
Abstract
Abstract
Although soy phytoestrogens have been postulated to exert a protective effect against breast cancer, the attendant mechanisms, in particular epigenetics underpinnings, have remained elusive. We investigated the putative effects on DNA methylation by two naturally occurring isoflavones, genistein and daidzein, in a study of the BRCA1 and BRCA2 oncosuppressor genes in breast cancer cell lines (MCF-7, MDA-MB 231, and MCF10a). A demethylant agent, the 5-azacytidine, and a methylant, the budesonide, were used as treatment controls. DNA methylation of BRCA1 and BRCA2 was investigated with methylated DNA immunoprecipitation coupled with PCR. In parallel, protein expression was determined by Western blot, immunohistochemistry, and confocal microscopy. Our results suggest that treatment with 18.5 μM Genistein or 78.5 μM Daidzein might reverse DNA hypermethylation and restore the expression of the oncosuppressor genes BRCA1 and BRCA2. 5-Azacitydine also enhanced the reexpression of these genes while budesonide had an opposite effect. To the best of our knowledge, these observations, while requiring replication, provide new evidence on potential epigenetic mechanisms by which genistein and daidzein might contribute to regulation of the BRCA1 and BRCA2. Future studies are warranted on whether the demethylating effect of genistein and daidzein is global or focused on select candidate genes.
Introduction
In this study, we focused on putative epigenetic effects of the two naturally occurring isoflavones—genistein and daidzein—on DNA methylation. Epigenetics is the study of mitotically and/or meiotically heritable changes in gene expression that are not due to alterations in the DNA sequence (Rossetto et al., 2010; Wu and Morris, 2001). Notably, DNA methylation is one of the most studied and well-understood epigenetic mechanism. In humans, DNA methylation occurs in cytosines that precedes guanines, called CpG dinucleotides, via addition of a methyl group on the 5′ position of the cytosine ring to form 5-methylcytosine. CpG sites are not randomly distributed in the genome; instead, there are CpG-rich regions known as CpG islands, which cover the 5′ end of the promoters of many genes. These islands are not usually methylated in normal cells, whereas CpG sites found in repetitive genomic sequences are highly methylated, presumably to prevent chromosomal instability, translocations, and gene disruption caused by the reactivation of transposable DNA sequences. In tumors, this methylation pattern has a trend that is inverted: there is a hypomethylation of the global genome, with a hypermethylation of CpG islands in the promoter regions of tumor-suppressor genes (Tost, 2010). For example, hypermethylation of the tumor suppressor BRCA1 has been found in sporadic breast cancer cases (Catteau et al., 1999; Dobrovic and Simpfendorfer, 1997; Esteller et al., 2000; Hedenfalk et al., 2001; Mancini et al., 1998; Rice et al., 1998, 2000; Wessels et al., 2002). BRCA2 promoter methylation has also been reported in sporadic breast cancer cases (Cucer et al., 2008).
As a growing number of studies show effects of soy phytoestrogens on DNA methylation (Day et al., 2002; Dolinoy et al., 2006; Fang et al., 2007; King-Batoon et al., 2008; Li and Tollefsbol, 2010; Lyn-Cook et al., 1995; Majid et al., 2009; Qin et al., 2009), the protective effects of soy isoflavones on breast cancer could be due, at least in part, to epigenetic effect on DNA methylation. On the other hand, the putative effects of the common soy phytoestrogens genistein and daidzein on BRCA1 and BRCA2 oncosuppressor gene methylation and the attendant downstream consequences on their expression have not been studied in detail in breast cancer cells. We therefore undertook this study to examine changes in DNA methylation of the CpG islands in the promoters of BRCA1 and BRCA2 in breast cancer cells following exposure to these soy phytoestrogens.
Materials and Methods
Cell Lines
MCF-7 and MDA-MB-231 breast tumor cell lines came from a pleural effusion of patients with invasive breast carcinoma (Cailleau et al., 1974; Soule et al., 1973). The MCF-10a cell line was established from the breast tissue of patient with fibrocystic breast disease (Soule et al., 1990). All three human cell lines were provided by the American Type Culture Collection (ATCC, Rockville, MD).
MCF-7 was cultured in RPMI 1640 media supplemented with 2 mM L-glutamine (Invitrogen, Carlsbad, CA), 20 μg/mL gentamicin (Panpharma, Fougères, France), 10% fetal bovine serum (Invitrogen), 0.04 UI/mL insulin (Novo Nordisk, Bagsvaerd, Denmark) in a humidified atmosphere at 37°C containing 5% CO2. This cell line has a positive estrogen-receptor status (ERα+/ERβ+).
