
Abstract
Abstract
Due to high prevalence and slow progression of prostate cancer, primary prevention appears to be attractive strategy for its eradication. During the last decade, curcumin (diferuloylmethane), a natural compound from the root of turmeric (Curcuma longa), was described as a potent chemopreventive agent. Curcumin exhibits anti-inflammatory, anticarcinogenic, antiproliferative, antiangiogenic, and antioxidant properties in various cancer cell models. This study was designed to identify proteins involved in the anticancer activity of curcumin in androgen-dependent (22Rv1) and -independent (PC-3) human prostate cancer cell lines using two-dimensional difference in gel electrophoresis (2D-DIGE). Out of 425 differentially expressed spots, we describe here the MALDI-TOF-MS analysis of 192 spots of interest, selected by their expression profile. This approach allowed the identification of 60 differentially expressed proteins (32 in 22Rv1 cells and 47 in PC-3 cells). Nineteen proteins are regulated in both cell lines. Further bioinformatic analysis shows that proteins modulated by curcumin are implicated in protein folding (such as heat-shock protein PPP2R1A; RNA splicing proteins RBM17, DDX39; cell death proteins HMGB1 and NPM1; proteins involved in androgen receptor signaling, NPM1 and FKBP4/FKBP52), and that this compound could have an impact on miR-141, miR-152, and miR-183 expression. Taken together, these data support the hypothesis that curcumin is an interesting chemopreventive agent as it modulates the expression of proteins that potentially contribute to prostate carcinogenesis.
Introduction
Curcumin (diferuloylmethane) is a polyphenol extracted from the rhizome of the plant Curcuma longa. It is a natural compound that has been used for centuries in Ayurvedic, Chinese, and Hindu traditional medicines (Fig. 1). Today, it becomes a promising chemopreventive compound able to reverse, inhibit, or prevent the development of cancer by inhibiting specific molecular signaling pathways involved in carcinogenesis (Aggarwal et al., 2007; Duvoix et al., 2005; Hatcher et al., 2008; Teiten et al., 2010a). Recent studies showed that curcumin possesses antioxidant, anti-inflammatory, antiproliferative, and antiangiogenic properties in several types of cancer (Kelkel et al., 2010; Menon and Sudheer, 2007; Reuter et al., 2008b; Teiten et al., 2009a). It inhibits multiple molecular mechanisms including glutathione conjugation (Blasius et al., 2004; Duvoix et al., 2003a) and metabolism (Reuter et al., 2009), heat-shock protein expression (Teiten et al., 2009b), JAK–STAT signaling pathways (Blasius et al., 2006), and cell death (Duvoix et al., 2003a, 2003b; Reuter et al., 2008b).
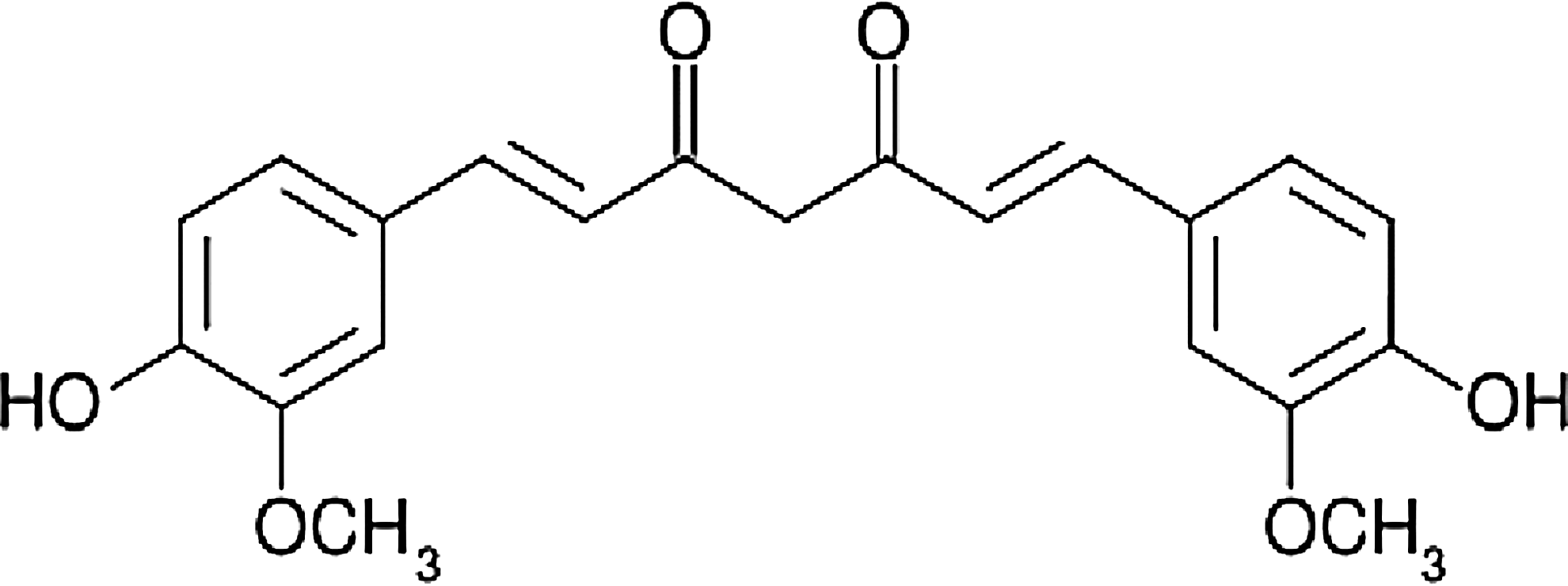
Chemical structure of curcumin (1,7-bis-4(hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione).
