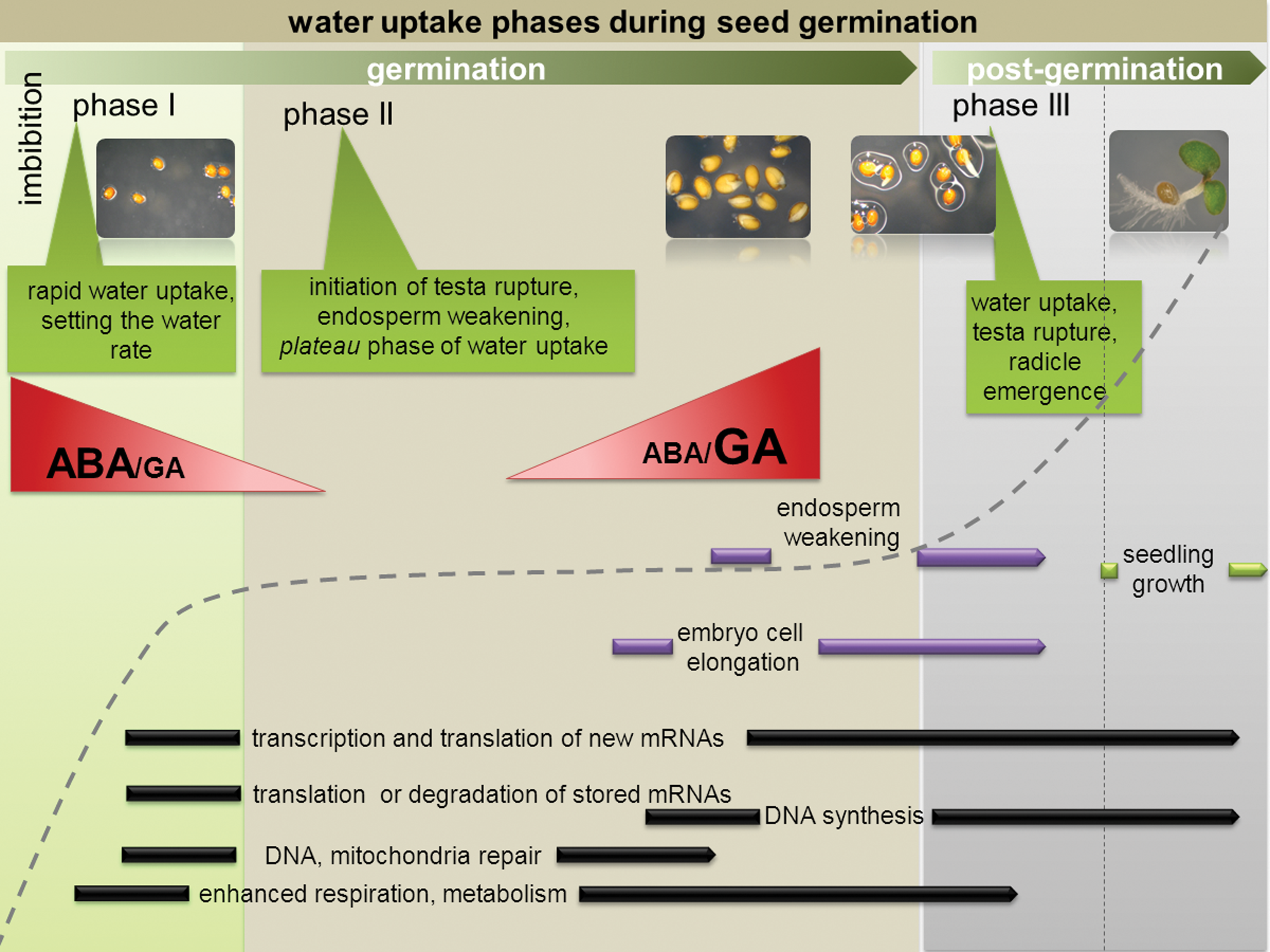
Although a precise description of genes and hormones involved in the seed coat development is available for Arabidopsis, yet little is known about the changes in the seed coat biochemical and physical properties that lead to testa rupture (Leubner-Metzger, 2003). Endosperm rupture by the expanding radicle is closely associated with endosperm weakening. The process of endosperm weakening is linked with cell wall remodeling enzymes (CWRE), which include endo-β-mannanase (Bewley, 1997), β-1,3-glucanases (Leubner-Metzger, 2003), xyloglucan endotransglycosylase (Chen et al., 2002), and others. Prior to the completion of seed germination, the embryo cells elongate. It is not clear which cells are responsible for the initial emergence of the radicle. The emergence could be facilitated by a reduction in the resistance of the tissue surrounding the radicle through specific modifications of the cell walls by CWRE (Schopfer, 2006). This implies that embryo growth during germination depends on changes in the extensibility of the cell wall and is accompanied by progressive vacuolation during late water uptake in Phase II (Bethke et al., 2007). Sliwinska and coworkers (2009) recently reported that embryo elongation occurs in the basal part of the hypocotyl and the hypocotyls-radicle transition zone.
Crosstalk Between Phytohormones During Seed Germination
Numerous genes that regulate the transition from Phase II to Phase III have been identified in Arabidopsis. The transition is closely associated with hormonal and environmental changes and includes the actions of genes involved in the signal transduction of ABA: ABI3 (ABA INSENSITIVE 3), ABI5 (ABA INSENSITIVE 5), gibberellins: RGL2 (REPRESSOR OF GA1-3 LIKE 2), and ethylene: EIN2 (ETHYLENE INSENSITIVE 2). Cooperation between different hormones plays a significant role in seed germination.
