
Abstract
Abstract
Comparative transcriptome study of the leaf and root tissues of Catharanthus roseus is a prerequisite for causing any favorable tissue-specific change in the secondary metabolism of this species. This study was aimed at comparative analysis of the leaf and root cDNAs in C. roseus, using a cDNA-AFLP approach. Using 64 primer combinations (EcoRI and MseI), a total of 784 distinct transcriptionally-defined fragments (TDFs) could be detected in the root and leaf tissue transcript populations. The leaf tissue yielded a larger number of TDFs than the root tissue (556 versus 464), indicating a greater variety of expressing genes in the leaf. The leaf-specific TDFs (320) outnumbered the root-specific TDFs (228), indicating a higher number of leaf-specific functions and the relative complexity of the leaf tissue vis-à-vis the root tissue. Among the 236 TDFs that were detected in both types of tissues, 42 had nearly equal expression levels in both the tissues (L=R). Common TDFs having higher expression levels in the leaf (L>R; 124) outnumbered those having higher expression levels in the root (L<R; 70), which again reinforced the predominance of the leaf transcriptome over the root transcriptome.
Introduction
The existence of different alkaloids and expressed genes within the aerial and underground tissues of C. roseus indicates the expression of distinct tissue-specific pathways. In turn, these pathways are responsible for the precise tissue-specific compartmentation of TIA biosynthesis (Laflamme et al., 2001). Additionally, there are several distinct functions associated with either of the tissue types, all of which result from the differential expression of structural and regulatory factors in these two tissues. Gene prospecting through comparative transcriptome analysis of the aerial and underground tissues may provide a better understanding of the tissue-specific differences. Earlier, an effort was made in this direction by our group using an expressed sequence tag (EST)-based approach to differentiate the root and leaf transcriptomes in C. roseus (Shukla et al., 2006). In the present study we describe a cDNA-AFLP approach to differentiate the root and leaf organ cDNAs in C. roseus only on a numerical basis.
Materials and Methods
RNA isolation
For cDNA synthesis, poly (A)+ mRNA was isolated from the leaf (middle level) and root tissues of 6-month-old mature field-grown cv. ‘Dhawal’ [a leaf alkaloid-rich genotype of C. roseus (Kulkarni et al., 2003), for which detailed chronomics of TIA biosynthesis has been worked out previously (Shukla et al., 2006)] plants (vegetatively maintained and genotypically similar; available from the National Gene Bank for Medicinal and Aromatic Plants at CIMAP, Lucknow). Sampling was carried out in the month of June (average daytime temperature ≥42°C), under conditions favoring high in planta bisindole alkaloid biosynthesis (Shukla, 2005), using a previously described protocol (Shukla et al., 2005).
cDNA-AFLP
In the present study, the modified protocol for cDNA-amplified fragment length polymorphism (cDNA-AFLP) described by Bachem and associates (1998) was implemented with slight modifications as described below. Double-stranded cDNA was synthesized from both (leaf and root) poly (A)+ mRNA samples using the Universal Riboclone® cDNA Synthesis System (Promega, Fitchburg, WI, USA). Exactly equal amounts (2 μg) of poly (A)+ mRNA (spectrophotometrically and electrophoretically determined) was taken for both tissues, and 1 μg oligo (dT)15 primer was used to prime the first-strand cDNA synthesis. All the steps were performed per the manufacturer's guidelines, and the double-stranded cDNA ends were blunted with T4 DNA polymerase.
