
Abstract
Abstract
Phytohormones are chemical messengers that play a leading role in regulating the vital activity of plants, including transcription, posttranscriptional pre-mRNA splicing, translation, and posttranslational modifications by interacting with specific protein receptors. Plant hormones are synthesized in one tissue and act on specific target sites in other tissues at vanishingly low concentrations. High salinity is one of the main factors limiting Arabidopsis growth and productivity. In this study, phytohormones including abscisic acid, auxin, ethylene, and cytokinin responsive genes regulating salinity stress in Arabidopsis roots were monitored using microarray data. We identified phytohormone responsive genes on the basis of their expression pattern at genomic level at various time points. Using publicly available microarray data, we analyzed the effect of salt stress on the transcription of phytohormone responsive genes. Gene ontology (GO) analysis of phytohormone responsive genes showed their role in important biological processes such as signal transduction, hormone metabolism, biosynthetic process, and gene expression. Gene enrichment terms also reveal that transcription regulator activity is the main class of ABA responsive genes under salinity stress. We conclude that expression of ABA responsive genes involves induction of several transcription factors under salt stress treatment in Arabidopsis roots.
Introduction
Arabidopsis thaliana, a model plant from Brassicaceae family, is used widely for basic research in laboratory because of its small size, short life cycle (about 8 weeks from germination to mature seed), small genome size (125 Mb), and its ability to self-pollinate. Phytohormones have been implicated in diverse processes throughout the life span of Arabidopsis where they control a vast array of plant responses affecting growth and development (Jiang and Deyholos, 2009; Pandolfini, 2009; Stamm and Kumar, 2010; Yu et al., 2004). Phytohormones interact with each other in both complementary and in antagonistic ways to accomplish signaling, protecting plants from abiotic stresses such as drought, salinity, cold, heat, excess light, and biotic stresses such as pathogens (Santner and Estelle, 2009; Schmelz et al., 2003; Singh et al., 2011). For example, literature has clearly established that phytohormone ABA serves as an endogenous messenger in a host of biotic and abiotic stress responses of plants (Adie et al., 2007; Christmann et al., 2006; Hrayama and Shinozaki, 2007; Melotto et al., 2006; Ton et al., 2009). Salinity, one of the most important abiotic stress, is a major threat to agriculture in India and various other countries. Drought and high salinity result in strong increase in plant ABA levels, accompanied by a major change in gene expression and in adaptive physiological responses (Christmann et al., 2007; Priest et al., 2006; Rabbani et al., 2003; Zeller et al., 2009). ABA is not only a stress signal but is also required to fine-tune growth and development under nonstress conditions. ABA also acts together with other phytohormones such as brassinosteroids, gibberellic acids, and auxins in regulating plant growth and development (Achard et al., 2006; Smet et al., 2003; Zhang et al., 2009).
The role(s) of other phytohormones in stress response in plants has also been indicated in scattered reports (Cao et al., 2007; Coba et al., 2008; Ghanashyam and Jain, 2009; Jain and Khurana, 2009). However, a comparative account about how the coordinated functioning of these different plant hormones determines the ultimate response of plants toward stress has not been worked out. In the present article, we have tried to identify the key salinity responsive genes involved in either biosynthesis, metabolism, signaling, or transcription-associated processes that are simultaneously linked with hormone responses in Arabidopsis. We have also tried to identify a set of genes that are common between responses of plant for more than one phytohormone and also tried to find out the metabolic pathways that are commonly altered between salinity and various phytohormones. For this purpose, we have chosen to work with the genome sequence of the most finished plant genome, that is, Arabidopsis, for obvious reasons. Here we focused on gene expression analysis using affymetrix data from publically available database called TAIR AtGenExpress (http://www.arabidopsis.org/portals/expression/microarray/ATGenExpress.jsp) of Arabidopsis roots at 0.5, 1, 3, 6, 12, and 24 h under salinity for four major phytohormones, that is, ABA, auxin, ethylene, and cytokinin-responsive genes from systematic and comprehensive database known as Arabidopsis hormone database (AHD2.0; http://ahd.cbi.pku.edu.cn). Furthermore, we analyzed the functional classification of upregulated and downregulated genes by salinity treatment using DAVID bioinformatics tool. We have also studied the expression analysis of transcriptional regulator activator genes of Arabidopsis and performed clustering using R statistical package to provide evidence of importance of transcription factors under salinity treatments.
Materials and Methods
Identification of phytohormone-responsive genes
Phytohormone-related genes of model organism Arabidopsis thaliana were picked up from online accessible database, that is, Arabidopsis Hormone Database (AHD2.0, http://ahd.cbi.pku.edu.cn) (Jiang et al., 2011). The database aims to provide systematic and comprehensive view of Arabidopsis hormone-related genes (AHRGs). It includes genes involved in the biosynthesis, metabolism, transport, perception, or signaling pathways of eight types of phytohormones. AHD2.0 crosslinks to PPI and KEGG for molecular biochemical studies for Arabidopsis phytohormone. In this regard, we have specifically collected genes for four major phytohormones viz. ABA, auxin, ethylene, and cytokinin only and investigated their relevance to salinity responses in Arabidopsis.
Affymetrix microarray data analysis
For abiotic stresses, such as salt, the .CEL files that contain measured intensities for each probe on the array for roots of Arabidopsis with control and stress replicates were downloaded from TAIR AtGenExpress (http://www.arabidopsis.org/portals/expression/microarray/ATGenExpress.jsp). Affymetrix microarray data analysis of .CEL files of roots of Arabidopsis under salinity stress were performed using Bioconductor (Gentleman et al., 2004) package. Affy bioconductor package was used for affymetrix pre-processing of .CEL files (Gautier et al., 2004). Another bioconductor package, that is, limma (Smyth et al., 2005) was used for assessing differential gene expressions of roots of Arabidopsis under salinity using R statistical package (http://www.r-project.org/). Normalization of data, probe summarization, and background correction was done using MAS5.0 method (Pepper et al., 2007) and absolute intensity values from MAS5.0 were transformed to log2 (logbase2) values, that is, fold change (log2fc). For each experiment, the log2 intensities for individual probe sets were averaged across two replicates for treatment and control. The analysis was performed for six time point, that is, 0.5, 1, 3, 6, 12, and 24 h the roots of Arabidopsis under salinity stress and control condition. Among the six salt-stress experiments in root tissues carried out at six time points (i.e., 0.5, 1, 3, 6, 12, and 24 h), if log2fc ≥1.0 and significant p-value (≤0.01) then the set of genes was considered to be upregulated, whereas if log2fc ≤1.0 and significant p-value (≤0.01), the corresponding gene array was considered to be downregulated. Likewise, up- and downregulated genes under salinity stress were identified at each given time scale. The scripts and programs were written in R statistical package, perl, and shell on a linux-based machine.