It is well known that gibberellins (GAs) and abscisic acid (ABA) act as antagonists during seed germination. The hormonal balance is under the control of environmental factors such as light and cold. Before imbibition, seeds are in a dormant state or are quiescent when nondormant seeds are considered. Seed dormancy is an inability of viable seeds to undergo germination under optimal conditions (North et al., 2010). In Arabidopsis, a large number of mutants that enable dormancy and germination processes to be better understood were obtained after mutagenesis, for example: abi3 (Giraudat et al., 1992; Parcy et al., 1994). In addition, studies that engaged natural variation have been carried out in order to understand the mechanisms of dormancy and seed germination that have evolved naturally. Differences in seed dormancy between Landsberg erecta (Ler) and Cape verdi (Cvi) led to the generation of a recombinant inbred line population and the identification of DOG1 (DELAY OF GERMINATION1), which controls seed dormancy. The transcription of DOG1, which belongs to a small gene family, is seed specific and drops during seed imbibition (Bentsink et al., 2006). The addition of glucose induced the expression of the DOG1 Cvi allele, but not the Ler or Col-0 alleles. It was found that sugar induction of ABI4 was promoted by the DOG1 Cvi allele (Teng et al., 2008).
Dormancy is induced and maintained by ABA and released by GAs. During imbibition, light and cold act together to break dormancy and to promote seed germination by increasing GAs levels. A rapid decrease of ABA endogenous content during Phase II is one of many factors that influence the successful completion of germination (Fig. 1). The endogenous content of ABA is mainly regulated by 9-cis-epoxycarotenoid dioxygenases (NCEDs) and ABA 8′-hydroxylases (CYP707As), enzymes of ABA biosynthesis and catabolism, respectively. The spatial pattern of CYP707A expression was identified in the radicle upon seed imbibition (Okamoto et al., 2006). The inhibitory action of ABA during seed germination is counteracted by GAs and by ethylene (ET). GAs play an important role during the early and the late phases of germination and act against the inhibitory effect of ABA. During the early phase of germination, GAs mainly induce the expression of genes encoding cell walls remodeling enzymes (CWRE), such as α-expansins (Debeaujon and Koorneef, 2000; Weitbrecht et al., 2011), whereas during the late phase of germination gibberellins are involved in the direct stimulation of the growth of the embryo (Debeaujon and Koorneef, 2000). Bioactive GAs were already present in dry, afterripened seeds. Transcripts of GA20ox and GA3ox, key regulatory genes involved in GAs biosynthesis, accumulate during the early phase of germination (Ogawa et al., 2003). In terms of rapid ABA degradation, the GA/ABA ratio increases threefold during the early germination phase and 10-fold during the late germination phase (Weitbrecht et al., 2011). Arabidopsis CHOTTO1 (CHO1), which encodes a double APETALA2 transcription factor, is thought to be involved in the regulation of GA3ox1 and GA3ox2 expression during seed imbibition through a collaboration with other seed-specific factors. cho1 mutation is able to restore the delayed seed germination of the cyp707a2-1 mutant but did not restore the seed germination of ga1-3 (GA-deficient1–3). Together these data show that CHO1 acts downstream of ABA to repress GA biosynthesis during seed imbibition. Changes in the expression pattern of the gene involved in ABA and GAs metabolism observed in imbibed seeds of cho1 were correlated with altered levels of endogenous ABA and GAs when compared to the wild type. These results clearly show that CHO1 is involved in the regulation of the endogenous ABA and GAs balance in imbibed seeds (Yano et al., 2009). In addition, CHO1 is involved in the seed germination process and phytohormone metabolism through interactions with other factors described below. The cho1 mutant, which shows reduced responsiveness to ABA, is able to germinate in the presence of a high concentration of glucose and potassium nitrate. Analysis show double mutants cho1 abi5, and cho1 abi4 showed that mutation in CHO1 enhanced the ABA response of the abi5 but not the abi4 mutant. Based on these data, CHO1 is thought to be involved in the same pathway as ABI4 but not ABI5. Induction of the expression of CHO1 did not occur in the abi4 mutant, which indicates that this process is ABI4-dependent. The response to nitrate was unique to the cho1 mutant when compared to abi4. CHO1 regulates nutritional responses downstream of ABI4 during seed germination (Yamagishi et al., 2009).
Ethylene plays an important role only during the late phase of germination. It is able to counteract ABA negative regulation of seed germination by interacting with ABA. The ethylene-responsive mutant ein2 (ethylene insensitive 2) is hypersensitive to ABA, overaccumulates ABA in dry seeds (probably due to an effect on ABA catabolism), and its seed hyperdormancy can be canceled by stratification. The effect of increased dormancy can be completely counteracted by mutations in ABI genes, such as abi3–4, which display a strong ABA-insensitive phenotype. As a consequence, the ein2 abi3–4 double mutant is as nondormant as the abi3–4 single mutant. The nondormant phenotype of the ein2 abi3–4 double mutant suggests that ethylene suppresses seed dormancy by inhibiting the action of ABA. Thus, EIN2 is a possible negative regulator of ABA response. ET, together with GAs, antagonizes ABA actions during the induction of seed dormancy (Beaudoin et al., 2000). During the early phase of seed germination, a decreased content of jasmonic acid (JA), salicylic acid (SA) and an increased level of auxins was observed in Arabidopsis seeds (Preston et al., 2009). Both JA and SA were shown to act as negative regulators of seed germination (Dave et al., 2011; Rajjou et al., 2006). Auxins are considered to be regulators of the seed germination process in a crosstalk with GAs, ABA, and ET (Carrera et al., 2008). The brassinosteroids signal leads to a reduced sensitivity to ABA and stimulates germination (Steber and McCourt, 2001).
