
Abstract
Abstract
The intracellular metabolic profile characterization of Schizochytrium sp. throughout docosahexaenoic acid fermentation was investigated using gas chromatography-mass spectrometry (GC-MS). Metabolite profiles originating from Schizochytrium sp. under normal and limited oxygen supply conditions were distinctive and distinguished by principal components analysis (PCA). A total of more than 60 intracellular metabolites were detected and quantified with the levels of some metabolites involved in central carbon metabolism varying throughout both processes. Both fermentation processes were differentiated into three main phases by principal components analysis. Potential biomarkers responsible for distinguishing the different fermentation phases were identified as glutamic acid, proline, glycine, alanine, and glucose. In addition, alanine, glutamic acid, glucose, inositol, ornithine, and galactose were found to make great contribution for dry cell weight and fatty acid composition during normal and limited oxygen supply fermentations. Furthermore, significantly higher levels of succinate and several amino acids in cells of limited oxygen supply fermentation revealed that they might play important roles in resisting oxygen deficiency and increasing DHA synthesis during the lipid accumulation. These findings provide novel insights into the metabolomic characteristics during docosahexaenoic acid fermentation processes by Schizochytrium sp.
Introduction
Schizochytrium sp. could produce about 35%–40% of their total fatty acids as DHA (Barclay et al., 2005; Yokochi et al., 1998). In order to achieve high lipid accumulation rate and high DHA content, the culture medium usually contains excessive carbon substrate and a limiting amount of other nutrients, such as nitrogen and phosphorus (Ratledg et al., 2002). Previous studies were mainly focused on improving the yield and productivity of DHA by modulating nutrition components and environmental factors (Ganuza et al., 2008; Zhu et al., 2007; Zhu et al., 2008) or reinforcing acetyl-CoA and NADPH supply (Ren et al., 2009). The parameters of biomass, lipid productivity, and DHA yield in different fermentation conditions were essential to evaluate the processes (Sijtsma et al., 2004). Since Schizochytrium species synthesize long-chain PUFAs via a polyunsaturated fatty acid synthase, oxygen supply is also one of the key factors limiting DHA content (Jakobsen et al., 2008).
Our previous studies reported the beneficial impact of the stepwise aeration control strategy on the performance of DHA production by Schizochytrium sp. Six fermentation parameters, including lipid yield coefficient and DHA yield coefficient, were used to examine the relationship between aeration and the fermentation characteristics (Ren et al., 2010). The effect of various oxygen supply conditions assessed by the volumetric oxygen mass transfer coefficient (KLa) was investigated in a 50-L bioreactor using fed-batch fermentation (Qu et al., 2010). These studies provided information on the biological processes involved in the adaptation to oxygen supply changes and their relationship with the agitation condition. However, the potential effect of oxygen availability on the intracellular metabolite levels remained to be elucidated. Relatively little was known about the variations of the primary cell metabolism that result in the environmental stress.
Metabolomics, focusing on comprehensive metabolites analysis in a biological system, was increasingly incorporated in determining the cellular stress responses (Koek et al., 2011; Mashego et al., 2007). Metabolites are not gene products, but their concentrations are affected by the activities of gene products (enzymes) and the availability of precursors (Villas-Boas et al., 2007). Because of this, it was decided that metabolic profile analysis would be a good choice to investigate the cellular responses to environmental changes and nutritional stresses at the metabolic level.
Metabolomics technology was utilized to study the responses of Schizochytrium sp. to oxygen stress condition. Considering the advantages of its relatively high reproducibility, high resolution, high-throughput analysis and good sensitivity, gas chromatography–mass spectrometry (GC-MS) was applied to detect the changes of intracellular metabolites (Lisec et al., 2006; Villas-Boas et al., 2012). In addition, principal components analysis (PCA) of intracellular metabolites was performed to distinguish the biomarkers associated with cell growth, lipid accumulation, and DHA production. Metabolites related to amino acid biosynthetic pathway and central carbon metabolism indicated significant changes during both processes.
Material and Methods
Microorganism
Schizochytrium sp. HX–308 (CCTCC M 209059), which was isolated from seawater and stored in the China Center for Type Culture Collection (CCTCC) (Ren et al., 2009), was used in the present study. This strain was preserved in 20% (v/v) glycerol at −80°C.