Identification of salinity-regulated phytohormone-responsive genes in Arabidopsis
Phytohormone-responsive genes from ABA, auxin, ethylene, and cytokinin (already fetched from AHD2.0 database) were mined from the set of upregulated and downregulated genes under salinity stress identified using affymetrix microarray analysis at six time scale, that is, 0.5, 1, 3, 6, 12, and 24 h. The mined phytohormone-responsive genes are said to be either upregulated or downregulated at given time scale series under salinity stress in roots of Arabidopsis. The gene expression data has been represented in the form of heatmap using gplots package of R. Hierarchical clustering method has been used for the clustering the genes. The scripts and programs were written in R statistical package, perl, and shell.
Functional characterization of differentially regulated genes
Arabidopsis phytohormone responsive genes that are regulated (up/downregulated) under salinity stress identified from aforesaid method were then processed for functional characterization. The classification of phytohormone responsive genes into group of similar function is powerful approach to understand gene regulation by different phytohormones at the genomic level. We performed a functional classification based on Gene Ontology (GO) enrichment term using Database for Annotation, Visualization, and Integrated Discovery, DAVID (Dennis et al., 2003; Huang et al., 2007). Analysis was performed for phytohormone-responsive genes under salinity stress belonging to three GO terms, that is, biological process (BP), molecular function (MF), and cellular component (CC). The classification is based mainly on the GO enrichment terms with significant false discovery rate (FDR) and significant p-value. Arabidopsis phytohormone-responsive genes under salt stress with significant BP, MF, and CC, respectively (FDR ≤0.05) and (p-value ≤0.05) were considered finally for further analysis.
Expression analysis of transcription-related genes
Based on the DAVID GO classification of Arabidopsis phytohormone-responsive genes under salinity, ABA genes were found to be significantly enriched in molecular function called transcription regulatory activity. Functional annotations of the transcription regulatory activity genes revealed that most of them belong to transcription factors. Detailed analysis of this group of genes (transcription regulatory activity) was performed by studying their expression patterns, that is, log-transformed fold change (log2fc) values by the affymetirx microarray method and plotted in the form of heatmap using gplots package of the R statistical package. Clustering was performed using the hierarchical clustering method of the R package. All the scripts were written in R to generate the heat maps.
Results
Analysis of salinity-responsive phytohormones-related genes
Our datasets taken from AHD2.0 database (Jiang and Guo, 2010) includes a total of 360 ABA-responsive genes, 387 auxin responsive, 255 ethylene responsive, and 98 cytokinin regulated genes. The results of this analysis are represented in the form of venn diagram using venny software (http://bioinfogp.cnb.csic.es/) as shown in Figure 1. There are 273, 306, 167, and 80 unique genes corresponding to ABA, auxin, ethylene, and cytokinin, respectively. ABA share 51, 60, and 7 genes with auxin, ethylene, and cytokinin, respectively. Similarly, in Auxin, 52 and 10 genes were found to be common with ethylene and cytokinin, respectively. In contrast, only five genes were found to be common between ethylene and cytokinin. It could be concluded that ABA, auxin, and ethylene shared a considerably high number of common genes among them. However, the response of cytokinin was not found to be overlapping with any other hormone studies here.
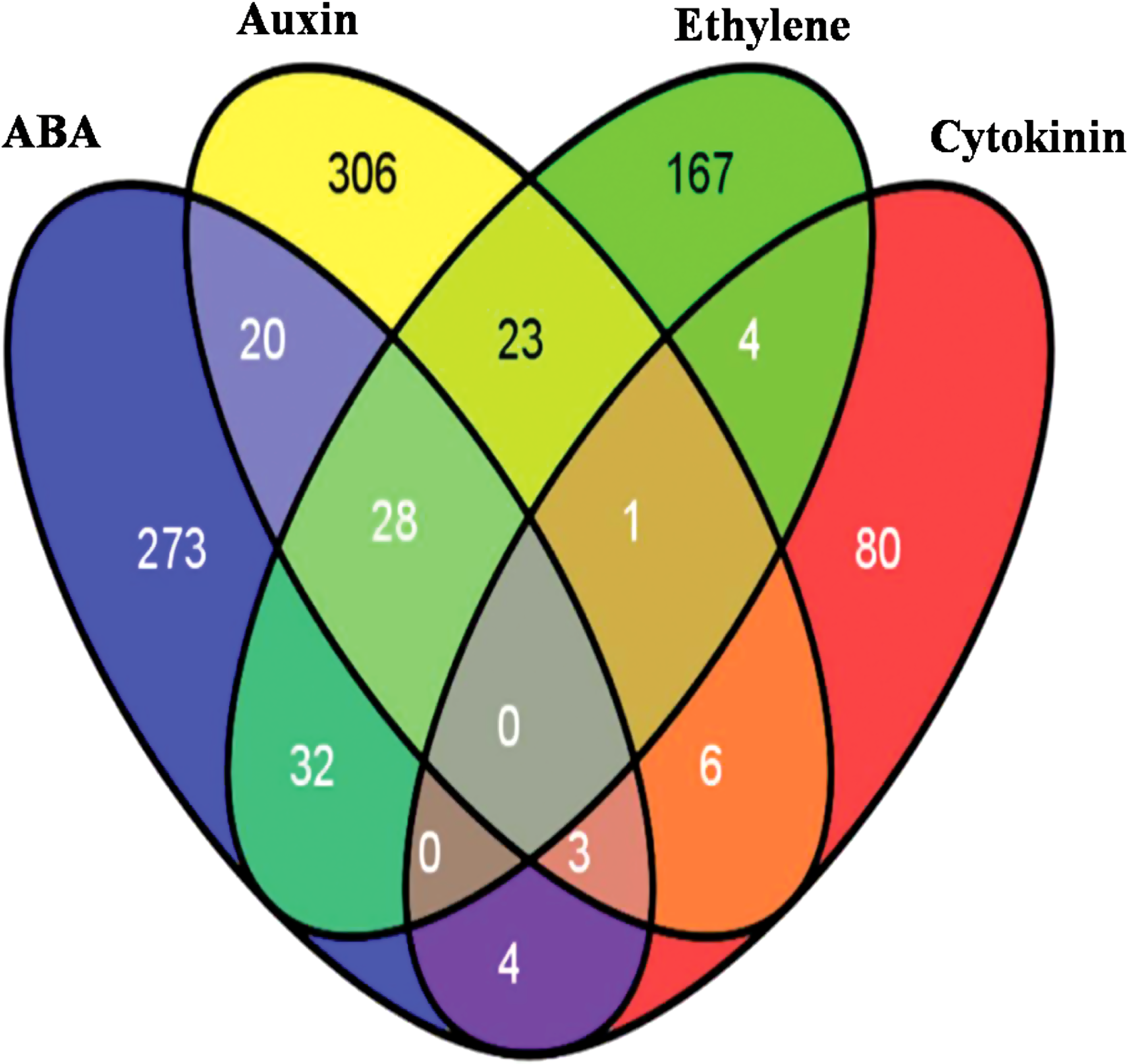
Venn diagram showing the total number of genes that are phytohormone responsive (based on AHD2.0) generated using venny software. There are 360, 387, 255, and 98 genes for ABA, auxin, ethylene, and cytokinin. 273, 306, 167, and 80 genes are unique in ABA, auxin, ethylene, and cytokinin. There are 51, 60, and 7 genes of ABA hormone common to auxin, ethylene, and cytokinin, respectively. There are 52 and 10 auxin responsive genes common to ethylene and cytokinin. It is clear that only five genes are common between ethylene and cytokinin.