MCF-10a cells were maintained in DMEM-F12 (Invitrogen) containing 10% horse serum (Invitrogen), 2 mM L-glutamine, 20 μg/mL gentamicin (Panpharma), 20 ng/mL epidermal growth factor (Sigma, St. Louis, MO), 100 ng/mL cholera toxin (Sigma), 0.25 UI/mL insulin (Novo Nordisk), and 0.5 μg/mL hydrocortisone (Sigma) held at 37°C with 5% CO2 in a humidified atmosphere. This cell line has a negative estrogen receptor status (ERα−/ERβ−).
MDA-MB-231 cells were grown in Leibovitz L-15 media with 15% fetal bovine serum (Invitrogen), 20 μg/mL gentamicin (Panpharma), and 2 mM L-glutamine in a 37°C humidified atmosphere without CO2. This cell line has ERα−/ERβ+estrogen receptor status.
Cell treatments
Treatments were carried out for 48 h with 18.5 μM genistein, 78.5 μM daidzein, or 2 μM 5-Azacytidine
DNA extraction
DNA was extracted using a Nonorganic DNA Extraction Kit (S4520, Millipore, Billerica, MA). After recovering the cells, 9 mL of wash buffer 1× was added to resuspend the pellet. After 15 min of incubation at room temperature, the cells were centrifuged at 1000×g for 20 min. The supernatant was discarded and the cells were resuspended in 3 mL of suspension buffer I 1×. Lysis Buffer I (800 μL) and 50 μL of protein digesting enzyme were added to the suspension. The samples were incubated for 2 h at 50°C. After adding 1 mL of protein precipitating agent, centrifugation at 1000×g for 15 min was carried out. The supernatant thus obtained was mixed with two volumes of absolute ethanol. The precipitated DNA was recovered using an inoculating needle, dried for 5 min at room temperature, and dipped in 5 mL 70% ethanol. The DNA was resuspended in 300 μL of Suspension Buffer II. After vortexing them during 5 min, the samples were left in incubation at 50°C overnight. The quantity of DNA collected as well as the quality of the extraction were then determined by spectrometry using a NanoDrop™ (ND-8000, 8-sample spectrophotometer, NanoDrop Technologies®, Wilmington, DE).
MeDIP
Methylated DNA was immunoprecipitated using the MagMeDIP kit from diagenode according to manufacturer's instructions. DNA shearing optimization was carried out with a Bioruptor (diagenode): 300 μL of a 0.1 μg/μL DNA solution in 1.5-mL tubes was sheared at “low” power using 15 s on and 15 s off cycles for a total time of 20 min. For each sample, 45 μL water, 24 μL MagBuffer A, 6 μL MagBuffer B, 1.5 μL methylated DNA (positive control), 1.5 μL unmethylated DNA (negative control) and 12 μL sheared DNA (at 0.1 μg/μL) were mixed. Seventy-five microliters of this mix were used for immunoprecipitation (IP) and 7.5 μL were kept as the “input sample.” Samples were incubated at 95°C for 3 min and quickly chilled on ice. For each IP, 11 μL magnetic bead solution was prepared. Beads were washed twice with a 1:5 dilution of MagBuffer A and resuspended in 22 μL of the same solution. In each IP tube, 0.15 μL of anti-5-methylcytidine monoclonal antibody, 0.60 μL MagBuffer A, 2.25 μL water, 2 μL MagBuffer C, and 20 μL of washed beads were added. Samples were incubated on a rotating wheel at 4°C for 16 h. Beads were washed three times with 100 μL MagWash Buffer-1 and three times with 100 μL MagWash Buffer-2. For each IP and input sample, 1 μL proteinase K was added to 100 μL DNA Isolation Buffer. The supernatant of IP samples was removed and 100 μL of the prepared solution was added, while 92.5 μL were added to input samples. All samples were incubated for 15 min at 55°C and 15 min at 100°C. After centrifugation, samples were stored at −20°C before polymerase chain reaction (PCR) analysis.