Fermentation conditions
The seed culture medium and the conditions were as indicated in our previous study (Ren et al., 2009). The culture preserved in the glycerine tube was inoculated into a 250-mL flask with 50 mL medium and cultivated for 24 h. After a three generation cultivation, the preculture was inoculated into 100 mL of medium in 500-mL shaking flasks with initial glucose concentration 120 g/L and 4% (v/v) inoculum size and incubated at 25°C. The shaker speed in normal oxygen supply condition was 150 rpm, while a speed of 120 rpm was used in the limited oxygen supply condition. The sulfite method (Na2SO3 method) was proposed for the measurement of oxygen transfer coefficients (kLa) in flasks (Puskeiler et al., 2005).
Analyses of residual glucose, biomass, and lipid
The concentration of glucose in the fermentation broth was measured by a bioanalyzer (SBA-40C, Institute of Biology, Shandong Academy of Sciences, China). Broth (10 mL) was used to determine cell dry weight by gravimetric method. The methods of lipid extraction, fatty acid methyl esters (FAMEs) preparation and analysis were according to our previous study (Ren et al., 2010).
Sampling, quenching, and extraction of intracellular metabolites
In order to compare the metabolic profiles of Schizochytrium sp. at different fermentation stages, cells were sampled every 12 h during the fermentation. Cells were quenched and extracted according to Ding and Yuan (2009a) with slight modifications. The samples were sprayed into cold 60% methanol (-40°C, v/v) to arrest the metabolism instantaneously. After centrifugation at 4000 g at −10°C for 3 min, cells were washed twice with cold 0.9% (w/v) sodium chloride solution. The centrifugation was repeated after each washing step. Cells were then ground into a fine powder in liquid nitrogen, and cell powder (100 mg) was first suspended with 1 mL 60% methanol (−40°C, v/v) and mixed thoroughly. The mixture was frozen and thawed for three times. After centrifugation at 8000 g at −10°C for 10 min, the cell pellet was re-extracted with 0.5 mL of extraction buffer, and the extract was combined with the former one.
To correct for minor variations occurring during sample preparation and analysis, ribitol (2 mg/mL in water, 20 μL) was used as an internal standard. 50 μL of the methanol/water phase were transferred into a sample glass vial and dried under a N2 stream using a sample concentrator (MD200-1, Aosheng Company, China) (Liu et al., 2012). Two-stage chemical derivatization was performed on the residue according to Ding (2009b).
Metabolome analysis by GC-MS
The sample was analyzed by GC-MS as described previously (Strelkov et al., 2004). The GC-MS system consisted of a Finnigan Trace gas chromatograph and a Trace mass spectrometer on electron ionization (EI) operated at 70 eV (Thermo Finnigan, San Jose, CA). One microliter sample was injected into a DB-5MS capillary column coated with 5% Phenyl and 95% methylpolysiloxane (30 m×0.25 mm i.d.×0.25 μm thickness) in splitless mode. The column temperature was held for 2 min delay at 70°C, and then increased to 290°C with the rate of 8°C/min, holding for 3 min. Helium was used as carrier gas and the flow was constantly at 1 mL/min. The transfer line and ion source temperatures were 280°C and 250°C, respectively. The mass scan range was 50–600 m/z.
The metabolites were identified by processing the raw GC-MS data with the AMDIS (v.2.1) software of NIST (National Institute of Standards and Technology, Gaithersburg, MD). The two-dimensional comparison of recorded spectra and retention indices (RI) with library entries resulted in identification of good confidence. The area of each acquired peak was normalized against that of the internal standard for further data processing.
Multivariate statistical analysis
After preprocessed by mean-centering and scaling, PCA was carried out to analyze the dataset with STATISTICA (Kind et al., 2007) and SPSS software (SPSS Inc., v. 18.0, Chicago, IL, USA). In the PCA score plot, each data point represents a linear combination of all the metabolites from each sample. This analysis allowed the clustering of biological samples based on their similarities and differences in the metabolites dataset. Distances between the groups of samples give a measure of the overall difference among the metabolic profiles of the strains with different treatments.
Results
Effect of oxygen supply on growth and fermentation of Schizochytrium sp.