Surprisingly, no gene common to all the four phytohormones emerged from this analysis. Analysis of the common phytohormone responsive genes between ABA, auxin, and ethylene revealed most of them to be downstream signaling components of hormones such as transcription factors like MYBs, CCA1, LCL1, LHY, TRB1, TRB2, and RVE2. It showed that signaling of different phytohormones leads to the alteration of gene expression through activation/repression of various transcription factors.
Based on the above analysis, we wanted to mine out the genes that are responsive to salinity stress in addition to being phytohormone responsive. AtGenExpress has been used to obtain affymetrix data for genes that are salinity stress responsive in root tissues. Using affy and limma packages of Bioconductor, the (log2fc) values were generated and processed to identify phytohormone-responsive genes under salinity stress (at different time points, that is, 0.5, 1, 3, 6, 12, and 24 h). Salinity responsive genes that are either upregulated (log2fc ≥1.0, p≤0.01) or downregulated (log2fc ≤−1.0, p≤0.01) by phytohormones were picked up for further analysis. The total number of phytohormones responsive genes that showed up- or downregulation at various time points in response to salinity stress is presented in Table 1. Total number of genes that showed upregulation in response to salinity (at any time point) in the case of ABA was found to be highest, that is, 129, indicating the contribution of this hormone in determining the ultimate response of plants towards salinity stress in roots of Arabidopsis. Similarly, the number of ABA responsive genes that showed downregulation in response to salinity (at any time point) were found to be 37. This trend was reported to be least in case of cytokinin where the corresponding 16 number of genes was found to be upregulated and 14 downregulation. For auxin, 89 number of genes were found to be upregaulated and 55 genes downregulated in response to salinity in Arabidopsis roots. Ethylene could upregulate a total of 74 genes and downregulate 25 genes under salinity stress.
“Very early changes” in transcriptome of Arabidopsis roots under salinity stress
Various phytohormone-related genes were found to be up/downregulated in roots of Arabidopsis during very early response to salinity, that is, 0.5 h after salt treatment (Fig. 2). There are total of 51 phytohormone-related genes that are upregulated and only 5 downregulated under salinity at this stage, out of which some are still not functionally annotated in Arabidopsis. The number of genes upregulated under these conditions was found to be 22 for ABA, 10 for auxin, 17 for ethylene, and 2 for cytokinin respectively (Table 1). Expression behavior of these genes is shown in (Fig. 2) as a histogram heat map. This response is dominantly represented by presence of ethylene-responsive factors such as AtERF4, AtERF8, AtERF13, and AtERF104, etc. It also includes six genes coding for ACC synthase, nine for terpene synthase and one for C-repeat/DRE binding factor. The transcription factors such as WRKY (WRKY33 and WRKY18), AtMYB (MYB15, MYB41, and MYB51), etc., were among the other genes that are highly induced during salinity stress. Similarly, downregulated genes included kelch repeat proteins, Dof-type zinc-finger domain containing genes, U-box superfamily, PYR1-like 6, etc.
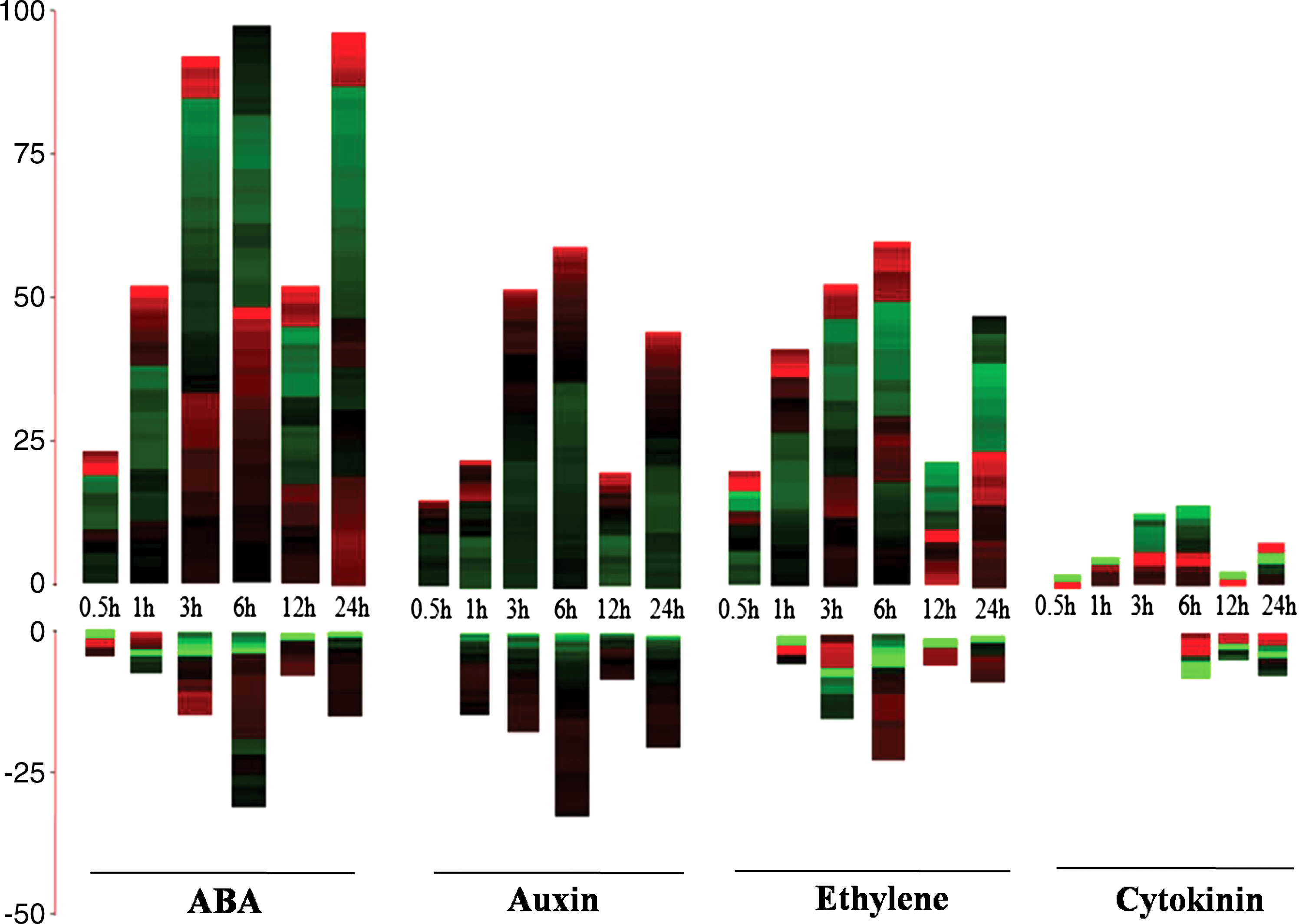
Gene expression pattern of ABA, auxin, ethylene, and ctyokinin phytohormones responsive genes in A. thaliana under salt treatment at 0.5, 1, 3, 6, 12, and 24 h, respectively. Upregulated genes (log2fc ≥1.0, p≤0.01) are shown in upward bars and downregulated genes (log2fc ≤−1.0, p≤0.01) are shown in inverted bars. The data is shown in the form of histogram heat maps where y-axis is the total number of genes upregulated and downregulated at particular time point and x-axis shows six time intervals, respectively. Red black and green color shows maximum, moderate, and minimum levels of gene expression.
“Early responses” in transcriptome of Arabidopsis roots under salinity stress