The emergence of proteomics allows us to design the present study to gain new insight in the effect of curcumin on prostate cancer cells. Using a two-dimensional difference in gel electrophoresis (2D-DIGE)-based proteomic analysis, we identified proteins involved in the anticancer activity of curcumin in androgen-dependent and -independent human prostate cancer cells. Previous studies that evaluated the impact of curcumin on a large scale of proteins were performed in breast and cervical cancer. Curcumin was reported to modulate the expression of proteins that contribute to DNA transcription; mRNA splicing and translation; amino acid synthesis; protein synthesis, folding and degradation; lipid metabolism; glycolysis and motility in breast cancer (Fang et al., 2011). In the case of cervical cancer, curcumin was reported to affect mainly cyclooxygenase-2, which is considered as a potential target for prevention and treatment of cervical cancer. Curcumin was also reported to affect proteins associated with cancer and cell cycle in cervical cancer cells (Madden et al., 2009). Our approach allowed us to investigate 425 differentially expressed spots after curcumin treatment. A total of 192 spots from this list were selected for MALDI-TOF-MS identification, and were shown to correspond to 60 proteins shown to be implicated in protein folding, RNA splicing, cell death, and androgen receptor signaling.
Taken together, our data support the hypothesis that curcumin is a potential chemopreventive agent by showing that curcumin modulates the expression of proteins already known to play an important role in prostate carcinogenesis.
Material and Methods
Cell culture and treatment
Three human prostate cancer cell lines, androgen-dependent (AD) LNCaP and 22Rv1 cells, and androgen-independent (AI) PC-3 cells, were cultured in RPMI 1640 medium (Lonza, Verviers, Belgium) supplemented with 10% fetal calf serum (FCS) (Lonza, Verviers, Belgium) and 1% (v/v) antibiotic–antimycotic (Lonza) at 37°C in a 5% CO2 humidified atmosphere. A stock solution was made by dissolving curcumin (Sigma, Bornem, Belgium) in 100% DMSO (Sigma-Aldrich, Bornem, Belgium) to a concentration of 20 mM. Subsequent dilutions were made in cell culture medium. In our experimental conditions, PC-3, LNCaP, and 22Rv1 cells were treated for 24 h with 20 μM of curcumin, or equivalent volume of DMSO. Cells were detached by scrapping in 1% phosphate-buffered saline (PBS) that had been washed and collected with supernatant.
Proteomic assays and statistical analysis
Proteins were extracted with a nuclear extraction kit (Active Motiv) according to the manufacturer's protocol to separate nuclear and cytosolic fractions. In brief, proteins were precipitated with acetone (Sigma-Aldrich), washed and resolubilized in DIGE/lysis buffer (7 M urea, 2 M thiourea, 4% CHAPS and 30 mM Tris) supplemented with nuclease mix (GE Healthcare) and protease inhibitors (Roche, Prophac, Luxembourg). Protein concentrations were determined using the 2D Quant kit (GE Healthcare).
For analytical gels, 50 μg of proteins were labeled with cyanine (Cy) 3 or Cy5 according to the manufacturer's instructions (GE Healthcare). A reference solution that was composed of equal amounts of each of the protein extracts and labeled with Cy2 was used as an internal control for normalizing. For preparative gels (picking and analytical gel), a mix of proteins labeled with each of the three cyanines (150 μg) was added with 200 μg of an equal amount of all proteins from the concerned cell line (22Rv1 or PC-3).