The fermentation characteristics under two different oxygen supply strategies of Schizochytrium sp. were investigated. The value of kLa at 120 rpm was only 25 s−1, while at 150 rpm the value reached 68.17 s−1, so the two speeds represented the normal and limited oxygen supply conditions. Figure 1 indicates the significant effect of the dissolved oxygen levels on glucose consumption, cell growth, and lipid content of Schizochytrium sp. HX-308. There were three phases including cell growth, main, and final fermentation phases, according to cell growth and lipid accumulation during the fermentation processes (Ren et al., 2010). Cell growth rate of the culture under limited oxygen supply condition was obviously slower than that under normal condition, while the fermentation period prolonged 48 hours than normal situation. It can be seen from the data that the oxygen supply had no clear impact on lipid accumulation, but the final dry cell weight decreased from 46.59 g/L to 36.18 g/L. In addition, the slower glucose consumption rate under oxygen limitation condition resulted in the higher lipid proportion of dry cell weight.

Docosahexaenoic acid fermentation with high-yielding mutant Schizochytrium sp. HX-308 at various dissolved levels. Normal oxygen supply condition: square (▲), glucose; circle (●), dry cell weight; down triangle (▼), lipid; limited oxygen supply condition: square (△), glucose; circle (◯), dry cell weight; down triangle (▽), lipid.
The changes of fatty acid composition of Schizochytrium sp. HX-308 in response to different oxygen supply conditions were also analyzed (Table 1). Under limited oxygen supply condition, the DHA proportion was obviously higher than that under normal condition. Furthermore, the DHA proportion maintained above 46% and the highest proportion reached 49.3% at 60 h. At the same time, the amount of C14:0 and DPA (C22:5), which were the major components of by-products in the fatty acids, increased slightly in normal oxygen supply condition. All the variations indicated that low dissolved oxygen level could promote DHA synthesis. On the other hand, the proportion of C14:0 was always below 10% during the fermentation processes, and the proportion of C16:0 decreased notably along with the increment of DHA.
Metabolomic profiling of Schizochytrium sp. with different oxygen supply conditions
GC-MS was applied here formetabolic profiling in Schizochytrium sp. fermentations with different oxygen supply levels. More than 60 putative intracellular metabolites were detected (Supplementary Table S1; supplementary data are available online at www.liebertpub.com/omi) and 27 of them were identified and quantified in all samples at different fermentation stages. As was shown in Table 2, identified metabolites mainly included the intermediates from central carbon metabolism, such as amino acids, organic acids, alcohols, as well as sugars. Figure 2 showed the total ion chromatogram (TIC) of samples from the two conditions, illustrating the significant variations of metabolic profiling under different oxygen supply conditions.

Total ion current (TIC) chromatograms of Schizochytrium sp. HX-308 during docosahexaenoic acid fermentation under various oxygen supply conditions.
RT, retention time.
Multivariate statistical analysis
The variations of glucose consumption and fatty acid composition indicated that there were significant differences in cellular behaviors and living environments of Schizochytrium sp. under the two oxygen supply conditions. Comparative metabolomic analysis could be useful for insights into the possible reasons for these changes. So the metabolic profile dataset was statistically analyzed by PCA to gain insight into the nature of the multivariate data and evaluate biological alteration. The metabolic profiles of Schizochytrium sp. from both processes were analyzed together by PCA (Fig. 3a and 3b). The maximum variability in the dataset differentiated precisely between the normal and limited oxygen supply conditions, with the first component (PC1) covering 29.42% of the data variance. The second principal component (PC2) and the third component (PC3) explained 20.08% and 11.58%, respectively, of the total detected metabolites pools variance. The PCA score were calculated for the first two PCs, the trajectory illustrated that the trend of metabolic profiles for both fermentation processes similarly changed during the first 36 h, then presented a significantly difference. Therefore, different dissolved oxygen levels exerted an influence on cellular metabolism, especially after the cell growth phase. Alanine, glutamic acid, and glucose were the metabolites with the highest contribution to the variance in PC1, while inositol, ornithine, and galactose were the highest ones in PC2. They contributed most for the differences of DHA synthesis and dry cell weight, and distinguished normal and limited oxygen supply conditions more significantly.