A total of 112 and 24 hormone-related genes respectively are highly upregulated and downregulated at 1 h of salinity stress. These include key regulators in abiotic stress responses such as low-temperature-responsive protein78 (RD29A) and C-repeat/DRE binding factor (DREB). Genes functioning in ABA synthesis and signal transduction also appeared in this list, including ABI1, PP2CA, and NCED3. Transcription factors especially MYBs, NACs, redox responsive 1, and WRKYs were also upregulated in roots under salinity stress conditions. Similarly, the downregulated genes under this category includes PYR1-like 6, UDP-glycosyltransferase, SAUR-like auxin-responsive protein family, P-loop containing nucleoside triphosphate hydrolases, TSPO, VQ motif-containing protein, RECEPTOR-like protein kinase, auxin-responsive GH3 family protein that acts as a bifunctional modulator in auxin and salicylic acid (SA).
Transcriptome changes after 3 h of salinity stress in roots of Arabidopsis
The hormone-related genes that are strongly upregulated by salt stress in roots at the 3-h treatment in various phytohormones were found to be 200 in number (Fig. 2). This include transcription factors such as basic helix-loop-helix (bHLH), NACs, MYBs (MYB15, MYB77, MYB74, MYB96, and MYB108). Lipid transfer proteins (LTP3, LTP4, glycolipid transfer protein, phospholipase C1), calcium-dependent protein kinase 32, stress induced proteins (PP2C, DREB, AP2, AtEFRs, ACC synthase, ACS6, ACS11, and ACS2), calneurin B-like protein 1, DNA binding superfamily protein, cytochrome P450, dehydrin family protein, RD29A, NAT, pseudouridine synthase, serine carboxypeptidase-like 30, RCC1, MAPK, homeobox 12 were also found to be strongly upregulated (with more than threefold change). Several phytohormone-responsive genes were found to be downregulated, that is, 45 by salt stress, which include transposable element gene, cysteine-rich secretory proteins, zinc-finger (C2H2 type), SAUR-like auxin-responsive protein family, etc.
Transcriptome changes after 6 h of salinity stress in roots of Arabidopsis
After 6 h of salinity stress, relatively more hormone-related genes were found to be up- or downregulated compared to other durations of stress, studied here (Table 1). In total, there are 221 upregulated and 95 downregulated genes in response to four phytohormones at the 6-h salinity treatment. Upregulated genes included abiotic stress responsive genes such as RD29A and DREB2A. Genes functioning in ABA synthesis and signal transduction also appeared in the list, including ABI1, PP2CA, and NCED3. Transcription factors such as bHLH, MYB2, MYB 77, MYB 108, etc., were also reported. Lipid transfer proteins, KIN1, KIN2, NAT, AFP2, UDP-glucosyl transferase, Stress-induced protein, salt tolerance zinc-finger, phosphatase 2C5, and alpha/beta-Hydrolases superfamily protein are strongly expressed by more than threefold. Downregulated genes in this list includes MYB28, MYB59, arabinogalactan-protein, tonoplast intrinsic protein, cytochrome P450, S-adenosyl-L-methionine-dependent methyltransferases, Sec14p-like phosphatidylinositol transfer family protein, wound-responsive family protein, CAP, diacylglycerol kinase 6, peroxidase superfamily protein, tryptophan aminotransferase-related 2, disease resistance-responsive, nodulin MtN21, actin depolymerizing factor 8 and Auxin-responsive GH3.
Transcriptome changes after 12 h of salinity stress in roots of Arabidopsis
It was noted that the number of upregulated and downregulated phytohormone-related genes in roots under salt treatment at 12 h declined compared to that observed after 6 h of salinity stress. There are around 93 upregulated and 22 downregulated genes, including NCED3, PP2CA, dehydrin, RD29A, homeobox 12, NAT, cytochrome 450, NCED2, MYBs (MYB2, MYB108, MYB121, MYB102, MYB79, MYB15), UDP-glucosyl transferase 71B6, stress induced protein, ABI, NAC, KIN1, KIN2, salt tolerance zinc-finger, TSPO, AP2, bHLH, VQ motif-containing protein, redox responsive transcription factor 1, DREB1, GAST1, HSP20, Auxin-responsive GH3, and AtERF 13. Downregulated genes in this category include CAP, defensin-like protein, iron-regulated transporter 1, MYB 28, histidine phosphotransfer protein 6, etc.