For the AFLP reactions, the AFLP™ Plant Mapping Kit (PE Applied Biosystems, Foster City, CA, USA) was used. The restriction of each of the cDNA samples with two restriction endonucleases (EcoRI and Tru9I, which is an isoschizomer of MseI), and ligation of EcoRI and MseI adaptors to the ends of restricted cDNA, was carried out in a single reaction using the manufacturer's guidelines. The sequence of the adaptor oligonucleotide was such that its ligation to the restricted cDNA did not regenerate the enzyme recognition site, and thus restriction did not recur after ligation. The sequences of the adaptors and the restriction site served as primer binding sites for a subsequent low-level selection or preselective amplification of the restricted transcriptionally-defined fragments (TDFs). The MseI complementary primer contained a 3′ C, and the EcoRI complementary primer contained a 3′ A. For the preselective amplification the thermal cycler was programmed as 72°C for 2 min; 20 cycles of 94°C for 20 sec, 56°C for 30 sec, 72°C for 2 min; 60°C for 30 min; and 4°C for infinite period. The amplification product obtained was found to be in the range of 50–700 bp, and was diluted to a final concentration of 1 ng/μL to serve as template for the selective amplification reactions using all the 64 available enzyme primer-pair combinations (eight MseI primers and eight EcoRI primers in the AFLP selective amplification start-up module of the kit). The eight MseI primers were complementary to the MseI adaptor sequence, had three additional bases at the 3′ end (3′ CXX), and an unlabeled 5′ end, whereas the eight EcoRI primers were complementary to the EcoRI adaptor sequence, had three additional bases at the 3′ end (3′ AXX), and a fluorescent dye label (FAM, JOE, or NED, giving them blue, green, or yellow colors, respectively) at the 5′ end. The selective amplification reactions were multiplexed and performed in a thermal cycler programmed as 94°C for 2 min; 10 cycles of 94°C for 20 sec, 66°C (−1°C/cycle) for 30 sec, 72°C for 2 min; 20 cycles of 94°C for 20 sec, 56°C for 30 sec, 72°C for 2 min; 60°C for 30 min; 4°C for infinite period. Selective amplification with an EcoRI and an MseI primer amplified primarily EcoRI-MseI-ended TDFs. The resulting products were size fractionated on a 5%, 29:1 polyacrylamide sequencing-type gel.
Data analysis
The data were analyzed by GeneScan Analysis® v 3.1 software (PE Applied Biosystems), and the size calling of the peaks representing the TDFs was done by identifying the peaks of the in-lane size standard (Gene Scan™-500 ROX™) found in each sample. Only peaks above 35 bp in size were considered for analysis. The peak height or fluorescent signal strength of a dye-labeled TDF was indicative of the quantitative abundance of the TDF, and was taken as a measure of its expression level.
Results and Discussion
In order to get a numerical estimate of the differences and similarities between the root and leaf tissue cDNAs of C. roseus, a cDNA-AFLP approach was used in the present study and the results obtained are described below.
TDF analysis of the selective amplification products (of the 64 multiplexed PCR reactions) revealed a comprehensive comparative picture of the root (R) and leaf (L) organ cDNAs (Table 1). Figure 1 (Venn diagram) illustrates the organ-specific distribution of the obtained TDFs. In this analysis, a total of 784 distinct TDFs could be detected in the root and leaf organ cDNAs. The analyzed leaf (L) subset yielded a larger number of TDFs than the root (R) subset (556 compared to 464), indicating a greater variety of expressing genes in the leaf (L). TDFs that were expressed either only in the root (228) or only in the leaf (320) may be considered organ-specific, and hence may be implicated in development and/or function of these distinct organs. Of the 236 TDFs that were detected in both of the organ types, 42 had nearly equal expression levels in both organs (L being equal to R with L/R or R/L expression ratio≤1.25). Common TDFs having higher expression levels in the leaf (L>R; 124) outnumbered those having higher expression levels in the root (L<R; 70). Size distribution of the detected TDFs was also determined (Table 2). Most of the detected TDFs were less than 150 bp in size (487/784; 62.12%). Approximately 25.76% (202/784) of the TDFs were in the range 150–300 bp, whereas only 12.12% (95/784) of the TDFs were more than 300 bp in size.
