
Abstract
Abstract
DNA methylation, histone modifications, and chromatin configuration are crucially important in the regulation of gene expression. Among these epigenetic mechanisms, silencing the expression of certain genes depending on developmental stage and tissue specificity is a key repressive system in genome programming. Polycomb (Pc) proteins play roles in gene silencing through different mechanisms. These proteins act in complexes and govern the histone methylation profiles of a large number of genes that regulate various cellular pathways. This review focuses on two main Pc complexes, Pc repressive complexes 1 and 2, and their phylogenetic relationship, structures, and function. The dynamic roles of these complexes in silencing will be discussed herein, with a focus on the recruitment of Pc complexes to target genes and the key factors involved in their recruitment.
Introduction
Polycomb Complexes and their Biological Roles
PcG proteins form conserved regulatory structures that can suppress genes through a variety of physiological roles and types of epigenetic patterning in higher organisms (Pirrotta, 2006). Their functions mostly emerge in particular spatial patterns. Margueron and Reinberg (2011) defined the Pc complexes as a set of genes that, when mutated, cause incorrect body segmentation, similar to the initial Polycomb mutant phenotype that has been described in Drosophila. For instance, improper reactivation of pluripotent genes such as the Hox (homeobox) genes initiates carcinogenesis (for a review, see Ringrose and Paro, 2004). In Drosophila, Hox genes are active during early embryogenesis and are maintained during adult life via epigenetic mechanisms. Clustered on chromosomes (Garcia-Fernandez, 2005), the Hox gene family is important for the proper positioning of segmented structures along the body axes (Deschamps and van Nes, 2005). Crosstalk between PcG (Bantignies et al., 2011; Cao et al., 2005) and the trithorax (Petruk et al., 2001; Salvaing et al., 2006) genes was first discovered in Drosophila melanogaster. Hox genes are critical developmental regulators and are expressed in distinct regions of the anterior–posterior axis (Alexander et al., 2009). Mutations in the Hox genes have been associated with various developmental disorders, such as limb malformations (Kmita et al., 2005; Zakany and Duboule, 2007) and neural crest defects (McNulty et al., 2005). The Hox genes are also specifically expressed in the allantois of placental mammals (Scotti and Kmita, 2012; Shaut et al., 2008). Several studies have provided evidence for the roles of Hox genes in oncogenesis (for a review, see Shah and Sukumar, 2010). For instance, overexpression of certain Hox genes has been linked to oral cancer (De Souza Setubal Destro et al., 2010) and breast cancer (Hayashida et al., 2010). However, a full account of the interplay between PcG proteins and trithorax group proteins during development and carcinogenesis is beyond the scope of this article. For a better understanding of histone modifications and chromatin, a number of excellent reviews (Grimaud et al., 2006; Ringrose and Paro, 2004; Schuettengruber et al., 2007) focus on these subjects. Moreover, Pc complexes contribute crucially during embryogenesis, tissue differentiation (Pietersen and van Lohuizen, 2008; Sparmann and van Lohuizen, 2006) and tumorigenesis (Schlesinger et al., 2007; Widschwendter et al., 2007) through X-chromosome inactivation (Casanova et al., 2011; Splinter et al., 2011) and regulation of imprinted genes (Jullien et al., 2006; Makarevich et al., 2006; Schubert et al., 2006). Pc complexes are also associated with nuclear reprogramming and chromatin remodeling (for a review, see Golbabapour et al., 2011).
Polycomb Complexes in Action
Histone modifications and chromatin remodeling are two main epigenetic mechanisms in the regulation of gene expression. The nucleosome, the basic unit of chromatin, consists of a histone octamer, which contains a pair of each of the core histone proteins (H2A, H2B, H3, and H4) (Luger et al., 1997). Nucleosomes consist of 147 bp of DNA wrapped around the core histone and are attached to one another by DNA strands and the histone protein H1 (Luger et al., 1997). Histone modifications consist of any post-translational alterations, such as histone methylation and histone acetylation, imposed on histone proteins. These modifications, in turn, define the configuration of chromatin and its accessibility to transcriptional machinery (for a review, see Golbabapour et al., 2011). Methylation of H3K9 is important in constitutive heterochromatin, while that of H3K27 is a key repressive factor for the regulation of developmental genes (for a review, see Alabert and Groth, 2012).