Isoelectric focusing was performed on a 24 cm pH 4–7 immobilized pH gradient (IPG) strip (GE Healthcare). It was rehydrated overnight with either the analytical batch of proteins, composed of 150 μg mix of proteins labeled with Cy3, Cy5, and Cy2, or 450 μg of the preparative batch, diluted in the rehydration buffer. A final volume of 450 μL was reached by adding 4.5 μL IPG buffer (GE Healthcare). The isoelectric focusing procedure was performed using an IPGphor II (GE Healthcare) with the following conditions: 40 V for 2 h, 70 V for 2 h, ramping to 300 V for 3 h, ramping to 1,000 V for 4 h, ramping to 8,000 V for 3 h, and 8,000 V for 6 h. Afterward, strips were equilibrated for 15 min in 10 mL of equilibration buffer (6 M urea, 30% glycerol, 2% SDS, 50 mM TRIS, and pH 8.8) added for reduction with 0.5% (w/v) dithiothreitol (DTT). Subsequently, solutions were alkylated by incubation for 15 min in 10 mL of equilibration buffer containing 2.5% (w/v) iodoacetamide. Second dimension separation was performed in 1-mm-thick, 12% SDS polyacrylamide gels (with bind silane-treated glass plate for preparative gels), proteins were run for 1 h at 1 W per gel and then for 16 h at 100 W total on an Ettan DALTtwelve System (GE Healthcare).
After electrophoresis, fluorescence-labeled proteins were visualized in the gels at 100 μm resolution using a Typhoon 9400 fluorescence scanner (GE Healthcare) with the appropriate excitation wavelength and emission filters.
Gel images were analyzed using Delta2D v.4 Software (Decodon). Raw data can be downloaded as supplementary material (Supplementary Files S1). Data extraction and statistical analyses were performed with the same software on a total of 12 gels. Spots were considered to be differentially expressed when Student's t-test p-values were ≤0.05 and when a spot was present on at least 8 out of 12 analyzed gels. A total of 192 spots were selected for picking and identification as they appeared differentially expressed in only one of the cell line tested. Some spots were added to the list, as they correspond to proteins differentially expressed in both cell lines.
Spot picking and mass spectrometry identification
Gel images were matched before manual spot picking with a 2-mm wide punch on a UV transilluminator. Gel plugs were stored at −20°C in a multiwell plate containing 50 μL of MilliQ water per well. Trypsin digestion was performed on an Ettan Spot Handling Workstation (GE Healthcare), after three steps of washing and desalting (two times in 50 mM ammonium bicarbonate in 50% v/v methanol, followed by 75% v/v CAN). Proteins were then digested with Trypsin Gold (MS grade, Promega, Madison, WI) at 8 mg/mL in 20 mM ammonium bicarbonate at 37°C for 6 h. After digestion, the resulting peptides were resolubilized in 2 μL of 50% (v/v) acetonitrile (ACN) containing 0.1% (v/v) trifluoroacetic acid (TFA) (Bohler et al., 2007). For protein identification, 0.7 or 1 μL of this solution was spotted on the mass spectrometer target and mixed with 0.7 μL of matrix, which was 7 μmg/mL α-cyano-4-hydroxycinnamic acid in a solution of 50% (v/v) ACN and 0.1% (v/v) TFA.
Peptide mass fingerprint and fragmentation spectra were acquired using a 4800 Proteomic Analyzer (Applied Biosystems, Foster City, CA), and the resulting spectra were subjected to a database search through the MASCOT interface (MASCOT 2.2, Matrix Science, London, UK) integrated in the GPS Explorer software suite (Applied Biosystems).
Searches in the NCBI (downloaded on September 24, 2010, with 11,888,344 sequences) and Swissprot (September 24, 2010, with 519,348 sequences) databases were performed using the following parameters: (1) error tolerance on peptide mass fingerprints of 100 ppm or 0.3 Da [tandem mass spectrometry (MS/MS)]; (2) fixed modifications: carbamidomethylation (C) and oxidation (M); (3) potential modifications: kynurenine (W) and double oxidation (W); and (4) a restriction to Homo sapiens in taxonomy. Search results were evaluated based on peptide scores, and identifications were manually validated.