“Late responses” in transcriptome of Arabidopsis roots under salinity stress
There are a total 182 genes that are upregulated and 51 genes that are downregulated under these set of conditions (Table 1). These include NCED3, PP2CA, RD29A, CAP160, NCED5, NAT, Zin finger AN1, Lipid transfer protein4, MYBs (MYB108, MYB121, MYB77, MYB41), UDP-glucosyl transferase71B6, Stress-induced protein, NAC, P-loop containing nucleoside triphosphate hydrolases superfamily protein, salt tolerance zinc-finger, TSPO, DREB2, lectin receptor kinase, AtERF 1, alpha/beta-Hydrolases superfamily protein, cupredoxin superfamily protein, gamma-glutamyl transpeptidase 1, cytochrome P450, 1-amino-cyclopropane-1-carboxylate synthase 8, bHLH, redox responsive transcription factor 1, AP2, xyloglucan endotransglycosylase 6, HSP 23.6, DREB 1, DREB2, ATP binding microtubule motor family protein, cysteine-rich RLK, Subtilase family protein, MYB 102, WRKY 28, MYB 79, SAUR-like auxin responsive, tolB protein-related, PPR, HSP 20, ACC synthase 6, Auxin-responsive GH3, and ERF1 are some of them.
Functional classification of phytohormone-related salinity-regulated genes in roots of Arabidopsis
To identify possible GOs of upregulated and downregulated phytohormone-related genes in response to salt treatment, their functional characterization was studied using Database for Annotation, Visualization, and Integrated Discovery, DAVID (http://david.abcc.ncifcrf.gov/). GO enrichment analysis was performed using default parameters and the significant results with (FDR ≤0.05) were further processed for the analysis. Supplementary Table S1 lists the results of the GO analysis for the upregulated probe sets (the downregulated probe sets showed no significant enriched GO terms).
Out of a total of 129 unique salinity-responsive upregulated probe sets for ABA, 33 featured GO term annotations in biological processes, 7 in molecular function, and 1 featured in the cellular component (Supplementary Table S1). The distribution of enriched GO terms showed several noteworthy findings. The significantly enriched GO terms included “abscisic acid mediated signaling” (GO:0009738; FDR=1.03E-026; p-value=7.81E-030), “hormone-mediated signaling” (GO:0009755; FDR=1.58E-011; p-value=1.20E-014), “response to desiccation” (GO:0009269; FDR=1.82E-007; p-value=1.38E-010), “abscisic acid metabolic process” (GO:0009687; FDR=2.86E-007; p-value=2.17E-010), “intracellular signaling cascade” (GO:0007242; FDR=4.95E-007; p-value=3.76E-010), “regulation of RNA metabolic process” (GO:0051252; FDR=1.10E-005; p-value=8.36E-009), “hyperosmotic salinity response” (GO:0042538; FDR=0.00284981; p-value =2.17E-006); details are shown in Supplementary Table S1). Statistically significant biological processes involved in ABA responsive genes under salinity stress are shown in Supplementary Figure S1A/S1B.
GO terms enriched in molecular function and the cellular component are shown in Supplementary Table S1. The most significantly enriched GO term in molecular function was “transcription factor activity” (GO:0003700; FDR=1.39E-012; p-value=1.16E-015), “transcription regulator activity” (GO:0030528; FDR=2.92E-011; p-value=2.34E-014), “DNA binding” (GO:0003677; FDR=4.89E-007; p-value=3.92E-010). The only enriched GO term in the cellular component was “nucleus” (GO:0005634; FDR=8.50E-008; p-value=8.42E-011).
Similarly, 89 unique upregulated auxin responsive genes under salinity stress, 11 probe sets featured GO term annotations in biological processes, 2 in molecular function, and 1 enriched in the cellular component (nonsignificant). The significantly enriched GO terms included “response to chitin” (GO:0010200; FDR=2.20E-008; p-value=1.55E-011), “auxin-mediated signaling pathway” (GO:0009734; FDR=2.18E-006; p-value=1.53E-009), “hormone-mediated signaling” (GO:0009755; FDR=5.70E-005; p-value=4.02E-008), “regulation of transcription, DNA-dependent” (GO:0006355; FDR=2.21E-004; p-value=1.56E-007) as presented in Supplementary Figure S1A/S1B). The significantly enriched GO term in molecular function was “anthranilate synthase activity” (GO:0004049; FDR=6.09E-005; p-value=5.33E-008), “indole-3-acetic acid amido synthetase activity” (GO:0010279; FDR=0.00709161; p-value=6.21E-006). Statistically significant biological processes involved in auxin responsive genes under salinity are shown in Supplementary Table S1. There was no significantly enriched GO term found in the cellular component.
Analysis of ethylene responsive genes has indicated 74 unique upregulated genes, 25 probe sets enriched in biological processes, 2 in molecular function, and 3 featured in the cellular component. The significantly enriched GO terms included “ethylene-mediated signaling pathway” (GO:0009873; FDR=1.71E-016; p-value=1.37E-019), “two-component signal transduction system” (GO:0000160; FDR=1.98E-014; p-value=1.58E-017), “hormone-mediated signaling” (GO:0009755; FDR=3.61E-012; p-value=2.90E-015), “regulation of macromolecule biosynthetic process” (GO:0010556; FDR=2.50E-011; p-value=2.00E-014), “regulation of gene expression” (GO:0010468; FDR=3.18E-011; p-value=2.54E-014). The significantly enriched GO term in molecular function was “anthranilate synthase activity” (GO:0004049; FDR=3.43E-011; p-value=4.27E-014). Enriched GO term in the cellular components involves, “nucleus” (GO:0005634; FDR=6.33E-013; p-value=6.24E-016;), “intracellular membrane-bounded organelle” (GO:0043231; FDR=0.00121348; p-value=1.28E-006), “intracellular organelle,” etc. Supplementary Figure S1C shows the statistically significant biological processes involved in ethylene responsive genes under salinity.