Studies addressing the mammalian PcG family (for a review, see Jones et al., 2000) suggest a variety of tasks for Pc complexes, such as tasks related to proliferative defects in lymphoid cells (Core et al., 1997; Jacobs et al., 1999) and sex determination (Katoh-Fukui et al., 1998). These complexes contribute to silencing through histone modification. Differences in the expression of each component of Pc repressive complexes (PRCs) suggest that the composition of Pc complexes is specific to cell type and/or developmental stage (Gunster et al., 2001), which could be referred to as dynamic regulation of Pc complexes (for a review, see Lange et al., 2011). PcG proteins also contribute to the covalent post-translational modification of histones. Pc complexes repress gene expression through methylation of histone H3 (H3K27 and H3K9) (Cao and Zhang, 2004a; Lindroth et al., 2004) and ubiquitination of histone H2A (Wang et al., 2004).
Components of Polycomb Complexes
In 1985, the first Pc protein was reported by Jurgens (1985). Since then, attention has been focused on understanding its role and characteristics. Pc complexes primarily proposed as a silencing mechanism for regulating homeotic genes in Drosophila (Chan et al., 1994). Initial studies of Pc complexes showed that these complexes could limit the accessibility of homeotic genes to the transcriptional machinery through spatial modification (for a review, see Cunliffe, 2003). PcG proteins bind nucleosomes and alter the intrinsic structure of chromatin to initiate epigenetic modifications and maintain these modifications during development (for a review, see McBryant et al., 2006). These multi-protein complexes modify chromatin structure to form flexible, repressive chromatin configurations that include numerous targeted genes and maintain silencing (for a review, see Morey and Helin, 2010, and Papp and Plath, 2011). Pc response elements (PREs) are regulatory elements with which PcG complexes interact (Chan et al., 1994; Pirrotta, 1998).
Mammalian PcG proteins have been shown to exist in several types of complexes with different components and structural configurations (Satijn et al., 1997). These complexes are composed of different nonperiodic repetitive units. Complexes of PcG proteins are mainly combinations of multi-protein structures. PRC1 consists of four core proteins: Pc [also known as Chromobox (Cbx) in mammals], polyhomeotic (Ph), posterior sex combs (Psc), and sex combs extra (Ring) (Francis et al., 2001; Saurin et al., 2001; Shao et al., 1999). PRC2 is comprised of three main, strongly conserved subcomplexes: Esc and Enhancer of Zeste [E(z)]; Suppressor of Zeste 12 (Suz12); and RbAp48/Nurf-55 (Cao et al., 2002; Kuzmichev et al., 2002; Muller et al., 2002). There is some evidence that PRC2 is composed of more than just these four components (Cao and Zhang, 2004b; Kim et al., 2009; Li et al., 2010a; Pasini et al., 2010). Ring proteins consist of various domains such as dRing plus two additional conserved cores: Ring1A and Ring1B. Ring1 mediates the ubiquitylation of histone H2A (de Napoles et al., 2004) (for a review, see Martinez and Cavalli, 2006). PRC1 recognizes H3K27me3 to inhibit transcriptional elongation through H2A ubiquitylation (Zhou et al., 2008) and to compact the chromatin structure (Eskeland et al., 2010). PRC2 methylates H3K27 (a key chromatin mark), a main feature of chromatin silencing mediated by PcG proteins. In mammals, yin and yang 1 (Yy1) (Thomas and Seto, 1999), homologues of Drosophila Pho, contribute to histone methylation by recruiting Ez homolog 2 (Ezh2) to H3K27 (Brown et al., 1998). A schematic representation of the components of PRC1 and PRC2 is illustrated in Figure 1. The main components of PRC1 and PRC2 in Drosophila, mouse and human are summarized in Table 1. Several Pc complexes, such as PRC1 and PRC2 (for a review, see Lund and van Lohuizen, 2004), pleiohomeotic repressive complex (PhoRC), and Pc repressive deubiquitinase, are defined as PcG proteins. PcG proteins accumulate widely along chromosomes (Decamillis et al., 1992; Zink and Paro, 1989) and are recruited to PREs in association with transcription factors (for a review, see Ringrose and Paro, 2007) and DNA binding proteins such as Pho (Oktaba et al., 2008). In mammals, several transcription factors are involved in the recruitment of PcG proteins (Bracken and Helin, 2009), which has led to conflicting findings about PRE motifs, protein interaction sites that bring together different proteins in multi-protein complexes and locate them within heterochromatin (Brasher et al., 2000).