To summarize, germination commences with water uptake, which is triphasic and lasts until the emergence of the radicle. A complicated network of changes in transcription and protein metabolism together with correlated changes in endogenous phytohormones such as ABA, GAs, BRs, ET, SA, and auxins regulate the germination process in order to ensure survival of next generations.
Seed Germination Under Abiotic Stress
In natural environmental conditions, seeds have to overcome abiotic stresses in order to ensure the survival of the next generation. The ability to tolerate and resist salt, osmotic stress, dehydration, and cold stress during germination is essential for plant survival. When taking into account changes in water content in a seed and triphasic water uptake during imbibition prior to radicle emergence, it is obvious that seed germination cannot take place in the absence of water. When sudden abiotic stress, osmotic stress, salt, or dehydration occurs between a seed's first exposure to water and cotyledon greening, the germination process can be arrested. This process is regulated by a network of transcription factors that have both overlapping and discrete functions (Finkelstein, 2004; Finch-Savage and Leubner-Metzger, 2006; Hilhorst, 2007). In order to maintain or break the period of arrested germination and to complete germination, different metabolic pathways including phytohormones biosynthesis and signal transduction pathways, chromatin modifications, and microRNA posttranscriptional regulation, are involved.
Salt stress can influence many physiological processes from seed germination to plant development. The complexity and severity of a plant's response to salt stress can be explained by the fact that salinity works on two levels: ionic toxicity and osmotic stress. Different sets of genes regulate response to salt during different stages of development (Quesada et al., 2002). At the cellular level, salt tolerance mechanisms function to reduce sodium accumulation in the cytoplasm. This is achieved by the active transport of sodium out of the cell through plasma membrane Na+/H+ antiporters, that is, SOS1 (Salt Oversensitive 1) (Shi et al., 2005) or addressing it into the vacuole by tonoplast antiporter NHX1 (Na+/H+ Exchanger) (Blumwald et al., 2000; Chung et al., 2008).
In order to cope with dehydration and osmotic stresses, plants have evolved complicated machinery to synthetize and accumulate metabolites, “osmoprotectants,” which help to withstand osmotic pressure and maintain turgor and the driving gradient for water uptake. Osmoprotectants include low molecular weight compounds such as amino acids, polyols, sugars, and methylamines (Hasegawa et al., 2000). These compatible metabolites stabilize the enzyme structure, cellular membranes, and other cellular components during any exposure to stress (Chaves et al., 2009; Zhu, 2001, 2002, 2003). In response to stress, de novo synthesis of dehydrins, osmotins, and LEA (Late Embryogenesis Abundant) proteins occurs. Osmotins and dehydrins stabilize the integrity of the cellular membranes and protein structure, whereas LEA are able to sequestrate ions and water in order to protect cellular components against stress (Hundertmark et al., 2011; Kosova et al., 2011).
The Regulation of Transcription by Phytohormones in Response to Abiotic Stresses During Seed Germination
ABA and its role in response to abiotic stress during seed germination
Before imbibition over 10,000 different mRNAs are stored in the dry seeds of Arabidopsis (Okamoto et al, 2010). Among them, transcripts of genes that have ABA responsive elements within their promoter sequences are overrepresented. This implies that ABA has a significant role during seed development (Nakabayashi et al., 2005). Many components of ABA signaling take part in the regulation of seed germination under abiotic stress. Kinases are important for stress response; some like SnRK2 (SUCROSE NONFERMENTING RELATED KINASES 2) are known to be signal mediators. The identification of ABA receptors—PYR/PYL/RCAR (PYRABACTIN RESISTANCE1/PYR1-LIKE/REGULATORY COMPONENT OF ABA RESPONSE 1)—has provided a breakthrough in understanding the relations between some ABA signaling components (Ma et al., 2009; Nishimura et al., 2010; Park et al., 2009; Santiago et al., 2009). These ABA receptors regulate the phosphatases PP2C (PROTEIN PHOSPHATASES 2 C) interacting with SnRK2 (Fujii et al., 2009; Melcher et al., 2009; Nishimura et al., 2010; Santiago et al., 2009; Yin et al., 2009; reviewed in Umezawa et al., 2010). SnRK2 are important for activating transcription factors that are crucial for seed germination, such as ABI5 (ABA INSENSITIVE 5) (Fujii et al., 2007; Nakashima et al., 2009; reviewed in Wasilewska et al., 2008).
Three well-characterized transcription factors ABI3 (ABA INSENSITIVE 3), ABI4 (ABA INSENSITIVE 4) and ABI5 (ABA INSENSITIVE 5) were isolated in screens for ABA insensitivity during germination. They belong to different transcription factor families: B3-, APETALA2-, and bZIP-domain and regulate overlapping subsets of seed-specific and/or ABA-related genes (Finkelstein, 1994; Finkelstein and Lynch 2000; Finkelstein et al., 1998; Giraudat et al., 1992; Lopez-Molina and Chua, 2000). ABI3 and ABI4 are expressed from globular stage embryogenesis, and their products can regulate ABI5 expression, which is activated at the heart stage. ABI5 also activates its own expression, similarly to ABI4 (Bossi et al., 2009; Garcia et al., 2008). All three transcription factors are most highly expressed in mature seeds, but differ slightly in localization within the seeds (Brocard et al., 2002; Finkelstein and Lynch, 2000; Lopez-Molina et al., 2002; Parcy et al., 1994; Penfield et al., 2006; Soderman et al., 2000). Although abi3, abi4, and abi5 mutants were initially isolated as ABA-insensitive (Finkelstein, 1994), they have also been shown to have defects in response to glucose, NaCl, and osmotic stress during germination and seedling growth (Arenas-Huertero et al., 2000; Brocard et al., 2002; Laby et al., 2000; Quesada et al., 2002).