Transcription factors as key regulator of salinity response in roots of A. thaliana
Gene enrichment analysis showed that the transcription regulator activity (28%) was significantly represented in molecular function associated with ABA response. Similarly, the corresponding value for DNA binding activity was noted to be 27%, transcription factor activity 27%, transcription activator activity with 7%, serine/threonine phosphatase activity (6%) as shown in Figure 3). Transcription factors are well known to be primarily involved in the initiation stage of RNA transcription and they are the key factors that regulate gene expression. A large number of transcription factors in the root tissue of Arabidopsis responded to the salt treatment. Table 2 shows the categorization of Arabidopsis transcription regulator activity genes with upregulation under the salt stress, including such key regulatory gene families involved in responding to abiotic stress as WRKY2, WRKY25, and WRKY33, AP2, NAC, MYBs (MYB51, MYB77, MYB41, MYB49, MYB99, MYB108, MYB112, MYB121, MYB94, MYB7), basic helix-loop-helix (bHLH), Bzip, and heat-shock transcription factor genes.
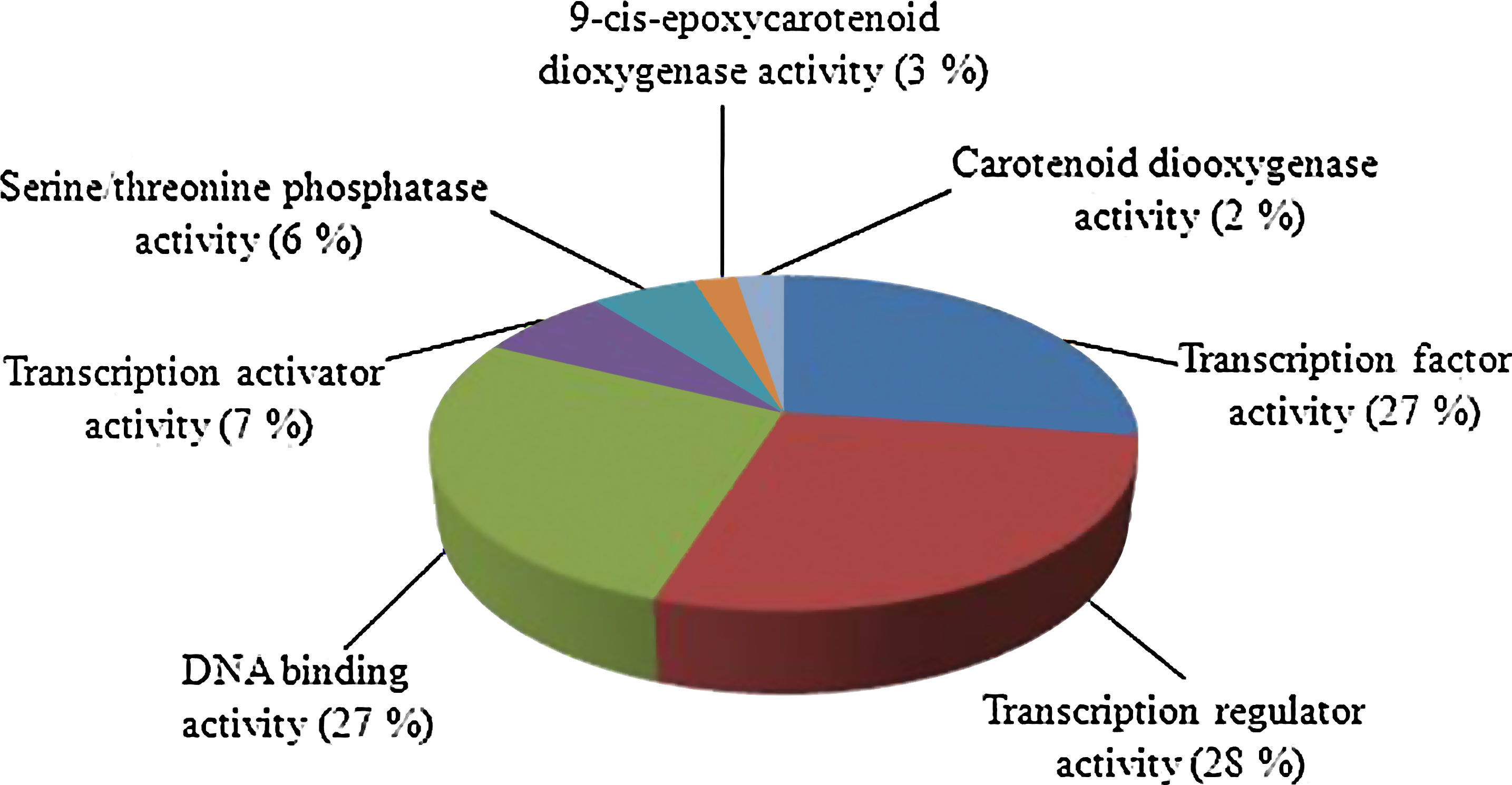
Pie chart showing molecular function assigned to A. thaliana ABA responsive genes under salinity stress. Major players involved proteins having transcriptional regulatory activity (28%), DNA binding activity (27%), and transcription factor activity (27%).
Figure 4 represents hierarchical clustered heatmap of the transcriptional regulator activity for ABA responsive genes in roots of Arabidopsis. There are four main clusters formed based on the gene expression after hierarchical clustering cluster 1 (C1), cluster 2 (C2), cluster 3 (C3), and cluster 4 (C4). Cluster 1 includes genes that highly expressed in roots of Arabidopsis during the late hours under salt stress. Genes such as AP2, MYB108, zinc-finger, NAC, MYB 121 are strongly upregulated at 1, 3, 6, 12, and 24 h salt treatment in roots. C2, includes genes like MYB41, MYB74, and MYB102 and homeobox12 that are strongly upregulated during all time intervals of salt stress in root tissue. Cluster 3, contains abiotic stress related genes, that is, C-Repeat/DREB 2 and also include transcription factors genes like MYB77, WRKY33, WRKY25, and bHLH that are strongly expressed during the immediate, early, and 3 and 6 h of salt treatment. It was noticed that these genes showed less expression at 12 h of salt treatment, whereas at 24 h, these genes are highly expressed in roots again. Cluster 4, possess mainly transcription factor genes such as MYB96, MYB 51, some signaling pathway genes, ethylene responsive element factors 4 (ERFs 4), and TAZ domain protein 2 that are highly upregulated during immediate 0.5 h, early 1 h, and 3 h in response to salt stress.