General constituents of Polycomb repression complexes (PRC1 and PRC2). PRC1 consists of Cbx, BMI1, Ring, Ph, and Pc proteins. PRC2 consists of Ezh2, Eed, Suz12, SET, and RBBP4.
Chromodomains, conserved protein motifs, are shared regions of homology between two chromatin regulators, the Pc proteins and heterochromatin protein 1 (HP1) (Paro and Hogness, 1991). Bracken and Helin (2009) proposed that so-called “cell fate transcription factors” are crucial to the recruitment of PcG proteins and targeting genes. Moreover, noncoding RNAs (ncRNAs) might mediate the recruitment of PcG members through cell fate transcription factors.
Evolution of Polycomb Complexes
During the past decade, PcG proteins have been shown to participate in a multitude of biological tasks, from stem cell regulation to differentiation, with conserved mechanisms of regulation (Muller et al., 1995). An evolutionary perspective of PcG proteins provides useful insight into their function. As highly conserved biological structures, these proteins are found in various organisms and have conserved biological activities. Figure 2 illustrates the phylogenetic tree of selected organisms (Drosophila, mouse, rat and human) and their PRC1, PRC2, and Cbx homologs (see Supplementary Fig. S1 and Supplementary Table S1 at www.liebertonline.com/omi). The mechanism underlying gene silencing by Pc complexes is conferred by their roles in the structural modification of chromatin and the post-translational modification of histones (for a review, see Margueron and Reinberg, 2011).

Polycomb Complexes during Embryogenesis and Development
Developmental studies have shown that PcG proteins bind to and interact with Pc domains in more than 200 PcG target genes (Schwartz et al., 2006) and dynamically regulate their expression (Oktaba et al., 2008). For biological activity, PcG proteins need to be part of complexes consisting of core components and associated components. During development, the responses of differentiated cells to environmental signals change due to the epigenetic regulatory effects of PcG members (Paro and Hogness, 1991). As reviewed by Cunliffe (2003), Pc complexes act via a cellular memory mechanism through which cell behavior can be controlled. During the very early steps following fertilization in Drosophila, activation of homeotic genes occurs in certain segments, whereas homeotic genes are repressed in the remainder of the segments (Chan et al., 1994). The suppression of homeotic genes is maintained by a PcG-mediated mechanism that contributes to regulation in the cis-regulatory region (Felsenfeld and Groudine, 2003). During development in female mammals (presence of two X chromosomes), one X chromosome is inactivated to equalize the expression of genes to a level that is equivalent to that of males, who have only one X chromosome (for a review, see Tie et al., 2003). A small ncRNA within Xist (a gene that is generally expressed from and accumulates on the inactive X chromosome) targets PRC2 and contributes to determining chromatin configuration (Brown et al., 1992; Lucchesi et al., 2005; Zhao et al., 2008).
Polycomb Complexes in Cancer
In a variety of cancers, such as lymphoma (McCabe et al., 2012; Tonini et al., 2008), bladder cancer (Hinz et al., 2007), breast cancer and prostate cancer (Ren et al., 2012), PcG proteins repress tumor suppressor genes (Dietrich et al., 2007; Gil and Peters, 2006) through the accumulation of DNA methylation in the promoter regions of their targets, thus altering their methylation profiles (Ohm et al., 2007; Schlesinger et al., 2007). DNA methylation, the addition of a methyl group to a cytosine to form 5-methylcytosine, is a hallmark for recruiting epigenetic factors to suppress gene expression and will be discussed later in this review. Studies on abnormal silencing patterns (Ben-Porath et al., 2008; Bracken and Helin, 2009; Schlesinger et al., 2007; Widschwendter et al., 2007) in both pro- and anti-proliferative genes indicate potential cooperation between PcG proteins and DNA methylation enzymes in silencing the expression of genes (Bracken and Helin, 2009). Abnormal functions of Pc complexes appear to be the main factor involved in a variety of different types of cancerous proliferation and may be due to abnormal expression of the PcG genes themselves (mostly upregulation) (Sparmann and van Lohuizen, 2006), resulting in aberrant DNA methyltransferase silencing (Bracken and Helin, 2009). Cancers may be caused by the inactivation of tumor suppressor genes and/or reprogramming of the nucleus to pluripotent stages due to an imbalance in the dynamic chromatin configuration (Golbabapour et al., 2011; Mills, 2010). Alterations in the expression of the components of Pc complexes, along with modification of their marks, contribute to the development of a variety of cancers. For instance, during the development of cancers such as prostate cancer (Hoffmann et al, 2007; Saramaki et al., 2006), breast cancer (Yoo and Hennighausen, 2012), lymphoma (Visser et al., 2001), lung cancer (Breuer et al., 2004), and liver cancer (Cheng et al., 2011), expression of Ezh2 is increased compared with their normal counterparts. In fact, the higher the level of Ezh2 is, the poorer the cancer prognosis (for reviews, see Chang and Hung, 2012, and Simon and Lange, 2008).