During seed imbibition under nonstress conditions, the level of ABI3 and ABI5 mRNAs and proteins rapidly decrease in order to enable the completion of the germination process. A decrease in the level of proteins occurs through the interaction of RING E3 ligases: AIP2 (ABI3-INTERACTING PROTEIN 2) with ABI3 and KEG (KEEP ON GOING) with ABI5 (Stone et al., 2006; Zhang et al., 2005). This action of the ubiquitin/26S proteasome system can be arrested or delayed in response to osmotic stress or a high exogenous ABA signal. In the presence of abiotic stresses and/or exogenous ABA, the degradation process of ABI3 and ABI5 is slowed. It has been shown that ABA promotes KEG self-degradation via the ubiquitin dependent 26S proteasome pathway (Liu and Stone, 2010). ABA and/or abiotic stresses induce the expression of ABI3 and ABI5 in order to activate their respective signal cascade transduction (Stone et al., 2006; Zhang et al., 2005). ABI5 action is also regulated by sumoylation, which both represses its activity and increases its stability (Miura et al., 2009). Posttranslational modification of the ABI3, farnesylation, occurs through its interaction with ERA1 (ENHANCED RESPONSE TO ABA 1) (Brady et al., 2003). Recent findings of Reeves and coworkers (2011) have shown that ABI4 is also regulated in the proteasome-mediated degradation pathway in order to prevent proteins from accumulating to high levels. The mechanisms and their components remain unknown.
Another ABA-related gene that regulates the response to salt, osmotic, and cold stress during seed germination is RAP2.6 (Related to APETALA 2), initially isolated as activated by JA, ethylene, SA, and two virulent strains (He et al, 2004; Wang et al, 2008a). The expression of RAP2.6 is also upregulated by ABA, salt, osmotic, and cold stresses (Zhu et al., 2010). Overexpression lines of RAP2.6 displayed a hypersensitivity to ABA and abiotic stresses during germination and early seedling development. Stomatal movements, drought, and salt tolerance were studied in adult wild-type and transgenic plants. No significant differences were observed, which suggests that RAP2.6 plays a role in the ABA response to abiotic stress during seed germination and early seedling growth. Further studies showed the presence of abiotic- (ABRE, DRE, MYBR) and biotic- (W-box, RAV-box) related cis-regulatory motifs within the promoter region of RAP2.6. These data suggested that RAP2.6 may serve as a common point in crosstalk, not only for hormones, but also for biotic and abiotic stress responses (Zhu et al., 2010).
RACK1 (Receptor for Activated C Kinase 1) is another ABA response gene associated with salt stress response during germination. Originally, RACK1 was identified as a receptor of activated protein kinase C (PKC) in mammalian cells (Mochly-Rosen et al., 1991; Ron et al., 1994). It is now known that this multifunctional protein plays a role in the regulation of many of metabolic pathways. The sequence and structure of RACK1 is highly conserved in plants (Chen et al., 2006; Guo et al., 2007; Ullah et al., 2002). The response of Arabidopsis rack1a mutants to salt stress during germination revealed its hypersensitivity, which is consistent with its hypersensitive response to ABA. The ABA-responsive marker genes, RAB18 (Responsive to ABA 18) and RD29B (Responsive to Desiccation 29B), showed upregulation in rack1a mutant. These data suggest that RACK1 encodes a critical regulator of ABA signaling in terms of salt stress response during seed germination (Guo et al., 2009).
Gibberellins—another group of phytohormones involved in stress response
Seed germination is antagonistically controlled by ABA and GAs. GAs promote seed germination by enhancing the proteasome-mediated degradation of RGL2 (RGA-LIKE2), a negative regulator of GAs signaling, primarily controlling seed germination. RGL2 has a conserved DELLA motif, which is essential for its proteasome-mediated degradation (Lee et al., 2002). After binding GAs to the receptor GID1 (GIBBERELLIN INSENSITIVE DWARF1), SLY1 (SLEEPY 1)—an F-box protein interacts with RGL2 facilitating its ubiquitination and subsequent destruction (Feng et al., 2008). There are four other DELLA factors: RGA (REPRESSOR OF GA1–3), GAI (GIBBERELLIN INSENSITIVE), RGL1, RGL3. All of these are expressed during seed germination (Tyler et al., 2004). The function of RGL2 in salt stress response was highlighted by Kim and coworkers (2008), who observed an elevated RGL2 transcript level in seeds germinated under salt stress and Achard et al. (2006), who showed that a quadruple DELLA mutant (gai, rga, rgl1, rgl2) is more sensitive to salt stress than the wild-type and DELLA single mutants.
Crosstalk between ABA and GAs under stress during seed germination
Ko and coworkers (2006) reported that RGA overaccumulation upregulates XERICO and subsequently stimulates ABA synthesis. XERICO encodes a RING-H2 zinc-finger factor, which promotes ABA accumulation in an unknown mechanism (Zentella et al., 2007). An increased level of endogenous ABA activates ABI5 expression, which consequently arrests seed germination.