PCR
PCR directed at the BRCA1 exon 1 and the BRCA2 exon 2 was carried out in a final 15 μL reaction with 6 μL of input DNA at 10 ng/μL or 6 μL immunoprecipitated DNA, 0.06 U/μL AmpliTaq DNA polymerase (Applied Biosystems, Bedford, MA), 600 nM of each primer, 600 μM dNTP (Applied Biosystems), 1× PCR buffer I (Applied Biosystems). Initial denaturation at 94°C for 5 min was followed by 30 cycles of annealing at 54°C for 20 s and extension at 72°C for 30 s. After PCR reaction, products were resolved on 2% agarose gels, stained with Gel Red™ (Biotium, Hayward, CA). The primers used were designed as follows: primers targeting BRCA1 exon 1 on position chr17: 41277212-41277680 negative strand (GRCh37/hg19 Assembly); Forward: 5′-GTGATGCAATAAGCCGCAA-3′; Reverse: 5′-TCGGAAATCCACTCTCCCAC-3′ and for BRCA2 exon 2 on position chr13: 32890440-32890750 positive strand (GRCh37/hg19 Assembly): Forward: 5′-CCAGGAGATGGGACTGAATTAG-3′; Reverse: 5′-CTGTGACGTACTGGGTTTTTAGC-3′. The amplified regions are located near the promoter of the corresponding genes and consequently near the CpG islands.
Western blotting
Proteins were extracted from cells with a lysis buffer containing 20 mM Tris (pH 8), 50 mM EDTA, 0.8% NaCl, 0.1% Triton X-100, and 1% Glycerol. rotease inhibitors (1%) (Protease Inhibitor Cocktail; Sigma) and 1% phosphatase inhibitors (Phosphatase Inhibitor Cocktail 2; Sigma) were added to the basic buffer extemporaneously. Fifty microgram proteins were electrophoresed on a SDS-polyacrylamide gel and transferred onto a nitrocellulose membrane. After 1-h blocking in Tris-buffered saline Tween 0.1% (TBST) containing 5% milk, membranes were incubated overnight at 4°C with anti-BRCA1 [1:150 Mouse (Ab-1); Calbiochem, LaJolla, CA), anti-BRCA2 [1:50 Rabbit (H-300), Santa Cruz Biotechnology, Santa Cruz, CA], or anti-actin [1:120,000 Mouse (Ab-1); Calbiochem] antibodies. Membranes were then washed three times in TBST and incubated for 1 h with alkaline phosphatase-conjugated secondary antibody [1:2,000 goat antimouse IgG (H&L) AP conjugate; Promega 5372B, Madison, WI; or 1:2,000 goat antirabbit IgG (Fc) AP conjugate; Promega 5373B]. Detection was then performed with the Western Blue detection system (Promega). Relative quantification of immunoblotted proteins was achieved using the Quantity One software (Biorad, Hercules, CA) with local background subtraction method. A ratio between the intensity of the protein of interest and a reference protein (Actin) was then calculated. The relative ratio was then calculated between each condition and the reference condition (DMSO-treated cells).
Immunohistochemistry
Alcohol–formalin–acetic acid-fixed and paraffin-embedded sections (4 μm) of soy-treated or untreated MCF-7, MDA-MB-231, and MCF-10a cell pellets were cut using a microtome. They were mounted on silanized glass slides (Starfrost, Duiven, The Netherlands), and dried overnight at 37°C. Slides were processed on an automated Benchmark XT immunohistochemical instrument (Ventana, Oro Valley, AZ). In particular, sections were deparaffinised and rehydrated using EZ Prep (Ventana) and heat-induced antigen retrieval using CC1 (Ventana) was performed for 30 min. Slides were then incubated at 37°C for 44 min with primary MoAbs anti-BRCA1 (17F8, GeneTeX, San Antonio, TX) (1:10) or anti-BRCA2 (5F6, GeneTeX) (1:20). For detection, we used the UltraView universal DAB detection kit (Ventana). Signal was amplified using the Ventana amplification kit. Slides were then counterstained with hematoxylin for 3 min, rinsed in distilled water, and coverslipped with an aqueous Faramount mounting media (DAKO, Glostrup, Denmark). The primary polyclonal antibody was omitted and replaced with PBS as a negative control.