Bioinformatics analyses
Protein lists were categorized into biologically meaningful categories using the bioinformatic enrichment tool DAVID (Dennis et al., 2003; Huang da et al., 2009), which outputs an EASE score. This score is a function of a penalized Fisher's exact test, which is more conservative than Fisher's exact test; therefore, fewer categories are considered significant. This procedure is performed to avoid false positives. Results were visualized using Enrichment Map plug-in version 1.1 (Merico et al., 2010), which is included in Cytoscape software version 2.8 (Shannon et al., 2003). Significant gene ontology (GO) categories associated with our protein lists were selected using the subset GO FAT Biological Processes. To highlight the most specific terms, we used this subset, which was released in version 6.7 and does not contain higher levels GO categories.
We used Pathway Studio (Ariadne Genomics Inc., Rockville, MD, version 8.0.132.0871) subnetwork enrichment tool to find significant expression regulators of over- and underexpressed proteins. Subnetworks were considered to be significant if they meet the following conditions: (1) p-value lower than 0.01, (2) effect on at least 2 proteins from our list, and (3) effect on selected protein expression, which includes promoter-binding and miRNA effects.
Pathway Studio was further used to contextualize our results by highlighting interactions between our proteins and curcumin that were previously established in the literature. Literature-reported relations were retrieved using Medscan software (Ariadne Genomics Inc., version 4.0.65.2626).
Western blot
After treatment, 107 cells were harvested and total proteins were extracted as previously described (Schreiber et al., 1989). We used a Bradford assay (Bio-Rad protein assay, Biorad, Nazareth, Belgium) to determine protein quantity. These proteins were then subjected to SDS-PAGE and were transferred onto a Hybond™-P membrane (GE Healthcare). Membranes were prehybridized overnight at 4°C in phosphate buffered saline at 1×containing 0.1% (v/v) Tween-20 (PBS-T) and 5% milk. Membranes were incubated with primary antibodies against the androgen receptor (Cell Signaling Technologies, Bioké, Leiden, The Netherlands), HMGB1 (Abcam, Cambridge, England) or beta-actin (Sigma-Aldrich), which was used as a loading control. Incubation was done in PBS-T containing 5% milk for 1 h at room temperature or overnight at 4°C. After washing, membranes were probed with horseradish peroxidase-labeled secondary antibodies for 1 h at room temperature. Proteins of interest were visualized with Ecl Plus Western blotting Detection System reagent with an Image Quant LAS 4000 (GE Healthcare).
Transient transfection
Transient transfections of LNCaP and PC-3 cells were performed by electroporation using the BioRad Gene Pulser (Biorad). For each experiment, 3.75×106 cells at a concentration of 1.5×107 cells/mL were electroporated at 250 V and 975 μF in the presence of 5 μg of pGL3EProb, a reporter gene construct for the androgen receptor gene promoter. As a control for transfection efficiency, 5 μg ph-RG-tk Renilla plasmid was used (Promega, Leiden, The Netherlands). After 48 h, transfected cells were treated with 0, 10, 20, or 50 μM curcumin for 24 h. At the end of the treatment, 75 μL of 106 cells/mL were placed in microtitration plates. In total, 75 μL of Dual-Glo™ Luciferase Reagent (Promega) were added to the cells for 10 min at 22°C and then 75 μL Dual-Glo™ Stop&Glo® Reagent (Promega) was added and incubated for 10 min at 22°C. Luciferase and Renilla activities were measured on an Orion microplate luminometer (Berthold Detection Systems). Results are expressed as a ratio of AR luciferase activity normalized to Renilla activity.
Real-time PCR
Reverse transcription (RT) was performed on 5 μg total RNA using oligo(dT) primers from the SuperScript™ III first strand synthesis system for RT-PCR (Invitrogen). Real-time PCR analyses were performed using the Power SYBR Green PCR Master Mix (BD Biosciences) and primers targeting androgen receptor (forward: GATAGCTACTCCGGACCTTACG and reverse: CAATGGGCAAAACATGGTC) and β-actin (forward: CTCTTCCAGCCTTCCTTCCT, reverse: AGCACTGTGTTGGCGTACAG). Real-time PCR were performed according to the manufacturer's instructions using a 7300 Real-Time PCR System (Applied Biosystems). Quantification was performed in triplicate, and expression levels were normalized using internal standards. Relative gene expression levels correspond to fold induction compared to untreated cells.