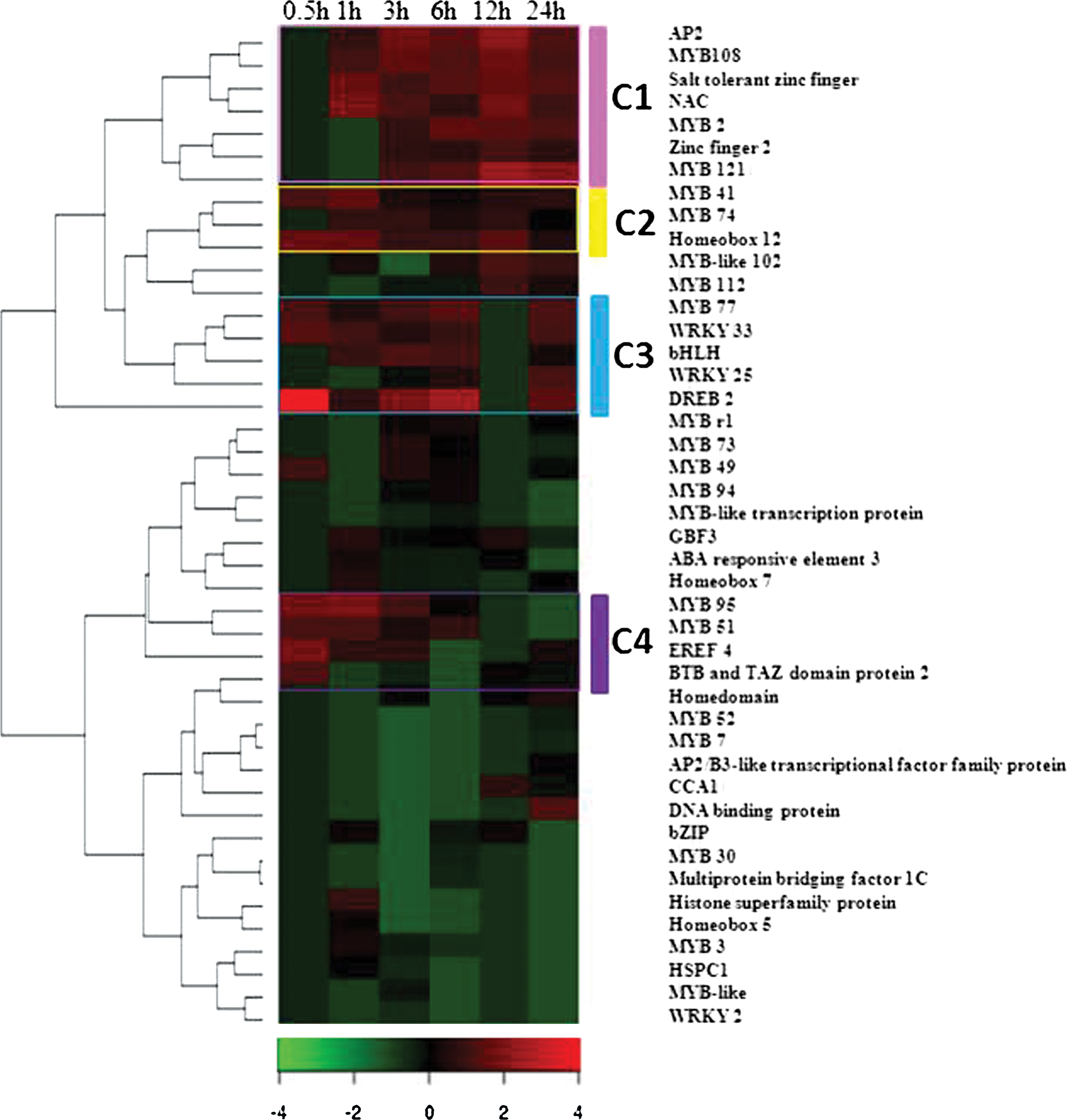
Heat map of the expression analysis of ABA responsive genes under salt stress with transcriptional regulatory activity. Hierarchical clustering method was used for clustering. Cluster C1 (pink), C2 (yellow), C3 (aqua), and C4 (purple) are the major clusters. The heatmaps was made using gplots package of open source R software. The scale shows the Z-score, which is defined as “actual value” minus the mean of the group divided by standard deviation.
Discussion
Salinity is a major threat to crop productivity, and efforts are going worldwide to improve the stress tolerance in crop plants (Ashraf et al., 2008). In this regard, several genes have been reported in the literature that are stress responsive, and engineering of a few of them has resulted in improved tolerance in plants (Ashraf and Aishaa, 2009; Pandey et al., 2003; Prasad et al., 2000; Singla-Pareek et al., 2003). Phytohormomes contribute a large amount in controlling the plant responses toward their microenvironment (Bari and Jones, 2009). Interactions between the various genes that are either responsive to phytohormones or salinity stress have not been studied in detail in any plant at a single platform. We wanted to find out the commonality between the phytohormone-responsive genes and those that are regulated by salinity stress. With this view, we have chosen to work on the data and information available from public databases with respect to the model plant—Arabidopsis thaliana.
ABA serves as an endogenous messenger in a host of biotic and abiotic stress responses of plants (Adie et al., 2007; Christmann et al., 2006; Hirayama and Shinozaki, 2007; Melotto et al., 2006; Ton et al., 2009). Auxins plays a critical role during plant growth including apical dominance, tropic responses, vascular development, organ patterning, flower development, and fruit development (Ghanashyam and Jain, 2009). A large number of auxin-responsive genes were also found to be differentially expressed under various biotic, abiotic stress conditions, and signaling pathways (Choeng et al., 2002; Dowd et al., 2004; Ghanashyam and Jain, 2009; Jain and Khurana, 2009; Navarro et al., 2006; Wang et al., 2007), supporting a possible crosstalk between ABA and stress response.
Cytokinin, another phytohormone, induces a variety of physiological events, including cell division, chloroplast development, and shoot formation (Mok and Mok, 1994; Skoog and Miller, 1957). Some reports have also shown the role of cytokinin under salinity that acts as major regulating leaf growth, senescence, shoot growth, and fruit yield in the tomato (Albacete et al., 2010; Ghanem et al., 2011). Salt stress-induced changes of cytokinins in maize and pea plants suggest that NaCl treatment reduces the roots, shoots, and in leaf growth (Atanassova et al., 1997). Recently, a cytokinin receptor homolog, MsHK1, from Medicago sativa root, was induced by salt stress in alfalfa roots, leaves, and nodules (Coba et al., 2008). Ethylene has long been regarded as a stress hormone (Morgan and Drew, 1997). It is not only involved in plant growth and development processes including seed germination, leaf, and flower senescence, and fruit ripening (Abeles, 1992), but also acts as signal in biotic stress such as pathogen defense and abiotic stresses such as wounding, ozone, and salt (Abeles, 1992; Cao et al., 2007; O'Donnell et al., 1996; Vahala et al., 2003). Recent studies have demonstrated that exogenous 1-aminocyclopropane-1-carboxylic acid (ACC), an ethylene precursor in Arabidopsis, confers salt tolerance (Cao et al., 2007). The upstream enzyme in the ACC pathway, ripening-specific-1-aminocyclopropane-1-carboxylate synthase (ACS), has been correlated with long shelf life of fruits (Wang et al., 2009).