Immunofluorescence staining and confocal analysis of DNA methylation and BRCA1 or BRCA2 expression
Staining was realized on MCF-7, MDA-MB-231, and MCF-10a treated during 48 h with genistein (18.5 μM), daidzein (78.5 μM), 5-azacytidine (2 μM), budesonide (2 μM), or DMSO. To prepare nuclei, cells were scraped with a rubber policeman after addition of dropewise addition of prewarmed (37°C) 0.07 M KCl (3 mL), washed three times with phosphate-buffered saline (PBS). Nuclei were then pelleted by centrifugation at 800×g, for 20 min (21°C), resuspended, and fixed by dropewise addition of 10 mL of a solution of fixation [methanol/acetic acid mixture (3:1)] over a period of 20 min at 21°C. After centrifugation at 800×g, this step was repeated twice (Koch and Stratling, 2004). The nuclear suspension was resuspended by 5 mL of solution of fixation. A total of 450 μL of suspension was transferred into slides (4 Chambers Polystyrene Vessel, Becton Dickinson Culture Slides®) and irradiated with ultraviolet light for 3 h (30 cm from UV-C 30W lamp) (Miller et al., 1974). After 3 washing, to saturate nonspecific sites, nuclei were incubated overnight at 4°C with 400 μL of PBS containing 2% bovine serum albumin. Nuclei were incubated (300 μL) for 1 h at room temperature with primary antibodies (dilution of 1:50 in PBS), respectively, anti-5-methylcytidine [mouse, monoclonal antibody (MoAb), 33D3, Abcam, Cambridge, MA], anti-MeCP2 [goat, polyclonal antibody (PoAb), N-17, Santa Cruz Biotechnology], anti-BRCA1 (mouse, MoAb, MS 110; Abcam], or anti-BRCA2 (rabbit, PoAb, H-300, Santa Cruz Biotechnology]. Incubations with 300 μL of secondary antibodies (dilution of 1:200 in PBS), respectively, Alexa fluor 488 antimouse IgG (Invitrogen), Alexa fluor 488 antigoat IgG (Invitrogen) or Cy3 antirabbit IgG (Jackson Immunoresearch, West Grove, PA) secondary antibodies were carried out for 30 min in the dark (dilution of 1:200 in PBS). After three washing, slides were mounted with Hoechst (33258 pentahydrate H3569, Invitrogen), and examined using TCS SP5 (Leica, Buffalo Grove, IL) confocal microscope. For a quantitative analysis of intensity of fluorescence, images were subjected to mean gray value measures using the Image J software from the National Institutes of Health (http://rsb.info.nih.gov/ij/). For statistical analyses, a Student's t-test was performed using the data presented in the table, and p<0.05 was considered statistically significant compared with cells treated with the solvent, DMSO.
Results
Effects of soy phytoestrogens on BRCA1 and BRCA2 methylated DNA
BRCA1 primers targeting to exon1 and BRCA2 primers targeting to exon 2 were used for PCR after the MeDIP because they are near the promoter of the correspondent genes, and hence, near the CpG islands. BRCA1 was found methylated in DMSO control conditions as there was an amplification in input (total DNA) and immunoprecipitated DNA. BRCA1 was demethylated after genistein, daidzein, or 5-azacytidine treatment, corresponding here to a decrease in the signal by comparison with the input DNA in MDA-MB-231, MCF-7, and MCF-10a cell lines. Notably, our results showed that genistein is a better demethylant agent than daidzein in MDA-MB-231, MCF-7, and MCF-10a, and 5-azacytidine was found a complete demethylant (Fig. 1a).

MeDIP-PCR with BRCA1-exon1 (
The BRCA2 gene was found methylated in DMSO control conditions for the three breast cancer cell lines as they exhibited an amplification in the input and the immunoprecipitated DNA. BRCA2 demonstrated a demethylation after genistein, daidzein, or 5-azacytidine treatment by comparison of the IP band to the input. As expected, 5-azacytidine was strongly demethylant due to the disappearance of the IP band in the three cell lines. Genistein or daidzein treatment exhibited relatively weaker demethylant effect, as the IP band did not completely disappear in these conditions (Fig. 1b).
Quantification of BRCA1, BRCA2 protein expression after Western blotting in soy-treated breast cell lines
To demonstrate the impact of isoflavone treatments on expression, we quantified the BRCA1 and BRCA2 proteins after Western blotting with MCF-7 cells treated with these phytoestrogens (Fig. 2a). We found an increase in BRCA1 and BRCA2 protein expression following treatment with 18.5 μM genistein or 78.5 μM daidzein or 2 μM 5-azacytidine (demethylating agent) for 48 h, compared to the control corresponding to the DMSO solvent. In MDA-MB-231 (Fig. 2b) and MCF-10a (Fig. 2c) cells, the relative ratio calculated for BRCA1 and BRCA2 proteins were also increased following treatment with genistein or daidzein or 5-azacytidine.

BRCA1 and BRCA2 protein expression in MCF-7 (
BRCA1 and BRCA2 protein staining by immunohistochemistry
By immunohistochemistry, we demonstrated that the pattern of staining with the MoAbs against BRCA1 and BRCA2 was modified in MDA-MB-231, MCF-7, and MCF-10a breast cells after 5-azacytidine (a demethylant agent), genistein, or daidzein treatment by comparison to DMSO cells corresponding to soy untreated cells. The demethylant agent was used as control to demonstrate the demethylant action of genistein and daidzein. With all these chemical substances intensive nuclear, nucleolar and cytoplamic staining were observed (Fig. 3).