Results
Differentially expressed proteins in curcumin-treated 22Rv1 and PC-3 cells
The 2D DIGE-based proteomic analysis was performed to identify differentially expressed proteins in nuclear and cytosolic fractions extracted from 22Rv1 and PC-3 cell lines, treated with 20 μM curcumin for 24 h. Twelve gels were processed simultaneously (three gels per condition). We found a total of 425 spots to be differentially expressed between treated and untreated cells in 22Rv1 or PC-3 cell lines. A total of 192 spots were selected for identification by MALDI-TOF-MS, leading to successful identification of 93 spots. As several spots correspond to the same protein, we obtained a final list of 60 proteins differentially expressed in either 22Rv1 (32 proteins) and/or PC-3 (47 proteins) cells (Table 1). A total of 19 proteins were differentially expressed in both cell lines. Some were of interest as they were described to play roles in cancer progression. These were PP2R1A, HSP90B1, HSP90AB1, and HSPA9. Proteins were identified that were shown to be involved in drug resistance, MSN and RBM17; and cell death, HMGB1 (high mobility group protein B1) and NPM1.
Data are presented as signed fold change for significant (p<0.05) spots. When several spots correspond to the same protein, results are shown for all spots.
ns, nonsignificant.
Curcumin-enriched KEGG pathways
To find and contextualize potential missing links that could exist between our modulated proteins, and to summarize these results in relevant processes, we applied bioinformatics tools (Deng et al., 2010; Jiang et al., 2009). We identified curcumin-regulated KEGG pathways using DAVID enrichment tools with a threshold p-value of 0.05. Pathways affected in both cell lines were “Antigen processing and presentation” and “Spliceosome.” “Tight junction” and “Glycolysis” were impacted in 22Rv1 and PC-3 cells, respectively (Table 2).
Impact of curcumin treatment on KEGG pathways in 22Rv1 (A) and PC-3 (B) cells. Significantly affected pathways were retrieved from DAVID tool, using p≤0.05 as a cutoff. Pathways are sorted by decreasing fold enrichment.
Curcumin-enriched biological processes in GO
GO analysis of identified proteins, performed by using DAVID tools, provided an overview of the biological processes impacted by curcumin treatment. Significantly affected categories were selected using p≤0.005 as a cutoff and similar categories were grouped using Enrichment Map tool in Cytoscape® (Fig. 2 and Supplementary Table S1). “Response to protein stimulus and unfolding” and “protein folding” processes were impacted in both treated cell lines. Curcumin induced an overrepresentation of processes related to “cell death” and “apoptosis” in 22Rv1 but not in PC-3 cells. A strong impact of curcumin on RNA splicing-related processes was pointed out in PC-3 cells and was consistent with KEGG results shown above.