Silencing Mechanisms of Polycomb Complexes
The silencing mechanism of the PcG proteins involves clonal transmission through molecular similarity between the PcG proteins and heterochromatin proteins (Paro, 1990). PcG proteins can bind to the promoter regions of genes and repress their functions (Fig. 3). However, a complete understanding of their functions in silencing genes remains to be deciphered.
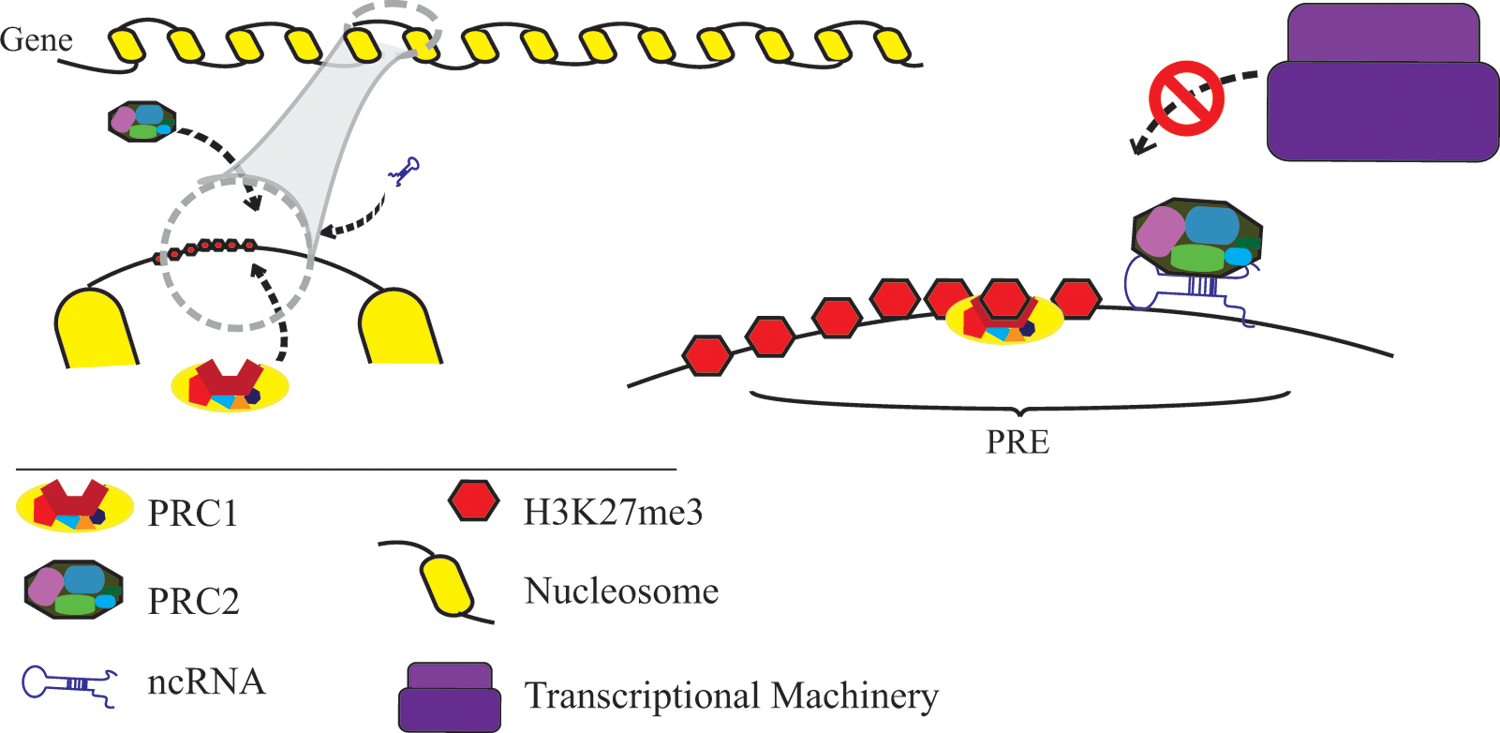
A schematic representation of recruitment of Pc complexes. Polycomb repression complexes (PRC1 and PRC2) recruit on Polycomb response elements (PRE), in association with ncRNA and other DNA-binding proteins (not drawn in the figure). PRC2 imposes high levels of H3K27me3 (as a repressive mark for chromodomains of PRC1).