For BRCA1 and BRCA2 expression in MDA-MB-231, MCF-7, and MCF-10a breast cells, 17F8 anti-BRCA1 and 5F6 anti-BRCA2 MoAbs showed intensive nuclear and cytoplamic staining (arrowhead) after 2 μM 5-azacytidine (a demethylant agent), 18.5 μM genistein, or 78.5 μM daidzein treatment, compared to DMSO control corresponding to soy untreated cells. Some nucleoli in the nuclei were stained. N, nucleus; Cyt, cytoplasm; NU, nucleoli.
Genistein and daidzein decrease DNA methylation and increase BRCA1 and BRCA2 expression
We determined the effects of two isoflavones genistein (18.5 μM) and daidzein (78.5 μM) on BRCA1 and BRCA2 expression. After treatments for 48 h with genistein and daidzein, intensity of fluorescence of BRCA1 antibody increased significantly in MCF-7, MDA-MB-231, and MCF-10a cells (Fig. 4a); likewise, treatment with a demethylant agent, 5-azacytidine (2 μM), which is also significant. On the contrary, intensity of fluorescence decreased significantly when cells were treated with budesonide (2 μM). Similarly, intensity of fluorescence after incubation with BRCA2 antibody and treatments with 5-azacytidine (2 μM) and isoflavones increased significantly in the three cell lines, whereas treatment with budesonide (2 μM) showed a significant decrease of intensity of fluorescence (Fig. 4b).
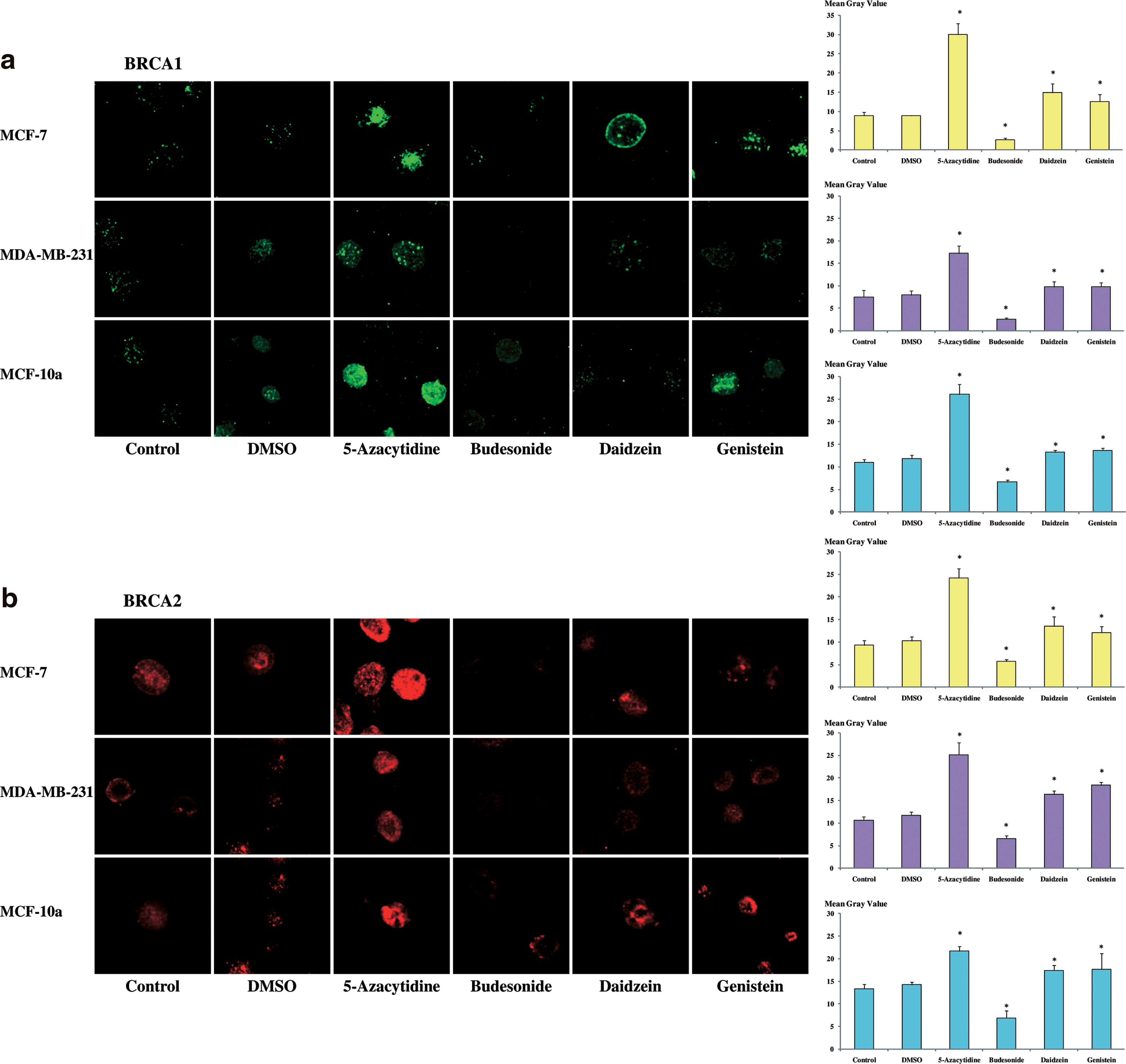
Effects of daidzein and genistein on BRCA1 and BRCA2 expression in MCF-7, MDA-MB-231, and MCF-10a cell lines. Cells were treated during 48 h with daidzein (78.5 μM), genistein (18.5 μM), 5-azacytidine (2 μM), or budesonide (2 μM). Cells were also treated with DMSO, solvent in which substances were dilued. Then, cells were immunostained (