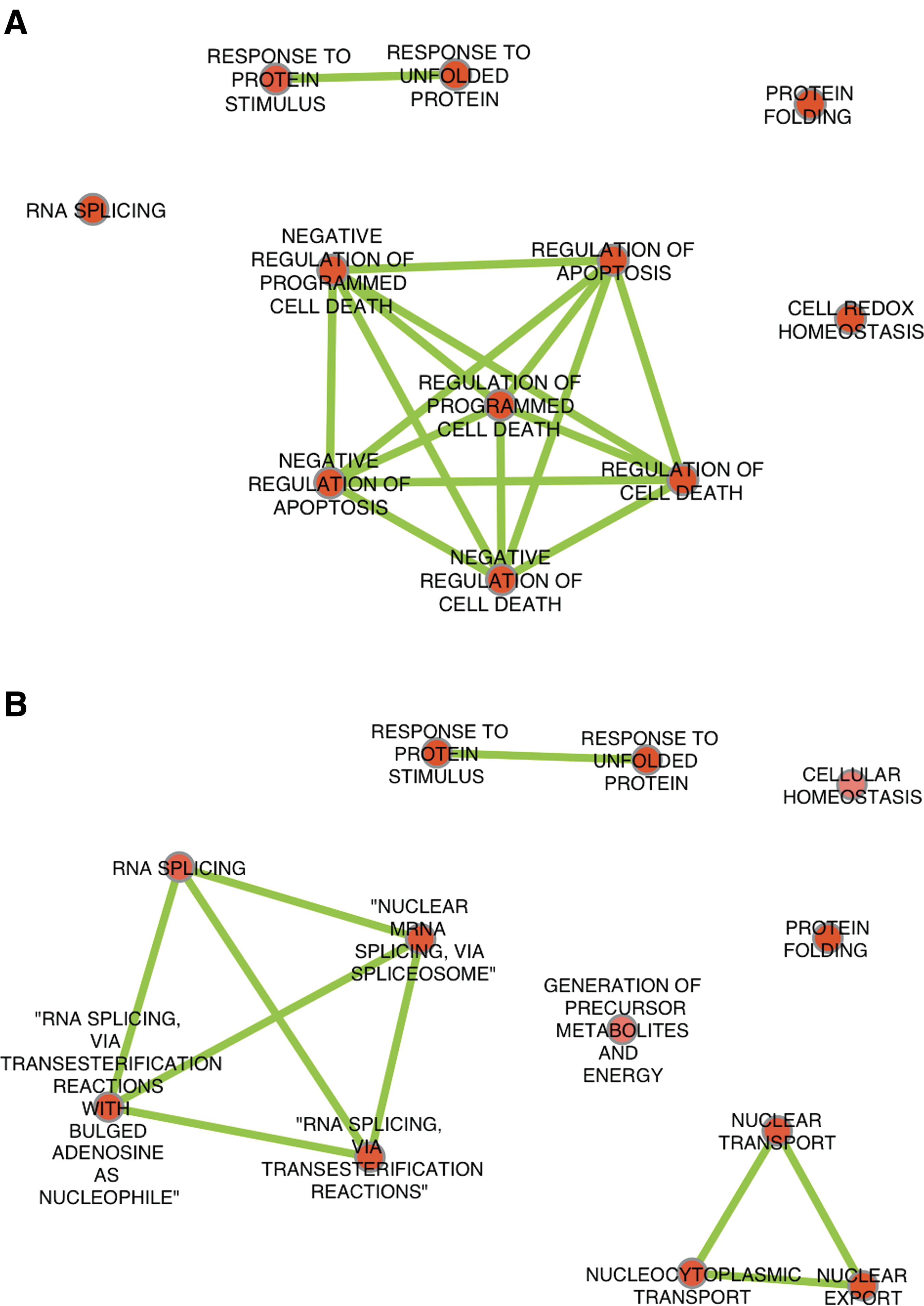
Impact of curcumin treatment on GO ontologies.
Potential regulators of curcumin
Subnetwork enrichment analysis in Pathway Studio was used to find potential expression regulators of curcumin-modulated proteins. We include a table of enrichment results in the supplementary data (Supplementary Table S2). To contextualize these results, the reported effects of curcumin on affected proteins or on their regulators were retrieved from the literature and added to the network. For the 22Rv1 cells, interactions with androgen receptor were also added. The networks obtained are illustrated in Figure 3.

Enriched regulators of proteins modulated by curcumin in prostate cancer cells, and their known relations with curcumin. (
Significant protein regulators that were modulated by curcumin in both 22Rv1 and PC-3 cells were MYC and HSF-1. Regulators specifically enriched in 22Rv1 cells were HSP90AA1, GJA1, PRKCE, XRCC6, MIR-141, and MIR-183. Regulators specifically enriched in PC-3 cells were IL6, INS, DDIT3, NDRG1, and MIR-152.
Curcumin modulates androgen receptor and cell death signaling
To validate the data showing the implication of curcumin on specific signaling pathways, we first evaluated the effect of curcumin on androgen receptor mRNA and protein expression and on AR transcriptional activity in prostate cancer cells. As shown in Figure 4, curcumin decreased the level of AR mRNA (Fig. 4A) and protein (Fig. 4B) expression in a dose-dependent manner (for concentrations higher than 10 μM) in both LNCaP and 22Rv1 androgen-dependent prostate cancer cells. Cells transfected with the androgen receptor reporter plasmid revealed that this natural compound inhibited androgen receptor transcriptional activity only in androgen-dependent LNCaP cells but not in androgen-independent PC-3 cells (Fig. 4C). We then evaluated the effect of curcumin on the level of expression of HMGB1 protein in 22Rv1 and PC-3 cells (Fig. 5). Curcumin increased the level of HMGB1 in 22Rv1 cells but did not impact the levels of this protein in PC-3 cells.

Modulation of androgen receptor signaling by curcumin. (

Modulation of cell death signaling by curcumin. The effect of curcumin on the level of expression of HMGB1 (high motility group protein B1) protein was analyzed by Western blot in 22Rv1 (left panel) and PC-3 (right panel) cells. Results are representative of at least three independent experiments.