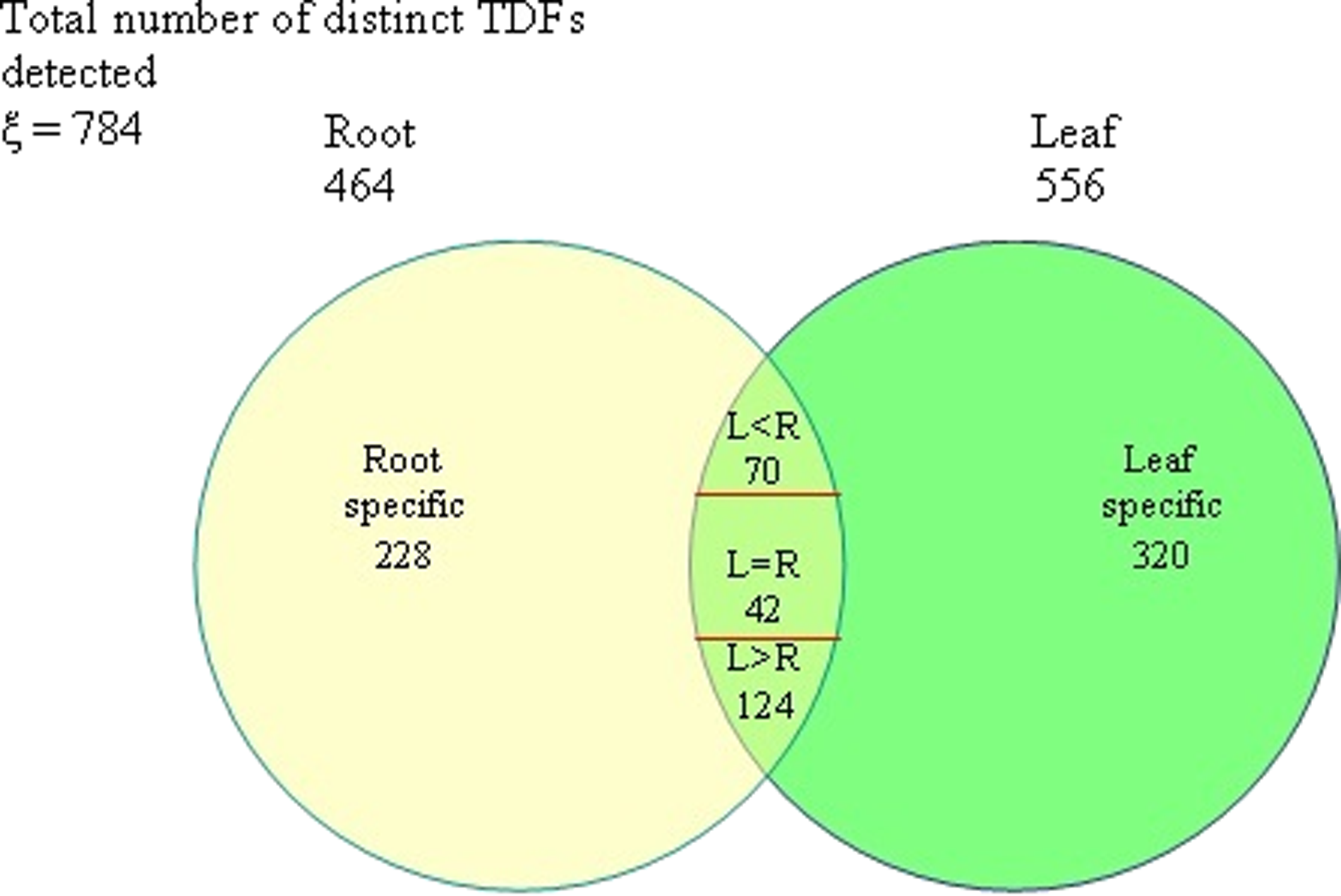
Comparative distribution of transcriptionally-defined fragments (TDFs) in the root and leaf transcriptomes of Catharanthus roseus.
L=R means L/R or R/L expression ratio≤1.25.
TDF, transcriptionally-defined fragment; cDNA-AFLP, cDNA-amplified fragment length polymorphism.
L=R means L/R or R/L expression ratio≤1.25.