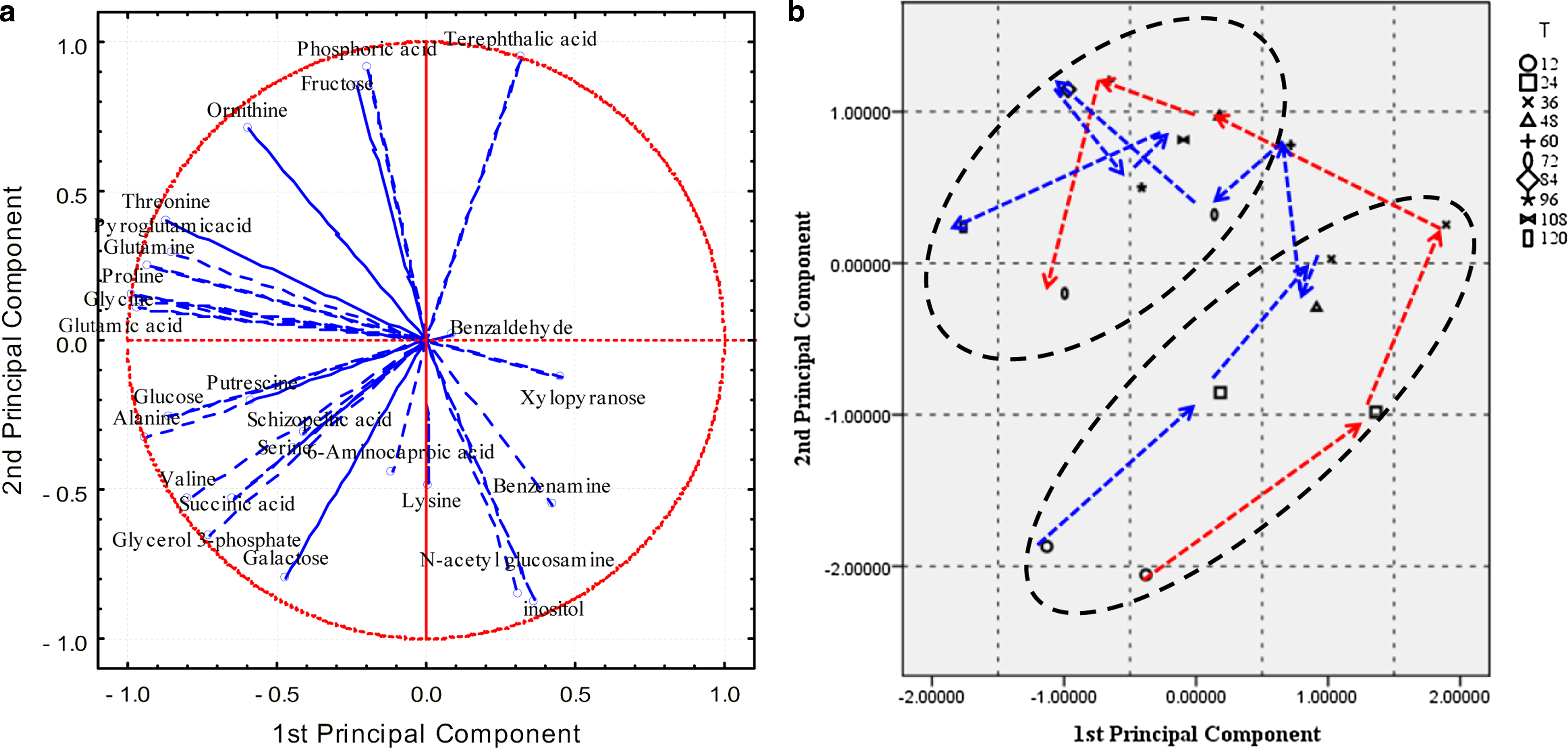
Principal component analysis (PCA) loading plot (
Further analysis was conducted for the samples from each fermentation process respectively. The score plots illustrated that three groups were observed in accordance with different fermentation phases (Fig. 3d). For normal oxygen supply condition, samples clearly separated from each other, forming distinct groups and representing cell growth (group A), the main (group B), and the final (group C) fermentation phases. A similar result was also found out in oxygen limitation fermentation (Fig. 3f). As shown in the loading plots (Fig. 3c and Fig. 3e), glutamic acid, proline, glycine, alanine, and glucose were found to make great contribution to distinguishing different fermentation phases of samples from both fermentation processes, so they were more conducive to lipid accumulation rate and fatty acid composition.