Discussion
The chemopreventive potential of curcumin was previously described in prostate cancer (Teiten et al., 2010b). However, our study is the first to illustrate an overview of the impact of this natural compound on the prostate cancer proteome. The 2D-DIGE and mass spectroscopy approach allowed identification of 60 differentially expressed proteins after curcumin treatment of 22Rv1 (32 proteins) and PC-3 (47 proteins) prostate cancer cells. These results are in line with previous studies on curcumin (Fang et al., 2011; Madden et al., 2009) but pointed out a higher number of differentially expressed proteins, as we studied separately nuclear and cytosolic protein fractions and resolved protein spots often overlapping in total cell extracts. This large number of protein defined pathways influenced by the treatment, interaction of proteins, and identification of common regulators.
Correlation between 2D-identified and already published curcumin-affected proteins
Among the proteins we identified, several have already been described as curcumin targets. This first set of data highlighted the reliability of our 2D-DIGE approach, generating a global overview of proteins already known to be regulated by curcumin treatment. Bioinformatic analysis (Fig. 2) revealed that these proteins are mainly implicated in protein folding, RNA splicing and cell death (Supplementary Table S1). Identified proteins implicated in protein folding correspond to protein disulfide isomerases (PDIA6 and PDIA3) and heat-shock protein (HSP) family members, already shown to be implicated in prostate cancer progression (Ciocca et al., 2010; So et al., 2007). This modulation of HSP family members is consistent with previously published studies (Teiten et al., 2009b; Wu et al., 2006), and was also related to the anti-inflammatory potential of curcumin (Knowlton, 2006).
Our approach also pointed out proteins involved in cell death (HMGB1 and NPM1) (Bell et al., 2006; Tang et al., 2011; Weng et al., 2011), a process already described to be affected by curcumin (Reuter et al., 2008a; Teiten et al., 2010b). Our Western blot analysis confirmed that curcumin increased the level of expression of HMGB1 protein in 22Rv1 cells (Fig. 5). At high levels, NPM1 was also reported to alter AR functions such as androgen-dependent transcriptional activation in androgen-sensitive LNCaP prostate cancer cells (Leotoing et al., 2008). Peptidyl-prolyl cis–trans isomerase (FKBP4/FKBP52), assembled in AR complexes (Cheung-Flynn et al., 2005) and involved in protein folding because of its interaction with Hsp90, was also affected by curcumin treatment (Table 1 and Supplementary Table S1). As illustrated in Figure 3, several curcumin-affected proteins in 22Rv1 cells interact with the androgen receptor. The hypothesis that curcumin impacts AR function in 22Rv1 and LNCaP cells was supported by additional experiments showing the effect of curcumin on AR mRNA, protein expression and transcriptional activity (Fig. 4) and goes in line with published data (Choi et al., 2010). Moreover, pathway studio analysis identified a link between androgen receptor and PPP2R1A, a protein implicated in tight junction pathway and affected by curcumin treatment (Table 2). The effect of curcumin on this KEGG pathway was also related to its impact on PPP2CA, that plays a role in the Wingless (Wnt) signaling pathway implicated in cell proliferation (Bos et al., 2006) and adhesion (Brembeck et al., 2006). Results correlate with our previous work that showed that this natural compound modulates the Wnt signaling pathway in 22Rv1 cells (Teiten et al., 2011).
New proteins affected by curcumin treatment
After identification of differentially expressed spots, our study demonstrated novel proteins affected by curcumin treatment. GO analysis showed that nuclear transport (DDX39, PDIA3, NPM1, and BAT1 proteins) and RNA splicing (SPF45 and HNRNP proteins) are modulated by curcumin. BAT1 and DDX39 are implicated in mRNA export through the nuclear pore and interact with the transcriptional machinery; they are implicated in cell growth and division and in RNA splicing (Cordin et al., 2006; Sugiura et al., 2007). Curcumin also appeared to modulate expression of the human splicing factor SPF45 (RBM17) (Sampath et al., 2003) and of proteins belonging to the heterogeneous nuclear ribonucleoprotein family (HNRNPC, HNRNPH1, HNRNPF) (Carpenter et al., 2006) implicated in cellular proliferation and tumorigenesis. This natural compound also affects proteins associated with lack of response to chemotherapy thanks to its impact on RBM17, Thioredoxin reductase 1 (TXNRD1) (Nguyen et al., 2006) and moesin (a membrane-organizing extension spike protein) (Goler-Baron and Assaraf, 2011), three proteins reported to play a role in chemoresistance.
Potential regulators of curcumin-affected proteins
Our bioinformatics analysis also allowed us to identify potential protein regulators (Supplementary Table S2) within the mechanism of action of curcumin in prostate cancer.