An anti-5-methylcytidine antibody was used to study DNA methylation (Fig. 5a). Further to MCF-7, MDA-MB-231, and MCF-10a treatments during 48 h with daidzein (78.5 μM) or genistein (18.5 μM) or 5-azacytidine (2 μM), intensity of fluorescence with anti-5-methylcytidine antibody decreased significantly. With methylant agent (budesonide 2 μM), intensity of fluorescence was significantly increased.
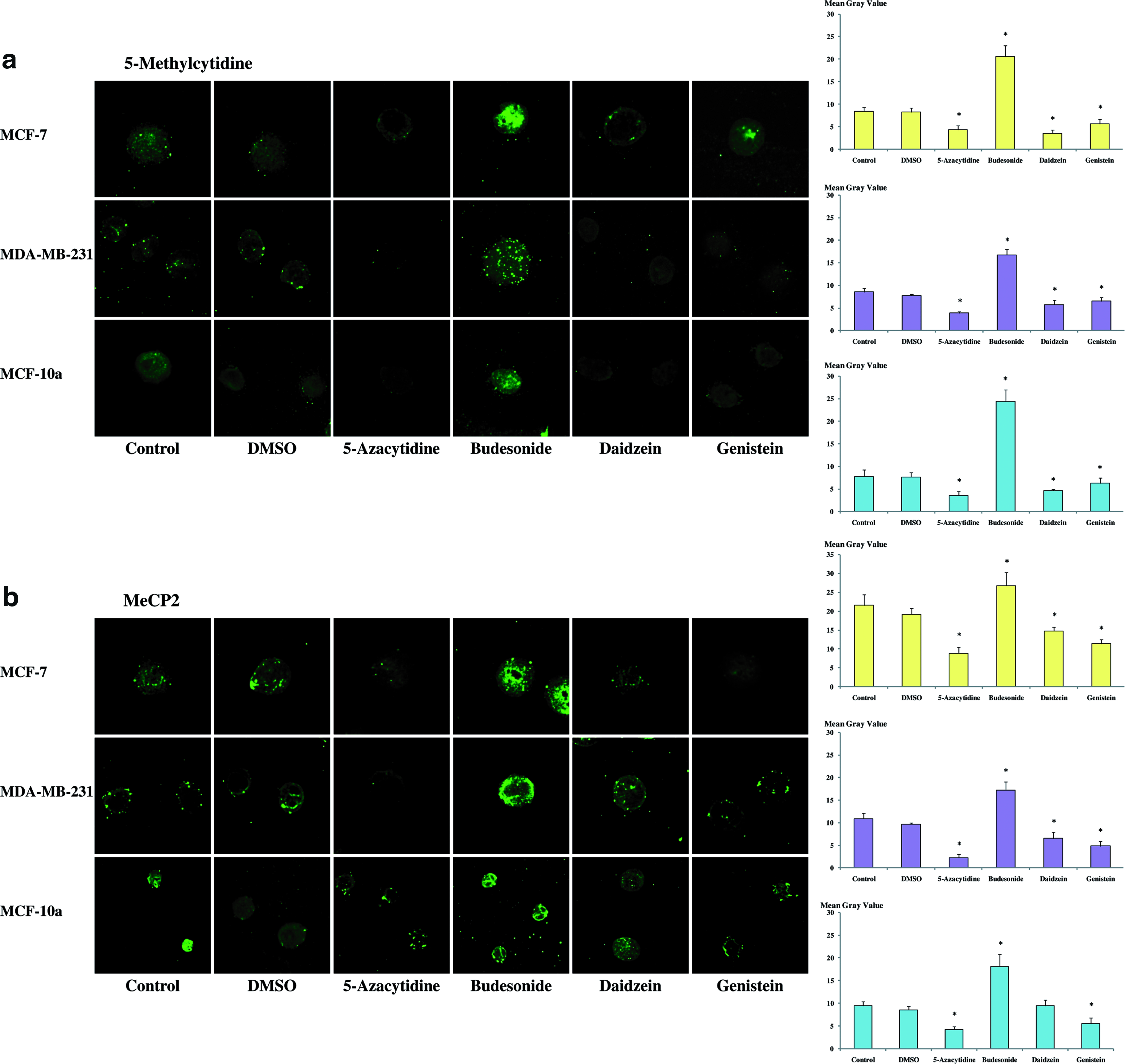
Effects of daidzein and genistein on DNA methylation in MCF-7, MDA-MB-231, and MCF-10a cell lines. Cells were treated during 48 h with daidzein (78.5 μM), genistein (18.5 μM), 5-azacytidine (2 μM), or budesonide (2 μM). Cells were also treated with DMSO solvent in which substances were dilued. Then, cells were immunostained (
MeCP2 protein variation was also investigated for its different deregulation in breast cancer, compared to BRCA1 and BRCA2. The protein decreased following phytoestrogens and 5-azacytidine exposure and increased after budesonide treatment (Fig. 5b). No difference between control and DMSO were observed.
Discussion
Although soy phytoestrogens have been postulated to exert a protective effect against breast cancer, the attendant mechanisms, in particular, epigenetics underpinnings, have remained elusive. We investigated the putative effects on DNA methylation by two naturally occurring isoflavones, genistein and daidzein, in the present study of the BRCA1 and BRCA2 oncosuppressor genes in breast cancer cell lines. In the present study, we suggest that genistein and daidzein might reverse DNA hypermethylation and increase the expression of the methylation-impacted genes BRCA1 and BRCA2. These observations, although requiring replication, provide new evidence on potential epigenetic mechanisms by which genistein and daidzein might contribute to regulation of the BRCA1 and BRCA2.