TDF, transcriptionally-defined fragment; cDNA-AFLP, cDNA-amplified fragment length polymorphism.
Leaf-specific TDFs (320) outnumbered root-specific TDFs (228), indicating higher numbers of leaf-specific functions and the relative complexity of the leaf transcriptome vis-à-vis the root transcriptome. This is in accord with earlier reports in literature that have described the plant root as a simple organ that is comprised of a relatively few differentiated cell types (Schiefelbein and Benfey, 1991). The root apical meristem is easily accessible (unlike the shoot apical meristem), and lacks pigmentation and branching primordial. Root morphogenesis normally occurs in a reiterative and uniform fashion, without any major change in the organization of the root apex. Thus, all the stages of root development are apparent at all times, and there is nothing analogous to the vegetative-to-floral developmental transition that occurs in shoot meristems.
Even though the root possesses a smaller number of specifically-expressing transcripts, attempts have been made earlier in other plants to find genes that express specifically in the root (Choi et al., 1996; Conkling et al., 1990), and those that express preferentially in the root (Evans et al., 1988; Lin et al., 2005). In a previous microarray-based study of 1400 Arabidopsis cDNAs to compare expression patterns between roots and leaves, it was found that 30% of the genes had higher expression levels in roots compared to leaves, but few differences were detected between open flowers and flower buds or leaves (Ruan et al., 1998). It has also been demonstrated that a considerable number of the leaf mRNAs in Arabidopsis are differentially expressed in a day- or night-dependent manner (Schaffer et al., 2000). These observations indicate that the root and the leaf organs are transcriptomically different, and are consequently functionally distinct, with higher numbers of distinct functions associated with the leaf, possibly due to its exposure to aerial stress factors. The leaf has to provide the first line of defense against herbivory and pests, which makes a sophisticated arsenal of secondary metabolites mandatory. Functional regulation of this arsenal, as well as the extra leaf-specific primary metabolic functions (like photosynthesis and transpiration), are primarily responsible for the complexity of the leaf transcriptome compared to the root transcriptome.
It is evident that the numerical differences obtained in the leaf and root organ transcriptomes of C. roseus could not be directly ascribed only to the TIA diversity present in the two organs. However, the leaf-specific TDF subset could provide important clues related to the biosynthesis and accumulation of key leaf-specific TIAs upon downstream analysis. In this context it is relevant to note that despite functional divergence of the two organ-specific transcriptomes, there might exist homology at the genetic level. An example of this could be observed in the C. roseus enzymes—root-specific minovincinine-19-O-acetyltransferase (MAT) and leaf-specific deacetylvindoline-4-O-acetyltransferase (DAT), which have 63% nucleic acid identity and 78% deduced amino acid sequence identity, but different substrate specificities (Laflamme et al., 2001).
Although redundancy was not determined in the present cDNA-AFLP-based study, the number of leaf-specific TDFs (320) obtained was higher than the number of leaf-specific ESTs (55) obtained through a subtractive hybridization approach, in which leaf and root cDNA populations were used as “tester” and “driver,” respectively (Shukla et al., 2006). This demonstrates the higher efficiency of the cDNA-AFLP approach to identify differential transcripts. Also, next-generation sequencing methods may be adopted for comparative root and leaf deep transcriptome sequencing analysis, as was shown recently for Avena barbata (Swarbreck et al., 2011). In conclusion, the cDNA-AFLP analysis of root and leaf organs of C. roseus provides a direct and easy tool for discovering novel transcripts and genes expressing in an organ-specific manner. This study was carried out only to obtain a numerical estimate of the differences between the transcripts expressing in the leaf and root of C. roseus. To fully exploit the benefits of the open system cDNA-AFLP approach used in the present study vis-à-vis bisindole alkaloid biosynthesis, further characterization of the leaf-specific TDFs is required.
Footnotes
Acknowledgments
We are grateful to the Council of Scientific and Industrial Research, India, and the Department of Biotechnology, India, for financial support.
Author Disclosure Statement
No competing financial interests exist.