In addition to other dynamics involved in their recruitment, Pc complexes bind to their targeted genes through their Pc domains and silence their expression (Bracken et al., 2006; Whitehead Institute for Biomedical Research, 2006). Several models have been proposed to describe the dynamic behavior of Pc complex binding (Beisel and Paro, 2011; Bracken and Helin, 2009; Hansen et al., 2008; Lanzuolo et al., 2011).
Packaging of DNA into chromatin structures is the main determinant of gene accessibility to the transcriptional machinery and is a regulatory mechanism of gene expression. Chromatin configuration regulates the expression of certain genes depending on cell type, which is necessary for cell differentiation and development (Golbabapour et al., 2011). In addition to the primary DNA sequence, epigenetic factors are fundamental in regulating gene expression. As such, chromatin remodeling, an epigenetic mechanism, has been considered to represent a crucial mechanism, particularly during development. This interaction between nucleic acids and proteins affects the accessibility of the DNA strand to the transcriptional machinery and, consequently, the regulation of gene expression. DNA methylation, particularly on CpG islands, defines targets for regulatory mechanisms (for a review, see Golbabapour et al., 2011). However, in different molecular physiological processes in cells, DNA methylation has been the subject of some controversy (Gilbert et al., 2007).
Chemical modifications of nucleosomal histones define the positions of nucleosomes (chromatin structure) and their accessibility to the transcriptional machinery. Different versions of PRC1 and PRC2 have been reported, and these diversities come with distinct functions (Vandamme et al., 2011). Genome-wide profiling has been widely exploited to understand PREs. A study on PRC1 and PRC2 revealed chromatin binding sites during PcG-mediated silencing (Tolhuis et al., 2006). PREs contain multiple Pc binding sites; however, no complete molecular profile of PREs has been described. This lack of understanding is due to the size of the complexes and the existence of various homologs and subunits (for a review, see Simon and Kingston, 2009). PREs contain motifs to which DNA binding proteins such as Pho (Klymenko et al., 2006), Zeste (Chen et al., 1992), and GAGA (Berger and Dubreucq, 2012) bind. Zinc-finger proteins, including Pho and Pho-like proteins, must bind to PRE to perform their biological functions (Brown et al., 2003; Fritsch et al., 1999; Mohd-Sarip et al., 2002). Upon binding to a PRE, Pho interacts with a posterior sex comb protein, SFMBT, and forms PhoRC (Klymenko et al., 2006) (for a review, see Schuettengruber and Cavalli, 2009). As mentioned previously, PcG proteins have numerous homologs and a variety of isoforms; consequently, these proteins engage in various binding patterns that, in turn, provide a diversity of complexes. Furthermore, Pc-like proteins can bind to PRC2 and alter its features and properties (O'Connell et al., 2001; Tie et al., 2003; Walker et al., 2010).
Studies addressing the PcG silencing mechanism have provided evidence for the dynamic control of gene expression (Kia et al., 2008; Kwong et al., 2008; Oktaba et al., 2008). Generally, long-term regulation of gene expression is controlled by Pc complexes and, antagonistically, by the trithorax group, through the modulation of chromatin structures and histone modifications. To exert their biochemical activities, PcG proteins must be assembled on PREs. Three main components are involved in the PcG-mediated silencing pathway: PcG proteins (Lund and van Lohuizen, 2004), DNA methylation systems (Bird, 2002), and Ezh2 (a PcG histone-lysine methyltransferase) (Vire et al., 2006). Ezh2 interacts with the other components and provides a recruitment platform for de novo methylation (Fuks et al., 2001). Generally, PRC2 inhibits transcription initiation, and PRC1 maintains the repressed status. PRC1 is able to ubiquitinate H2AK119 (Cao et al., 2005), and PRC2 can trimethylate H3K27 (Cao and Zhang, 2004a). Moreover, PRC2 exhibits catalytic activity that is involved in the methylation of H1K26 (Kuzmichev et al., 2004). Mediated by Ezh2, the addition of methyl groups to Lys27 of histone H3 by PRC2 results in silencing (Hansen et al., 2008) and is essential for the bioactivity of PRC1 (Cao et al., 2005). PcG complexes can also induce significant methylation of H3K27 (for a review, see Mueller and Verrijzer, 2009). The idea that PRC2 functions upstream of PRC1 is consistent with the fact that the Pc protein, a subcomplex, binds specifically to methylated H3K27; however, this premise remains to be proven (Margueron and Reinberg, 2011). PRC2 catalyzes the methylation of H3K27; the methylation of H3K27, particularly trimethylation, is the main hallmark of PcG-mediated silencing (Levine et al., 2004; Ringrose et al., 2003), which occurs through enzymatic activity (in association with PRC1). In addition to the nonenzymatic chromatin-compacting regulatory effect of PRC1 (Eskeland et al., 2010), this Pc complex is able to ubiquitylate histone H2A through Ring1A and Ring1B (for reviews, see Eckert et al., 2011; Richly et al., 2011; Vidal, 2009).