Yuan and coworkers (2011) showed an interaction between components of ABA (ABI3 and ABI5) and GAs (RGL2) signaling pathways in response to salt during germination and early seedling development. This strongly supported the cooperation between these two phytohormones that was already known. The findings of Yuan and others (2011) have enriched knowledge about the involvement of GAs in abiotic stress response. They studied two ABA signaling mutants, well known for their salt insensitive phenotype, abi3 and abi5, and the GAs signaling mutant rgl2. The rgl2, abi3 and abi5 are resistant to high sodium chloride (salt) and mannitol (osmotic) concentrations during germination. These data suggest that both ABA and GAs are involved in salt and osmotic stress inhibition and the delay of seed germination. The level of expression of RGL2, ABI3, and ABI5 was investigated in each mutant background with similar results. After treatment of seeds with NaCl, the expression of RGL2, ABI3, and ABI5 was significantly lower in any mutant background than in the wild-type. In the wild-type genotype expression of all the genes being studied was highly induced by salt stress. These data clearly show that RGL2, ABI3, and ABI5 are required for induction of each other during salt stress and strongly also demonstrate crosstalk between ABA and GAs when seeds are exposed to abiotic stress (Yuan et al., 2011) (Fig. 2).
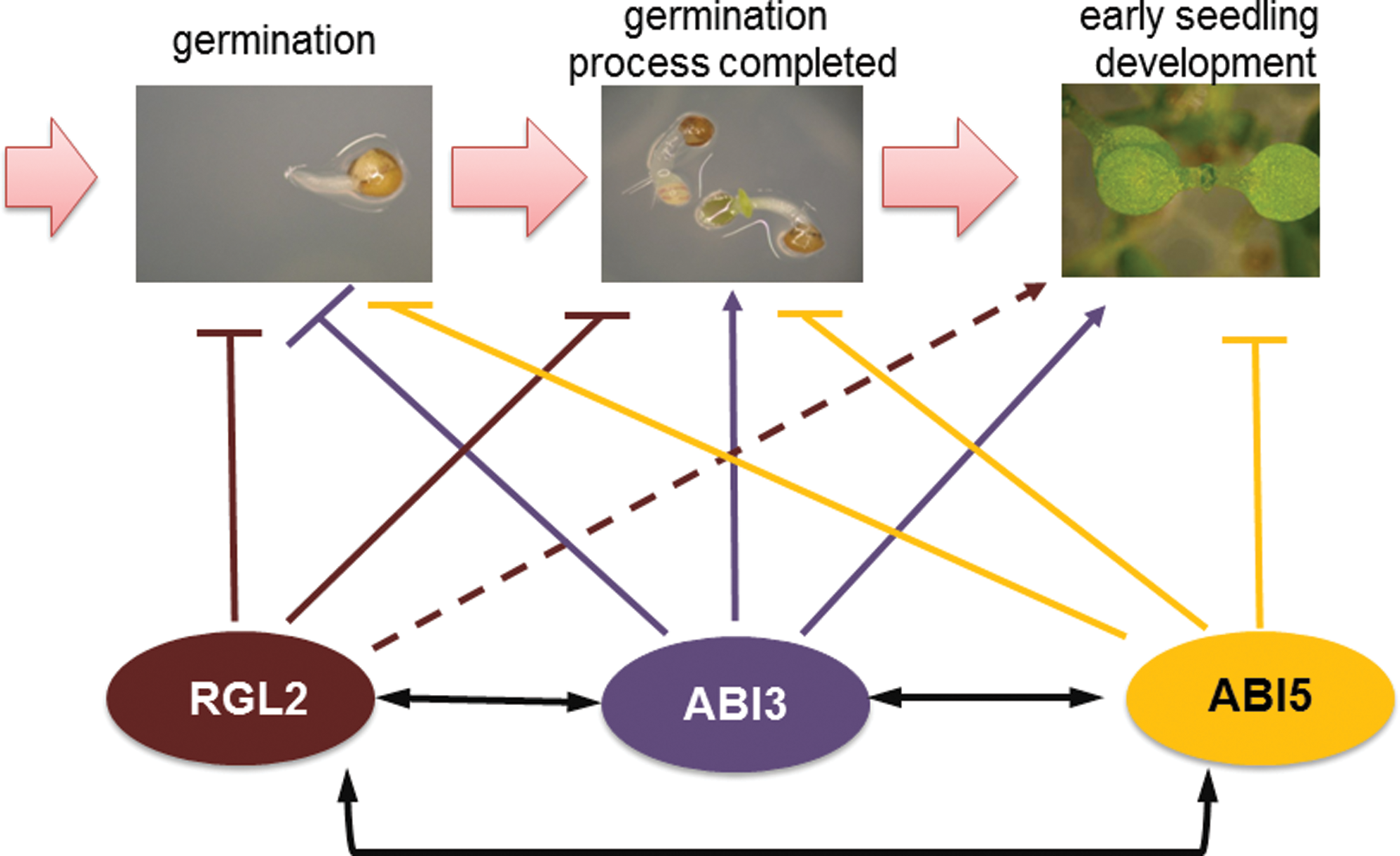
The possible interaction between ABA signaling components: ABI3, ABI5, and GA signaling component: RGL2 during seed germination under salt stress (Yuan et al., 2011). Lines with blunt end indicate negative regulation, arrows indicate positive regulation, dashed lines indicate hypothetical interaction.