Our MeDIP-PCR results show that genistein and daidzein appear to demethylate BRCA1 and BRCA2. To the best of our knowledge, this has not been reported in the literature previously. Although this demethylation effect is less intense than with 5-azacytidine, studies with longer exposures to these phytoestrogens should be undertaken for effects on efficiency of the demethylation. The use of techniques such as bisulfite sequencing would allow more precise localization and quantification of the methylation, thus bringing further clarity over the mechanism of action of genistein and daidzein in regard to their putative chemoprevention activity against breast cancer.
We performed Western blot analysis and quantification of BRCA1 and BRCA2 proteins following soy phytoestrogens or 5-azacytidine treatment in the three cell lines. All treatments resulted in a change in the direction of an increase in protein, although the observed effects were slight or moderate in some cases of the two studied proteins. Clearer results, however, were found using immunohistochemistry. Indeed, intensive nuclear, nucleolar, and cytoplasmic staining, corresponding to the BRCA1 and BRCA2 proteins, were observed after soy phytoestrogens or 5-azacytidine treatment. Previous studies had shown that the methylation of BRCA1 was negatively correlated to mRNA expression (Catteau et al., 1999; Lee et al., 2007; Rice et al., 2000), so association of a demethylation of BRCA1 and an increase in subsequent protein is consistent with this data. For BRCA2, no data was found for association between methylation of the gene and its expression in the literature, but our results suggest it behaves similarly to BRCA1 in this respect. Quantification of the mRNA of BRCA1 and BRCA2 in our three cell lines following phytoestrogen or 5-azacytidine treatments would be of interest in future studies in order to confirm that the observed demethylation of the genes causes an increase in their mRNA, which would be responsible for an increase in protein synthesis.