Furthermore, Ezh1 and Ezh2 are PRC2 subcomplexes that are able to methylate histone H3 by adding methyl groups to lysine 27 (Schuettengruber and Cavalli, 2009; Simon and Kingston, 2009). A recent review on the evolution of PRC2 discussed evidence that the primary silencing function of this multi-protein complex is accomplished through the methylation of H3K9. Specific cell lineages are needed to determine the specific functions of PRC2 (for a review, see Margueron and Reinberg, 2011). The maximum holoenzymatic activity of PRC2 is reportedly attributed to the additive contribution of each component of PRC2 (Margueron and Reinberg, 2011). Although PRC1 and PRC2 are two distinct complexes with different structures and functions (Bracken et al., 2006), they exhibit interdependence in regulating gene expression (Jorgensen et al., 2006). Methylation of H3K9 is associated with chromatin configuration and transcriptional regulation. Methylation of H3K9 is a repressive mark and a site for binding by HP1, which recognizes the methyl group and forms a protein dimer to perform its biological functions (Kwon and Workman, 2008), for example, the repression of imprinted genes (Monk et al., 2008). However, methylated H3K9 appears in the coding regions of some active genes (heterochromatin) during transcription elongation through mammalian chromatin (Vakoc et al., 2005). The specific function of H3K9 is dependent on its location in chromatin and its binding to different effector proteins (Kokura et al., 2010). The biochemical activities of the Pc complexes in Drosophila are exerted through the binding of these complexes genome-wide (Schwartz et al., 2006) and to PhoRC (Brown and Kassis, 2010; Klymenko et al., 2006). These complexes ubiquitylate histone H2A and suppress gene expression (Buchwald et al., 2006; Mueller and Verrijzer, 2009; Wang et al., 2004).
Another potential silencing mechanism is enabled by the ability of Pc complexes to bind to chromatin structures. PcG proteins are important in chromatin compaction (for a review, see Mueller and Verrijzer, 2009). To maintain their suppressive role during cell division, PcG proteins remain bound to chromatin and DNA during DNA replication (Francis et al., 2009). Bantignies and Cavalli (2011) argued that PcG proteins dynamically target chromatin in association with RNA polymerase II in a PcG-dependent manner. However, PRC1 has the ability to affect chromatin configuration independently of histone modification (Bantignies and Cavalli, 2011). In addition to post-transcriptional modifications (Berger, 2007), the dynamic structure of chromatin is mediated by two main features: chromatin remodeling and histone modifications such as acetylation, methylation, phosphorylation, ubiquitylation, deamination, ADP ribosylation, sumoylation, and proline isomerization (for a review, see Kouzarides, 2007). In addition to their interactions with DNA, chromatin modifications can affect each other. Histone modifications therefore provide spatially and temporally dependent profiles (known as the “histone code”) to mediate gene regulation and accessibility to the transcriptional machinery (Guil and Esteller, 2009; Turner, 2002). Certain types of DNA sequences, such as CpG islands, promoters, and repetitive elements, along with epigenetic factors, are the main features in determining chromatin configuration.