We reported that MYC and HSF-1 could regulate several proteins modulated in both 22Rv1 and PC-3 cells. In 22Rv1 cells, GJA1, PRKCE, and XRCC6 appeared to be potential regulators of curcumin-modulated proteins. GJA1 (gap junction alpha-1 protein) protein, a component of the gap junction, already reported to be a target gene of the Wnt signaling pathway (Samarzija et al., 2009). PRKCE (Protein kinase C epsilon type) is involved in different cellular function (e.g., apoptosis, heat-shock response) and X-ray repair crosscomplementing protein 6 (XRCC6) is involved in DNA double-strand break repair. In the case of PC-3 cells, NDRG1 and DDIT3 could regulate curcumin-impacted proteins. NDRG1, a member of the N-myc downregulated gene family, was also reported to play an important role in tumor progression, aggressiveness and metastases in prostate cancer (Song et al., 2010). DNA damage-inducible transcript 3 (DDIT3) prevents the binding of C/EBP members to C/EBP sequences in the DNA. This regulator is implicated in adipogenesis, erythropoiesis, in the induction of growth arrest and in the endoplasmic reticulum stress response.
Finally our bioinformatic analysis highlighted three miRs—miR-141, -183, and -152—that could be regulated by curcumin in prostate cancer cells. These three miRs were already described as potential markers of prostate cancer (Gonzales et al., 2011), but never as curcumin-targets. MiR-141 is an androgen-regulated circulating miR that is upregulated in cancer and involved in the progression of prostate cancer (Waltering et al., 2011) through the regulation of TRPS1, involved in oxidative stress and in DU145 cell death by apoptosis (Albertini et al., 2011), but also of E2F transcription factor 3 (E2F3) and estrogen-related receptor gamma transcript variants 1, 2, and 3 (ESRRG) that control cell cycle, cell proliferation, and tumor suppressor proteins. MiR-183 was reported to be upregulated in prostate cancer (Schaefer et al., 2010) and in medulloblastomas associated with Wnt signaling pathway (Gokhale et al., 2010). It also plays a central role in the regulation of migration and metastasis in breast cancer (Lowery et al., 2010) and lung cancer by downregulating the expression of ezrin and other genes involved in metastasis (Wang et al., 2008). MiR-152 was identified to be a tumor suppressive miR as it regulates DNMT1 (Huang et al., 2010). It also appeared to be an immune system enhancer by upregulating natural killer cell-mediated cytolysis of host cells (Huang et al., 2010; Zhu et al., 2010).
These three prostate cancer progression-related miRs provide a first insight on the miR profile that has still to be identified for prostate cancer after curcumin treatment. Moreover, studies about the impact of curcumin on miRNAs is a hot topic in cancer research as several studies have been recently published on selected miRNAs in various cancer types excluding prostate (Mudduluru et al., 2011; Saini et al., 2011; Sun et al., 2008; Yang et al., 2010; Zhang et al., 2010).
Conclusion
For several years, numerous publications have shown that curcumin has the ability to reverse, inhibit, or prevent cancer formation and progression. By this 2D-DIGE approach, we have studied the overall impact of curcumin on two prostate cancer cell lines. Our results confirm already published data showing curcumin effect on proteins implicated in the protein folding process and AR pathway. Our results also highlight new processes affected such as RNA splicing, protein transport, and multidrug resistance, and suggest several proteins and three miRNAs (miR-141, -152, and -183) as potential regulators of processes modulated by curcumin. However, more experiments are needed to validate and understand the cellular impact that the modulation of these proteins would have as well as their chemopreventive potential roles in curcumin in the future.
Footnotes
Acknowledgments
M.H.T., A.G., S.C., and F.M. are supported by Télévie grants (Fonds National de la Recherche Scientifique, Belgium). Research at the Laboratoire de Biologie Moléculaire et Cellulaire du Cancer (LBMCC) is financially supported by “Recherche Cancer et Sang” foundation, by «Recherches Scientifiques Luxembourg» asbl, by “Een Häerz fir Kriibskrank Kanner” association, the Action Lions “Vaincre le Cancer” Luxembourg, The Fonds National de la Recherche Luxembourg, Télévie Luxembourg, and the Foundation for Scientific Cooperation between Germany and Luxemburg for additional support. Further support was received from the European Union (ITN “RedCat” 215009 and Interreg IVa project “Corena”). LNS Print costs were covered by the Fonds National de la Recherche (FNR) Luxembourg.
Author Disclosure Statement
The authors declare that no competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.