Metabolic differences between normal oxygen supply and limited oxygen supply conditions
Apart from performing multivariate data analysis to examine the global variations between different experimental conditions, the change of individual compound was also analyzed in greater detail. Through analyzing the metabolites differentiation, the intermediates of glycolysis, tricarboxylic acid (TCA) cycle, especially amino acid biosynthesis were identified to be of particular importance (Supplementary Table S2).
The levels of metabolites involved in central carbon metabolism under different oxygen supply strategies at different fermentation stages were shown in Figure 4. Succinate, which was the only identified one in TCA intermediates, presented the similar variation tendency under both processes. However, after 48 h the concentration of succinate in limited oxygen supply condition was significantly higher than that in normal condition.

Changes of detected intermediates of glycolysis, TCA cycle, and glycerol biosynthesis in Schizochytrium sp. HX-308 during normal oxygen supply and oxygen-limited fermentation processes. The y-axis was relative abundance which was calculated by normalization of peak area of each metabolite to internal standard.
Higher intracellular levels of several measured amino acids (e.g., serine and alanine) were characterized at the limited condition. Particularly the level of ornithine strongly increased, reaching 8.01-fold of that in normal condition. On the other hand, in normal oxygen supply condition, the cellular contents of phosphoric acid, inositol, and lysine could reach 1.21-, 0.82-, and 3.84-fold higher than those in the limited oxygen supply fermentation.
The variations of measured metabolites were further viewed in a heat-map of clustering analysis. As shown in Figure 5, these metabolites could be classified into two clusters. Cluster A included the metabolites that exhibited higher levels and great changes during the fermentation processes. The levels of polyols in cluster B maintained at a relatively stable level. Amino acids exhibited distinct regular patterns during different fermentation processes. Under normal oxygen supply condition, the amount of alanine and proline raised before 36 h, and then declined until the end of fermentation. Other amino acids did not represent obvious trend, but all of them decreased at the final fermentation phase. On the contrast, during limited oxygen supply fermentation, most detected amino acids emerged at the lowest level at 84 h, except pyroglutamine, serine, and glycine. The heat-map also revealed that the levels of most amino acids with oxygen limitation were much higher relative to normal oxygen supply after 36 h. Furthermore, total amino acid content difference was consistent with the result of heat-map analysis (Figure 6).
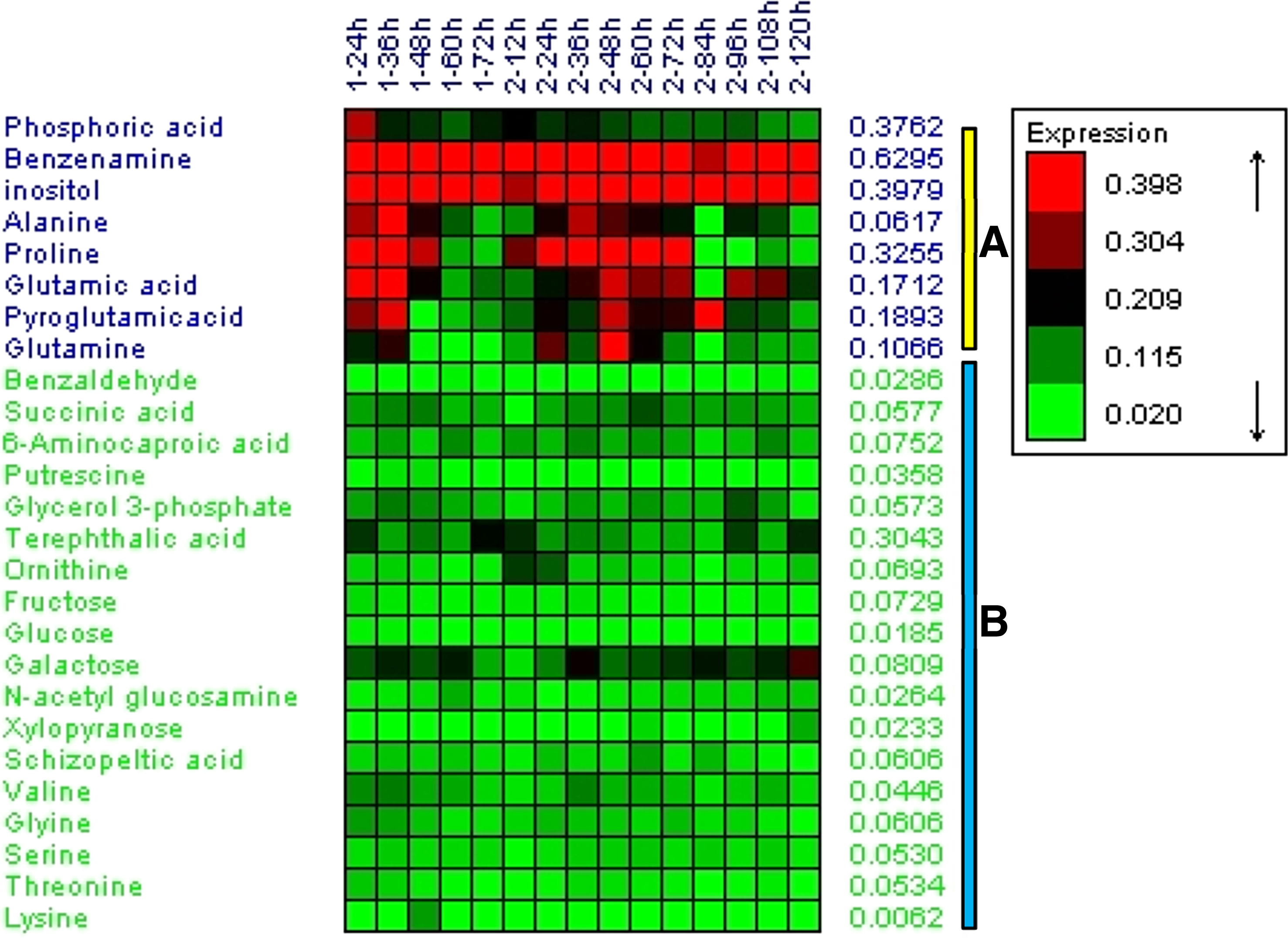
Heat map representation of metabolites in Schizochytrium sp. HX-308. Shades of red and green represent an increase and decrease of a metabolite during normal oxygen supply and limited oxygen supply fermentation processes.