Recruitment of Polycomb Complexes
PcG proteins cannot specifically bind to DNA sequences. Studies on the associations of the transcriptional factors that recruit these proteins have revealed that multiple transcriptional factors are involved in specific binding to target genes (Bracken and Helin, 2009). During development in Drosophila, a number of transcription factors recruit PcG proteins to PREs (Ringrose and Paro, 2007), which are a combination of several binding sites (elements) (Bracken and Helin, 2009). Promoters in mammals mainly contain either low or high GC dinucleotides, which are classified as low- and high-CpG content promoters, respectively. These classes exhibit different patterns of histone modification and roles in regulation (Broad Institute of Harvard and MIT, 2007; Weber et al., 2007). CG-rich regions are important in PRC2 recruitment, and CpG islands are required for initial localization (Mendenhall et al., 2010) (for a review, see Margueron and Reinberg, 2011). PRC1 and PRC2 mostly target high-CpG content promoters (Boyer et al., 2006; Ku et al., 2008) (for a review, see Zhou et al., 2011). DNA-binding factors and CpG islands are two main features involved in the recruitment of Pc complexes to chromatin, particularly on PREs (Gal-Yam et al., 2008; Meissner et al., 2008; Mendenhall et al., 2010). Nucleosomal array analyses have shown that Pc components are able to remodel chromatin structure and compact chromatin independently of histone modifications (Eskeland et al., 2010; Francis et al., 2004; Margueron et al., 2008). PRC1 inhibits transcription at the promoter region of the targeted gene through chromatin remodeling (Lavigne et al., 2004), although this conclusion is controversial (Eskeland et al., 2010). A genome-wide study comparing active and inactive PcG-targeted regions revealed different levels of PRC1 recruitment. Coincidently, active regions lacking E(z) and Pc exhibit significant trimethylation at H3K9 and H3K27 (Breiling et al., 2004). Trimethylation usually occurs around the promoter regions of active genes, suppressing their expression (Bernstein et al., 2005; Kim et al., 2005). Studies of histone methylation in humans have demonstrated that monomethylation is a hallmark of transcriptional activation, while trimethylation is a hallmark of gene repression (Barski et al., 2007).
Polycomb Complexes and ncRNAs
ncRNAs are involved in the repressive function of silencing complexes and their interactions with nucleosomes. Many ncRNAs have been identified that interact directly with Pc complexes (Kanhere et al., 2010). For instance, HOTAIR and Xist RepA, two ncRNAs, interact with PRC2 to impose a silencing status on their respective genes (Rinn et al., 2007; Zhao et al., 2008). A microarray study of immunoprecipitated PcG proteins demonstrated that ncRNAs associate with PRC2 (Khalil et al., 2009). The SET domain is a sequence motif that catalyzes lysine methylation on histones. PcG proteins generally contain SET domains and Cbx, are mainly present in histone methyltransferases, such as Ezh2, and are involved in protein–genome interactions (Krajewski et al., 2005).
Studies have revealed a link between ncRNAs (which act as recruiters of PcG proteins) and Pc components such as PRC2 (Khalil et al., 2009). ncRNAs are important in the recruitment of PRC1 complexes and interact with Cbx (Yap et al., 2010). Critical in the recruitment of PcG proteins and targeting genes, long ncRNAs are able to recruit PRC1 and PRC2 complexes (for a review, see Bracken and Helin, 2009). Long ncRNAs (such as HOTAIR, KCNq1OT1, and REPA) recruit PcG proteins to chromatin (Bracken and Helin, 2009; Khalil et al., 2009; Zhao et al., 2008).
In addition to the transcriptional roles of long ncRNAs, the mechanism underlying their recruitment is based on their ability to bind specifically to the promoter regions of target genes (Fig. 3) and SET domains/Cbx (for a review, see Bracken and Helin, 2009). Long ncRNAs, such as HOTAIR (Rinn et al., 2007; Tsai et al., 2010), KCNq1OT1 (Kotake et al., 2011; Mohammad et al., 2010) and REPA (Zhao et al., 2008), mediate the methylation of H3K27 by PRC2. In humans, many long ncRNAs interact with PRC2 complexes and are implicated in regulatory functions in PcG-mediated silencing in trans (Khalil et al., 2009). Most ncRNAs exert their activity in cis (such as KCNq1OT1), although some act in trans (such as HOTAIR5). In a proposed model, long ncRNAs bind specifically to the promoters of their target genomic sequences and recruit Pc complexes (SET domains/Cbx) (Mercer et al., 2009; O'Meara and Simon, 2012; Yang et al., 2011).
ncRNAs mediate the relocation of target genes based on the interaction between nuclear subcompartments and nonhistone protein methylation to regulate gene expression (Yang et al., 2011). Yang et al. (2011) concluded that dimethylation of PRC2 on specific residues localized the targeted gene transcriptional machinery through NEAT2, an ncRNA related to interchromatin granules.