Brassinosteroids are engaged in salt stress response in concert with ABA
Brassinosteroids are steroid hormones that play an important role in the growth and development of various organisms (Steber and McCourt, 2001). Exogenous application of brassinosteroids (BRs) was reported to improve salt stress tolerance in rice and Arabidopsis (Anuradha and Rao, 2001, 2003; Kagale et al., 2007), but little is known about the impact of endogenous BRs on stress response. The components of BR signaling have been identified and characterized using BR-deficient or BR-insensitive mutants. The membrane-localized receptors BRI1 and BAK1 bind BR and transfer a signal into the cytoplasm to inhibit BIN2, a glycogen synthase kinase-3/SHAGGY-like kinase. BIN2 negatively regulates the downstream signaling components by phosphorylating two transcription factors, BES1 and BZR1, and inhibiting their binding to BR target promoters (Li and Jin, 2006; Wang and He, 2004). Arabidopsis DET2 encodes an enzyme involved in BR biosynthesis. The defects in root and leaf development of the mutant det2-1 can be rescued by the application of brassinolide (Li et al., 1996).
The involvement of endogenous BR in stress response can be studied using BR-deficient or BR-signaling mutants. Koh and others (2007) demonstrated that the rice knock-out mutant OsGSK1, which is hypersensitive to BRs, is more tolerant to salt stress. Zeng and Hua (2010) investigated the response to salt stress of Arabidopsis BR-deficient mutant det2-1 and BR-signaling mutant bin2-1. The seed germination and seedling growth of both, det2-1 and bin2-1, were hypersensitive to salt stress. The hypersensitivity of det2-1 was associated with an inhibited induction of stress-related genes, such as COR78 (COLD REGULATED 78) when compared to the wild type, whereas RD22 (RESPONSE TO DESICCATION 22) the salt-induced level was maintained in both det2-1 and the wild-type. The proline content was lower in det-2 than in the wild-type. The same results were observed in the case of a bin2-1 mutant. Exogenous epibrassinolid (EBR) could partially rescue the growth of det2-1 under salt stress and improve NaCl-induced proline accumulation (Zeng and Hua, 2010). Recently, the inhibitory role of ABA in the modification of the phosphorylation status of BES1 and its impact on BR-responsive gene expression has been shown (Zhang et al., 2009). These data, together with det2-1 hypersensitivity to ABA (Zeng and Hua, 2010), point to a link between BR and ABA in seed germination under abiotic stress.
SA—alone and in concert with GAs and ABA—is involved in stress response during seed germination
SA is a phenolic compound. It has been demonstrated that SA is a critical signaling molecule involved in the regulation of plant responses to pathogen infections and abiotic stresses through extensive signaling crosstalk with other hormones (Vlot et al., 2009; Wolters and Jurgens, 2009). Recently, it was suggested that SA is engaged in the seed germination process as a key regulator (Alonso-Ramirez et al., 2009). Nishimura and coworkers (2005) reported that SA inhibits seed germination in a dosage-dependent manner. A high SA concentration such as >1 mM may delay or even inhibit germination, possibly by causing a toxic effect by inducing reactive oxygen species (ROS) biosynthesis (Rajjou et al., 2006). Low doses of exogenous SA (1–10 μM), close to physiological concentrations, significantly improve seed germination under stress (Alonzo-Ramirez et al., 2009). Proteomics studies have shown that SA treatment improves seed germination by promoting the synthesis of proteins that are crucial for germination, and by the mobilization or degradation of seed proteins accumulated during seed maturation. Proteomics data have also revealed that SA can induce an accumulation of proteins characteristic for ABA signaling, such as LEA. Taking into account that SA can induce a transient accumulation of ABA in wheat seedlings (Shakirova et al., 2003), it can be assumed that Arabidopsis undergoes the same process during seed germination. This could explain the accumulation of ABA-related proteins and the delay of seed germination by SA in nonstress conditions (Rajjou et al., 2006). It was proven that exogenous application of GA counteracted the inhibitory effects of salt, oxidative or heat stress during germination and seedling development in Arabidopsis (Achard et al., 2006). This is closely related to an increase in SA levels as a result of GAs action (Navarro et al., 2008; Robert-Seilaniantz et al., 2007). Alonzo-Ramirez et al. (2009) reported that exogenously applied SA partially rescued seed germination in ga1–3 (GA-deficient 1–3) mutants in the presence of NaCl. After GA3 was applied, nearly 100% of Col-0 seeds were able to complete germination and start seedling growth compared with no significant differences in the germination rate of a sid2 (SA-induction-deficient 2) mutant that was hypersensitive to salt stress. To summarize, these data together with the SA induction of LEA proteins accumulation showed a link between SA, ABA, and GAs during stress response in germinating seeds.
ABA is thought to be the main stress hormone when seed germination is considered. However, seed germination under stress conditions is regulated by various phytohormones as was revealed through the use of mutants in genes involved in phytohormone metabolism and signaling pathways (Table 1). All of the possible interactions between the phytohormones involved in the seed germination process under stress are shown in Figure 3.