Identification of phytohormone-related salinity responsive genes in Arabidopsis genome
Various plant hormones such as ABA, auxin, ethylene, cytokinin, JA, and SA regulate growth and development and mediates plant adaptation to various environmental stresses (Santner and Estelle, 2009). Our study demonstrates that phytohormone-responsive genes from ABA, auxin ethylene, and cytokinin are involved in response to salt stress in Arabidopsis roots. We used the expression data available from microarrays, which is becoming a routine tool for the global analysis of gene expression in several organisms (Wullschleger and Difazio, 2003). It is often irresistable to correlate the expression data with biological functions of genes that show differential expression patterns. To investigate gene expression on genomic scale with stringent significance level of p-value<0.01, we identified ∼3,000 genes differentially regulated by four major phytohormones like ABA, auxin, ethylene, and cytokinin. The expression data can be grouped into related groups using hierarchical clustering (Eisen et al., 1998), self-organizing maps (Tamayo et al., 1999), singular value decomposition (Alter et al., 2000) and evolutionary clustering (Ma et al., 2006). We used the hierarchical clustering method and then visualized the data in the form of heat maps using the R statistical package. Clusters of genes with a synchronized expression pattern suggest that genes share regulatory elements in their promoters (Bucher, 1999). Our data indicates that there are phytohormone-responsive genes that are differentially regulated under salinity stress (Table 2 and Fig. 2). Around 415 hormone-responsive genes were found to be differentially regulated under salinity stress in the roots of Arabidopsis, indicating the overlapping nature of gene regulatory networks.
Salinity response is a complex trait affecting several biological processes
Microarray is a powerful method to measure the gene expression of thousands of genes simultaneously. Because the clustering is a classical way to classify the genes that show similar expression pattern, this makes it hard to determine if the identified significant clustered represents different functional outcomes or rather a redundant view of the same biological processes (Lu et al., 2008). GO (Ashburner et al., 2000) analysis serves as a structured way to arrange genes of specific informative groups. We used DAVID to functionally annotate differentially regulated gene lists under salt treatment with similar functions with stringent significant FDRs and a p value <0.05 (Yao et al., 2011). This analysis indicated that ABA-responsive genes are involved in signal transduction (Raghavendra et al., 2010), dessication (Goldgur et al., 2007), metabolism (Millar et al., 2006), cold acclimation (Xue-Xuan et al., 2010), transcription regulation (Busk and Pages, 1998), hyperosmotic salinity response etc. Auxin genes are also involved in response to chitin (Mathesius et al., 1998), signaling pathways (Quint and Gray, 2006), metabolism (Kerk et al., 2000), and transcription (Tiwari et al., 2003). Ethylene responsive genes play an important role in signaling (Sakai et al., 1998), gene expression (Zegzouti et al., 1999), biosynthesis (Wang et al., 2002), and ripening (Zegzouti et al., 1999). This data suggest toward a highly complex nature of salinity and hormone-responsive gene networks operative in roots of Arabidopsis (Supplementary Table S1).
Transcription factors appear to be key player in hormone-mediated salinity response
Arabidopsis has 1500 transcription factors (TFs) (Ratcliffe and Riechmann, 2002) in its genome. These TFs can be classified into several families based on the structure of their binding domains. Members of the MYB, MYC, ERF, bZIP, and WRKY transcription factor families have already been implicated in the regulation of stress responses in plants (Schwechheimer et al., 1998; Singh et al., 2002). Therefore, TFs can serve as an attractive targets for the purpose of gene regulation and manipulation of the regulatory elements, beneficial under abiotic stresses. Most of drought and salinity stress-induced genes have been identified and analyzed in Arabidopsis, using genomewide microarray analyses (Denby and Gehring, 2005). Our analysis of ABA-responsive genes under salinity has shown that transcription factors are the best-characterized genes that are differentially regulated in Arabidopsis roots under salinity stress (Table 2). These included members such as WRKYs (Jiang and Deyholos, 2006. 2009; Ma et al., 2006), DREB (dehydration-responsive binding protein) genes (Agarwal et al., 2006; Ma et al., 2006), MYBs (Nagaoka and Takano, 2003), bZIP encoding genes (Uno et al., 2000), plant-specific NACs (Kim et al., 2007). Despite playing role in abiotic and biotic stress conditions, these transcription factors involved in regulatory mechanisms, developmental processes, stress signaling, and genes related to several metabolism (Berrocal-Lobo and Molina, 2004; Jakoby et al., 2002). Systematic analysis of Arabidopsis transcription factor families can lead to the identification of candidate genes that have the potential to improve tolerance to environmental stresses in crop plants.
Conclusions
The role of phytohomormones in abiotic stresses derived from previous studies have provided us knowledge to identify differentially regulated genes under salt stress in Arabidopsis roots. The microarray platform was used with main goal of identifying differentially regulated phytohormone-responsive genes under salinity stress treatment. This has helped us to draw a complete picture to establish a relationship between phytohormones and salt stress, which may serve as new paradigm to explore more genes to engineer crop plants for improved tolerance towards salinity stress. Functional annotation of genes lists using GO term elucidating a systematic classification of the phytohormone-responsive genes in various biological process, molecular function, and cellular component showed their role(s) in vital activity in plant cells. This can also be extended to find pathways associated with these genes, to study how various phytohormone-responsive genes crosstalk with each other. The detailed analysis of transcription factors identified in this work may help in dissecting out the “candidate genes” for improving tolerance of plants towards salinity stress employing the tool of genetic engineering.
Footnotes
Acknowledgments
The authors thank Jawaharlal Nehru University (JNU), Department of Biotechnology (DBT), Government of India for financial support in the lab. Khushwant also thanks the Department of Biotechnology for Junior Research Fellowship.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.