Argonaute proteins are approximately 100 kDa (Ender and Meister, 2010) and consist of two principal domains: a Piwi-Argonaute-Zwille domain and a Piwi domain (for reviews, see Ender and Meister, 2010; Parker and Barford, 2006 ). These proteins act as the molecular scaffolds in RNA silencing mechanisms and are necessary for the complementary binding of small ncRNAs to their targets (Parker and Barford, 2006). The silencing mechanism of Argonaute proteins is enabled by small ncRNAs that guide these proteins to their target sites (Takeda et al., 2008).
To exert their biological activity, particularly for guiding small RNAs in gene silencing activity, small ncRNAs are incorporated into Argonaute protein-containing complexes (Ender and Meister, 2010). These complexes contain specific domains that bind to small ncRNAs. Based on phylogenetic analyses, Argonaute proteins are classified into the Ago subfamily, which mostly associates with miRNAs and siRNAs, and the Piwi subfamily, which mainly associates with the germline and piRNAs (Ender and Meister, 2010). The biogenesis and germline functions of the Piwi subfamily are generally enabled by piRNAs (Girard et al., 2006; Houwing et al., 2007). Interactions between the Argonaute proteins and guide molecules are mainly mediated through interactions between the sugar–phosphate backbone of the nucleic acids and these proteins (Wang et al., 2008). The complementary levels of small ncRNAs and their RNA targets represent the main regulatory feature involved in Ago-mediated silencing mechanisms. Gene silencing by small ncRNAs that contain mismatches in their midregion occurs at the level of translation, particularly during its early steps (Ender and Meister, 2010). However, the expression levels and roles of the ncRNAs during development are different. Argonaute proteins exert their silencing role in the cytoplasm at the translational level and in the nucleus at the transcriptional level (for a review, see Guang et al., 2008). Despite recent progress in understanding and modeling the biological functions of ncRNAs in PcG-mediated silencing pathways (for a review, see Beisel and Paro, 2011), our understanding of epigenetics is still in its infancy, and further studies are required to thoroughly understand and grasp epigenetic silencing mechanisms.
Conclusions
Throughout the course of this review, we have discussed the structure and recruitment mechanisms of Pc complexes and their connection to ncRNAs. The phylogenetic distributions of PRC1, PRC2, and Cbx in Drosophila, mouse, rat, and human were also briefly presented. PcG proteins and their regulatory roles in epigenetics, particularly during development, were discussed. Multiple epigenetic pathways govern cell fates and developmental states. Thus, homeostasis, or cellular balance, is the outcome of epigenetic regulation with proper timing. Cancer generally results from an imbalance among epigenetic mechanisms, such as those mediated by PcG proteins and trithorax group proteins that maintain homeostasis in the activity of certain genes. For instance, improper reactivation of pluripotent genes such as the Hox genes initiates carcinogenesis (for a review, see Ringrose and Paro, 2004). In Drosophila, Hox genes are active during early embryogenesis and maintained during adult life by epigenetic mechanisms.
In the last decade, several researchers have focused on understanding the epigenetic mechanisms involved in modifying chromatin. Among these factors, Pc complexes impact the structural modification of chromatin and maintain the silenced state with respect to the expression of certain genes. In fact, PcG proteins cooperate with specific domains of chromatin to silence the expression of genes. Polycomb proteins form at least two distinct complexes, PRC1 and PRC2. The components of Pc complexes, which are known to regulate homeotic genes, vary in different stages of development to control hundreds of other genes in mammals and insects. These complexes are also important in a variety of different cancers. The dynamic nature of Pc complexes increases their flexibility and permits their accurate interaction with environmental signals to regulate the expression of genes as global epigenetic repressors. In cancerous diseases, epigenetic studies have shown that these complexes have roles in transcriptional misregulation, particularly of tumor suppressor genes, leading to unscheduled activation or repression of undesired pathways and thereby enhancing cancerous proliferation. Pc complexes govern the methylation marks on histone H3 through their chromodomains, establishing the chromatin configuration and the spatial distribution of genes within the nucleus. Consequently, Pc complexes control the level of expression of a gene. Numerous studies have emphasized the importance of ncRNAs in epigenetic regulation mediated by PcG proteins. However, our knowledge of Pc complexes and their roles and mechanisms does not provide insight into epigenetic networks, nuclear reprogramming or diseases.
Footnotes
Acknowledgments
Preparation of this review was supported by University of Malaya Grants: UM/MOHE High Impact Research Grant No. F000009-21001, PG016-2012B and FP041/2010A.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.