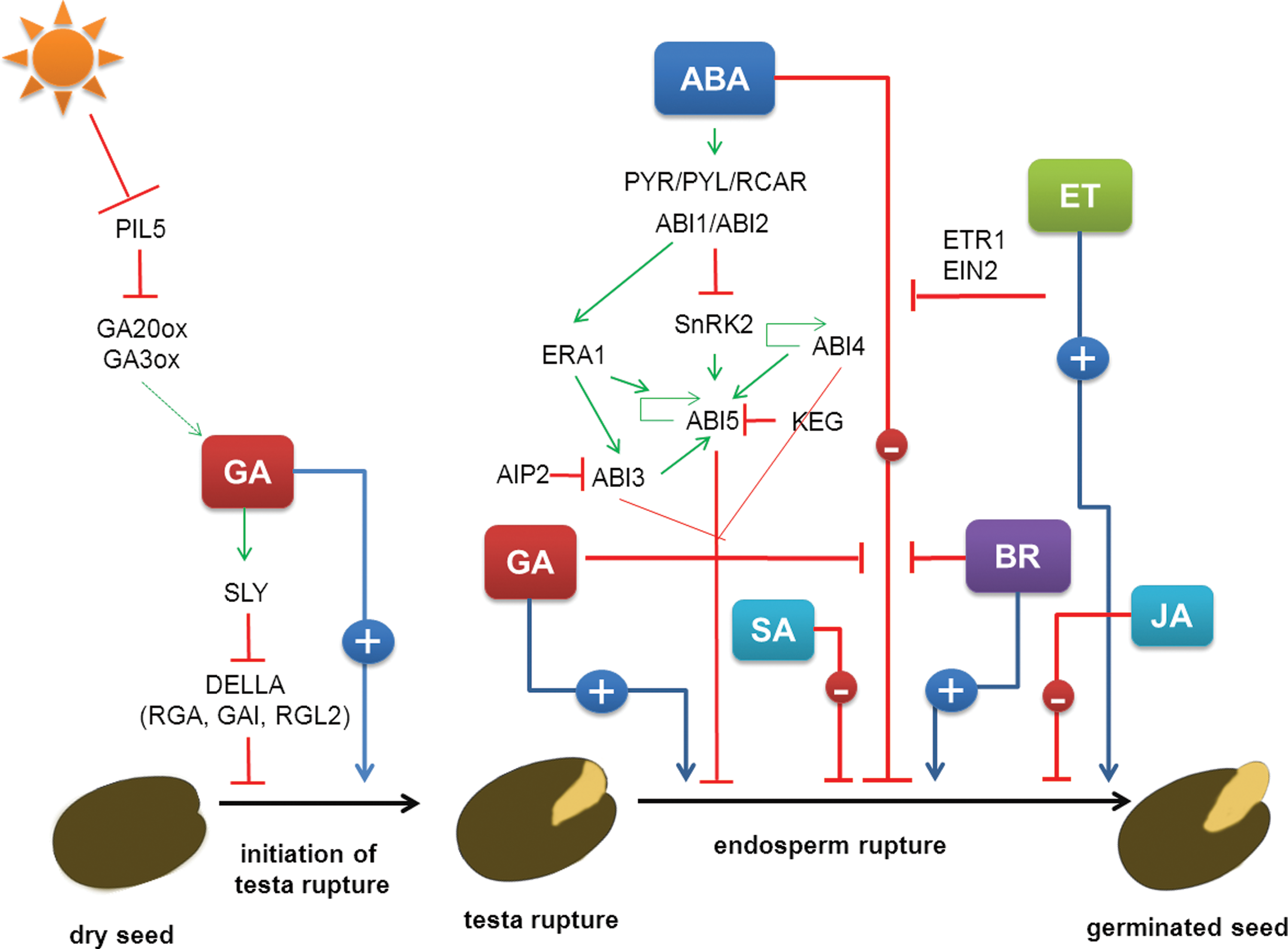
Hormonal regulation of the seed germination process under both nonstress and stress conditions. Visible events during two-step germination: testa and endosperm rupture. GA, gibberellin; ABA, abscisic acid; BR, brassinosteroid; SA, salicylic acid; JA, jasmonic acid; ET, ethylene; PIL5, PHY-INTERACTING FACTOR 2; GA20ox, GIBBERELLIC ACID-20-OXIDASE; GA3ox, GIBBERELLIC ACID-3-OXIDASE, SLY1–SLEEPY1; RGA, REPRESSOR OF GA1–3; GAI. GIBBERELLIN INSENSITIVE; RGL2, RGA-LIKE 2; PYR/PYL/RCAR, PYRABACTIN RESISTANCE1/PYR1-LIKE/REGULATORY COMPONENT OF ABA RESPONSE; ABI1/2/3/4/5, ABA INSENSITIVE 1/2/3/4/5; SnRK2, SUCROSE NONFERMENTING RELATED KINASE 2; KEG, KEEP ON GOING; AIP2, ABA INSENSITIVE 3 INTERACTING PROTEIN 2; ERA1, ENHANCED RESPONSE TO ABA 1; ETR1, ETHYLENE RECEPTOR 1; EIN2, ETHYLENE INSENTIVE 2. A plus or a minus indicates a positive or negative effect on the seed germination process. The interactions that are shown are described in detail in the text.
TF, transcription factor; PP, phosphatase; E3, E3 ligase; RBP, RNA binding protein; RF, RING FINGER protein; FT, farnezyltransferase; **, phosphatidylethanolamine binding; MAP, MAP kinase; K, kinase; BE, biosynthesis enzyme; R, receptor, ?, unknown function.
Epigenetics Is Involved in the Regulation of Seed Germination Under Stress
Abiotic stresses can regulate gene expression through the induction of changes in the DNA methylation status. One example is the action of PICKLE (PKL)-mediated chromatin-remodeling events during seed germination in response to osmotic stress and ABA induction. During seed imbibition under nonstress conditions, the levels of ABI3 and ABI5 mRNAs and proteins rapidly decrease in order to enable the completion of germination process. This process was described in detail above. Using pkl mutants Perruc and others (2007) showed that pkl overexpressed both ABI3 and ABI5 in response to ABA. PICKLE encodes a putative SWI/SNF-class chromatin-remodeling factor of the CHD3 type. Chromatin immunoprecipitation (ChIP) analysis revealed that mutant pkl seeds had two- to 2.5-fold lower H3-K9 and H3-K27 methylation levels at the ABI3 and ABI5 promoter when compared to the wild type after ABA treatment. In contrast, 7-day-old wild-type and pkl seedlings displayed no differences in the methylation of H3-K9 and H3-K27 within the promoters of both ABI genes. It has been suggested that PKL is essential for repressing ABI3 and ABI5 during seed germination in the presence of exogenous ABA and osmotic stress (Perruc et al., 2007).