Total amino acids pool in Schizochytrium sp. HX-308 in response to oxygen stress. The relative abundance was calculated by normalization of peak area of each metabolite to internal standard ribitol, and each value represents the mean of independent replicates±SD.
Discussion
In this study, we detected the fermentation parameters and intracellular metabolites of Schizochytrium sp. under normal and limited oxygen supply conditions, and then compared the metabolic profiling by PCA and heat-map between the samples from both fermentation processes.
Fermentations under different oxygen supply conditions were carried out to evaluate the impact of oxygen levels on metabolic regulation. Therefore, Schizochytrium sp., which was exposed to an oxygen limitation condition, could adjust its metabolism to defend and adapt to the variable condition by evoking stress responses, including rapid synthesis of protective molecules (e.g., serine and alanine) and activation of signal transduction pathways (e.g., glutamic acid and glutamine biosynthetic pathways). Our results suggested that the changing dissolved oxygen concentration had a great effect on growth rate, and further affected cell metabolism via a series of interactions.
Oxygen strategy induced the levels of most detected amino acids to change significantly, which seemed to be responsible for the variety of some metabolites involved in glycolysis and TCA cycle. Phosphoric acid was one of the most important metabolites and participated in regulating some signal transduction pathways. In our study, its level declined remarkably in limited oxygen supply condition (Fig. 7), which might be ascribed to less participation in phosphorylation converting ADP to ATP in glycolysis and oxidizing citrate in TCA cycle. On the other side, cells might exploit phosphoric acid to activate the proteases for regulating the signal transduction pathway, so the growth can adjust to the environmental stress. Our results implied that the phosphorylation level was more activated in normal oxygen supply fermentation, so it might be related to the higher dry cell weight and faster growth rate.

Differences of phosphoric acid under normal and limited oxygen supply conditions. The relative abundance was calculated by normalization of peak area of each metabolite to internal standard ribitol, and each value represents the mean of independent replicates±SD.
For instance, some amino acids (glycine and valine) derived from pyruvate, 3-phosphoglycerate, and malate increased significantly in the final fermentation stage, while those derived from α-ketoglutarate reduced under limited oxygen supply condition (Figs. 5 and 6). Remarkably, such changes were possibly caused by reduced oxygen levels as the fermentation progressed. Furthermore, higher levels of amino acids after 36 h under limited oxygen supply condition revealed that nitrogen metabolism was more active. Amino acids were essential parts of carbon and nitrogen metabolism, and also precursors for a number of cell components such as proteins and nucleotides. Fritz (2006) suggested that analysis on amino acids provided information on the relationship between carbon–nitrogen status and amino acid metabolism).