Other studies that have been undertaken with other genes and cell lines are of noteworthy in the context of the present report. In 2005, Fang et al. reported, in human esophageal squamous carcinogenesis, that genistein inhibited DNA methyltransferases, thus reversing the methylation and permitting the reexpression of RARβ, p16, and MGMT (O6-methylguanine methyltransferase) genes. More recently, King-Batoon et al. (2008) reported that both genistein and lycopene, at very low concentrations that are relevant for the human diet, could mitigate tumorigenic processes via promoter demethylation and increase of gene expression.
Immunofluorescence staining and confocal analysis of BRCA1 and BRCA2 proteins showed the increase of protein expression following phytoestrogens or 5-azacytidine treatments in our study. The quantification allowed us to show the significance of this effect. Related to the analysis of 5-methylcitidine presence in the nucleus, which is reduced following phytoestrogens and 5-azacytidine treatment, it seems that those treatments demethtylate globally DNA, including the BRCA1 and BRCA2 genes, thus allowing an increase in their expression. The mechanism of action of 5-azacytidine implies the demethylation of all DNA. For genistein and daidzein, the observed demethylation could be the result of the demethylation of a certain number of genes only. This hypothesis could be tested with a precise analysis of the methylation of many locus in DNA, for example, by meDIP-on Chip approaches.
Finally, it is noteworthy that we investigated variations of MeCP2, a methyl binding domain protein, whose deregulation is different from BRCA1 or BRCA2 expression in breast cancer, to see how its expression would behave following soy phytoestrogen exposures. Indeed, Billard et al. (2002) has suggested an increase in the MeCP2 mRNA in low-differentiated neoplasia, whereas Müller et al. (2003) clearly showed an increase of MeCP2 mRNA in neoplastic breast tissue. MeCP2 protein expression was decreased following soy phytoestrogens or 5-azacytidine treatments. As 5-azacytidine is a demethylating agent, we suggest MeCP2 expression might be linked to the global amount of methylation in the genome. This hypothesis has to be tested in future studies; our present findings do not allow us to conclude whether soy phytoestrogens change methylation of select candidate genes, or have a more global and genome-wide effect on DNA methylation.
Müller et al. (2003) has pointed out that MeCP2 expression was more important in estrogen receptor positive cells but, despite the fact we used cell lines with different estrogen receptor status, we did not observe any difference.
We wish to underscore that DNA methylation is only a part of broader range of possible epigenetic mechanisms, including histone modifications (Esteller, 2007), which should also be considered in breast cancer chemoprevention studies to obtain a more comprehensive view of the effects of soy phytoestrogens. Studies of gene silencing associated markers, such as H3-K27-me or H3-K9-me or gene-activating markers as H3-K4-me or H3-K9-ac can be pursued as well (Elsheikh et al., 2009).
Interestingly, Li et al. (2009) has reported that genistein could deplete telomerase activity in both precancerous and cancerous cells. They showed that direct regulation of key transcription factors and epigenetic modulation were caused by genistein. This result is challenging, as those two mechanisms might be linked, thus complicating the understanding of phytoestrogens' mode of action.
Conclusions
Our findings in the present study lend evidence for a demethylating effect of genistein and daidzein on BRCA1 and BRCA2 oncosuppressor genes, which is associated with an increase in the expression of the encoded proteins. We note that this effect might not be specific to these genes and could be more global in nature, at the scale of the genome, an area that is in need of future investigations. Additionally, the localization and quantification of the methylation of BRCA1 and BRCA2 using bisulfite sequencing, extension of the study to a broader set of genes, or use a more global approach such as meDIP-on-Chip are recommended in regard to future line of research to achieve a deeper understanding of phytoestrogen mechanism of action in breast cancer. Finally, as epigenetic mechanisms are often intertwined and may act in concert, we suggest that the study of histone modifications by genistein and daidzein are warranted. Ultimately, this line of research is essential before future clinical chemoprevention studies of phytoestrogens can be justified in humans and grounded on a firm molecular understanding of their putative mechanism of action against breast cancer pathogenesis.
Footnotes
Acknowledgments
We gratefully acknowledge the engaged help of Caroline Vachias and Jean-Louis Couderc from the ICCF (Imagerie confocale de Clermont-Ferrand) platform for confocal microscopy use. We thank the technical assistance of Christelle Picard, Nicolas Sonnier, Elodie Heinry, Sinda Mabouli, Julie Durif, and Marine Gueugneau. We also thank “La ligue contre le Cancer, comités de la région Auvergne.” Rémy Bosviel received a grant from the Auvergne Regional Council/CPER 2008+FEDER no. 32316–0930FDBG–106NL.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.