The Role of microRNA in the Regulation of Seed Germination Under the Stress
microRNAs (miRNAs) are small RNAs that perform posttranscriptional gene silencing through the guided cleavage of complementary target mRNAs (Llave et al., 2002) or translational repression (Brodersen et al., 2008). miRNA regulation is also involved in early plant development events, such as seed germination and young seedling growth. Findings of Reyes and Chua (2007) showed the crucial role of miR159 in the negative regulation of ABA signaling during seed germination. The response to exogenously applied ABA mimics the response to abiotic stress. When ABA is applied, miR159 accumulates in seeds and downregulates MYB33 and MYB101, positive regulators of ABA signaling, which enables the completion of seed germination. Reyes and Chua (2007) also reported the involvement of ABI3 and ABI5 in the regulation of MIR159 transcription.
Another miRNA involved in seed germination is miR160, which regulates ARF10 (Auxin-Responsive Factor 10). miR160 mutants exhibited a changed ABA sensitivity (Liu et al., 2007). ARFs are important factors for auxin signaling; therefore, this result suggests crosstalk between auxins and ABA during seed germination (Guilfoyle and Hagen, 2007). Microarray analyses indicate upregulation of ABA-related genes in seeds of an miR160-resistant mutant mARF10. The downregulation of ARF10, an important factor for auxin signaling, may be a regulatory step to decrease ABA sensitivity in mature seeds and to switch on the germination program (Nonogaki, 2008b).
Jung and Kang (2007) reported the essential role of miR417 in seed germination under the stress. The expression of miR417 was upregulated by salt, dehydration stress, and after ABA treatment. Seed germination of overexpressing miR417 plants was retarded compared to the wild type in the presence of NaCl and ABA. These results imply a negative regulatory role of miR417 during seed germination.
Crosstalk Between Epigenetic and microRNA Regulation of Gene Expression in Terms of Abiotic Stress During Seed Germination
miRNAs are able to indirectly regulate the DNA demethylation process as an adaptive mechanism to stress conditions during seed germination in plants. An excellent example is miR402, which affects seed germination under stress conditions. It targets the DML3 (DEMETER-LIKE 3) gene that encodes a protein involved in DNA demethylation. miR402 is upregulated by various abiotic stresses such as cold, dehydration, and salt. miR402 mutants germinated earlier than the wild type under stress conditions. In seeds of the overexpressing miR402 mutant DML3 showed a decreased level of expression under stress. An analysis of the dml3 and miR402 knock-out and overexpressing lines clearly demonstrated that miR402 is a positive regulator of seed germination in the presence of stress factors. miR402 targets DML3, which consequently maintains DNA methylation within the promoters of negative regulators of seed germination (Kim et al., 2010).
To sum up, it is worth stating that not only the regulation of transcription but also posttranscriptional regulation have a strong impact on the stress response of germinating seeds. miRNA such as miR160 or miR159, which downregulate positive regulators of auxin and ABA, respectively, and chromatin remodeling achieved by PKL or demethylation performed by DML3 were shown to be important for the further seed germination program.
RNA Binding Proteins—Important Components of Posttranscriptional Regulation Machinery in Response to Stress During Seed Germination
Gene expression at the posttranscriptional level can be regulated either directly by RNA-binding proteins (RBPs) or indirectly when RBPs modulate the function of other regulatory factors. The RRM, also known as the consensus sequence type RNA-binding domain, is the most widely found and best-characterized RNA-binding motif (Lorkovič and Barta, 2002). RRMs motif is localized in the N-terminal half of RBPs, and is involved in the recognition and binding of target RNA molecules. Although proteins that contain RRMs in the N-terminal part and a glycine-rich region in the C-terminal half (glycine-rich RNA binding proteins; GRPs) have been extensively described in plants, their role still remains unclear. Sachetto-Martins and others (2000) showed that their mRNA levels increased following exposure to a variety of environmental stresses and phytohormones. Kim and others (2007) investigated the role of GRP2 (glycine-rich RNA binding protein 2) in seed germination under abiotic stress. They showed, using the knock-out insertional grp2 and overexpressing 35S::GRP2 mutants, that GRP2 plays a role as a positive regulator of germination and seedling growth under cold or salt stress. No differences to ABA treatment were observed during seed germination between grp2 mutants and the wild type. This implies that GRP2 acts in an ABA-independent pathway. Kim and others (2007), using proteome analysis, also proved that GRP2 functions as a positive regulator of germination during abiotic stress through the regulation of the expression and activity of various sets of genes. Their analysis revealed a modulation of several mitochondrial-encoded genes by GRP2.
Conclusions and Further Perspectives
Seed development and germination are crucial processes for plant survival. This minireview focused only on recent reports and some aspects of the cooperation between phytohormones in order for Arabidopsis to develop a correct response to abiotic stress during seed germination. Elucidating the complexity of the integral mode of action of different pathways during seed germination will enhance the understanding of abiotic stress response and enable this important step in the plant life cycle to be controlled.
Footnotes
Acknowledgments
The author thanks Prof. Iwona Szarejko for her critical reading of the manuscript and helpful suggestions. This work was supported by the Ministry of Science and Higher Education, Poland (NN304508938).
Author Disclosure Statement
The author declares that no conflicting financial interests exist.