Among the detected amino acids, higher level of pyroglutamic acid under limited oxygen supply condition was observed. Orlowski (1969) reported that pyroglutamic acid was a signal for activation of glutathione metabolism, so the glutathione metabolism was more activated by the oxygen stress. Glutamic acid, which was considered to be activated in response to acid stress (Natera et al., 2006) and osmotic stress (Yap et al., 1983) of bacteria, represented a similar change with pyroglutamic acid. In addition, the carbon flux to the glutamic acid and glutamine biosynthetic pathway was significantly increased at lower oxygen availability. Glutamic acid and glutamine play essential roles in cellular metabolism in microorganisms (Chen et al., 2010). Judging from our result (Fig. 4), glutamine was also presumed to act as the oxygen stress protectant. Proline was synthesized from glutamic acid (Brandriss and Falvey et al., 1992) and could be converted to glutamic acid (Brandriss et al., 1983). It was an important amino acid in terms of its biological functions for glutamic acid various stress conditions. Wethered and co-workers (1985) reported that proline of Thraustochytrium aureum and Thraustochytrium roseum decreased linearly with the external salt concentration and contributed to the osmotic pressure. In our result, similar to glutamic acid, proline exhibited a quite high level before 84 h in oxygen stress, and then decreased rapidly to zero after adaptation to limited oxygen supply condition. They could be important regulatory substances for Schizochytrium sp. to resist oxygen deficiency and increase DHA synthesis during the lipid accumulation.
Polyols were also considered as compatible solutes in response to several environmental stresses, as well as myo-inositol was one of the widely studied polyols (Ding et al., 2010). Ding et al. (2009a) suggested that intracellular myo-inositol content of S. cerevisiae was considered as the oxygen stress protectant under vacuum conditions. Moreover, in our study myo-inositol under oxygen limitation obtained was lower than that under the normal condition, which reflected that more myo-inositol was used for defending the stress in Schizochytrium sp. On the other hand, myo-inositol is the direct precursor of phosphatidylinositol (PI) (Nikawa et al., 1997). As a result, the decreasing level of myo-inositol after the adaptation of Schizochytrium sp. to the oxygen stress might be due to the formation of PI for regulation of several cellular processes, including membrane trafficking and few nuclear functions by evoking inositide signaling pathways (York et al., 2006).
Our results demonstrate that a limited oxygen supply condition imposed a substantially stressed environment for Schizochytrium sp. growth and metabolism. It was speculated that Schizochytrium sp. accumulated protective compounds such as succinate and several kinds of amino acids to be able to survive in such a condition. On the other hand, reduction of phosphoric acid and inositol might promote the synthesis and accumulation of lipid and increase the DHA proportion in fatty acids.
Conclusions
Taken together, our results indicated that metabolomics was a good tool for analyzing the significant metabolic distinction between the normal and limited oxygen fermentations. The time-resolved analysis of the metabolomic responses was investigated to understand metabolic perturbation and the subsequent trend of returning to cellular homeostasis. Oxygen stress was proven to be the obvious factor influencing the metabolic trajectory and growth pattern of Schizochytrium sp. Furthermore, the observed variations of intracellular metabolites led to a better understanding of different metabolic status of Schizochytrium sp. during fermentation conditions or involvement in oxygen stress response. This study has highlighted the benefits of GC/MS-based metabolomic strategy to gain new insight into the response mechanisms of Schizochytrium sp. to environmental stress. In addition, we can further research the regulation mechanism of representative metabolites such as phosphoric acid, inositol, and amino acids on DHA fermentation by Schizochytrium sp.
Footnotes
Acknowledgments
This work was financially supported by the National Science Foundation for Distinguished Young Scholars of China (No. 21225626), the National Basic Research Program of China (No. 2009CB724700), the Nature Science Foundation of Jiangsu Province (No. BK2012424), the National Science and Technology Pillar Program (No. 2011BAD23B03), and the National High Technology Research and Development Program of China (No. 2012AA021704). We gratefully acknowledge the contributions of Mengqi Zhang and his classmate who helped to revise the manuscript.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.