
Abstract
Abstract
Phosphorylation is a major regulatory mechanism in eukaryotic cells performed by the concerted actions of kinases and phosphatases (PPs). Protein phosphorylation has long been relevant to sperm physiology, from acquisition of motility in the epididymis to capacitation in the female reproductive tract. While the precise kinases involved in the regulation of sperm phosphorylation have been studied for decades, the PPs have only recently received research interest. Tyrosine phosphorylation was first implicated in the regulation of several sperm-related functions, from capacitation to oocyte binding. Only afterwards, in 1996, the inhibition of the serine/threonine-PP phosphoprotein phosphatase 1 (PPP1) by okadaic acid and calyculin-A was shown to initiate motility in caput epididymal sperm. Today, the current mechanisms of sperm motility acquisition based on PPP1 and its regulators are still far from being fully understood. PPP1CC2, specifically expressed in mammalian sperm, has been considered to be the only sperm-specific serine/threonine-PP, while other PPP1 isoforms were thought to be absent from sperm. This article examines the “Omics” of human sperm, and reports, for the first time, the identification of three new serine/threonine-protein PPs, PPP1CB, PPP4C, and PPP6C, in human sperm, together with two tyrosine-PPs, MKP1 and PTP1C. We specifically localized in sperm PPP1CB and PPP1CC2 from the PPP1 subfamily, and PPP2CA, PPP4C, and PPP6C from the PPP2 subfamily of the serine/threonine-PPs. A semi-quantitative analysis was performed to determine the various PPs' differential expression in sperm head and tail. These findings contribute to a comprehensive understanding of human sperm PPs, and warrant further research for their clinical and therapeutic significance.
Introduction
In spermatozoa, increased protein-tyrosine phosphorylation has been associated with capacitation (Petrunkina et al., 2001), hyperactivated motility (Nassar et al., 1999), zona pellucida interaction (Burks et al., 1995), acrosome reaction and sperm-oocyte binding and fusion (Urner and Sakkas, 2003). Different tyrosine-phosphorylated proteins have been identified in sperm from diverse species, localized to different subcellular regions of the spermatozoa, and their phosphorylation status linked to a specific sperm function (Urner and Sakkas, 2003).
Concerning protein-serine/threonine phosphorylation, PKA, the cAMP-dependent kinase, plays a pivotal role in sperm motility, capacitation and the acrosome reaction (Bielfeld et al., 1994; Skalhegg et al., 2002; Visconti et al., 1995). PKA, together with PPs, is anchored to AKAPs (A Kinases Anchored Proteins) allowing for a tight regulation of the reversible phosphorylation mechanisms performed by the anchored PKA and PPs (Fardilha et al., 2011a). Noteworthy, the AKAPs are themselves regulated by phosphorylation (Carrera et al., 1996; Johnson et al., 1997; Mandal et al., 1999). Furthermore, members of the MAPK pathway (mitogen-activated protein kinases, or ERK, extracellular signal-regulated kinases) have been identified in spermatozoa as playing a role in capacitation (de Lamirande and Gagnon, 2002) and in the acrosome reaction (de Lamirande and Gagnon, 2002; du Plessis et al., 2001; Luconi et al., 1998).
Much less is known about the role of serine/threonine-PPs (STPPs) in spermatozoa (Fardilha et al., 2011a; Signorelli et al., 2012). STPPs are divided into three distinct gene families, the PPM (Mg2+ or Mn2+-dependent protein phosphatases), the FCP (TFIIF-associating C-terminal domain phosphatases), and the PPP (phosphoprotein phosphatases). The PPM family comprises pyruvate dehydrogenase, PP2C, and relatives (Barford et al., 1998). The FCP family comprises CTD phosphatases (FCP1) and small CTD phosphatases (SCPs) (Gallego and Virshup, 2005; Yeo et al., 2003). The PPP family includes PP1/PPP1, PP2/PPP2 (PP2A/PPP2CA, PP4/PPP4C, and PP6/PPP6C), PP2B/calcineurin/PPP3C, PP5/PPP5C, and PP7/PPP7C gene subfamilies, which share high homology in the catalytic domains but differ in their N- and C-terminal domains (Barford et al., 1998; Berndt et al., 1987; Cohen, 1997; da Cruz e Silva and Cohen, 1987; da Cruz e Silva et al., 1987; da Cruz e Silva et al., 1988; Honkanen and Golden, 2002). Of the three mammalian phosphoprotein phosphatase 1 (PPP1) genes, PPP1A/α, PPP1B/β, and PPP1C/γ, the latter is known to undergo alternative splicing to yield the ubiquitous PPP1CC1/γ1 and the testis/sperm-enriched PPP1CC2/γ2 isoforms. Previous work has shown that PPP1CC2 is specifically expressed in mammalian sperm and likely to play a key role in the control of sperm motility and morphogenesis (Chakrabarti et al., 2007a; Huang et al., 2002; Mishra et al., 2003; Shima et al., 1993; Smith et al., 1996; Smith et al., 1999; Varmuza et al., 1999; Vijayaraghavan et al., 1996). In sperm, PPP1CC2 is present along the entire flagellum, including the middle-piece, consistent with a role in sperm motility, but it is also found in the posterior and equatorial regions of the head, indicating a role in the acrosome reaction (Huang et al., 2002). Immotile spermatozoa have higher activity levels of PPP1CC2 compared to motile spermatozoa. Inhibition of PP activity by okadaic acid and calyculin A initiates motility in caput epididymal sperm without requirement for a change in cAMP levels (Smith et al., 1996; Vijayaraghavan et al., 1996). PPP1CA and PPP2CA have been shown to be expressed in hamster and human sperm (Huang et al., 2004; Vijayaraghavan et al., 1996). PPP3C, calcineurin, the calcium/calmodulin-dependent phosphatase, has been detected in hamster (Suzuki et al., 2010) and dog spermatozoa (Tash and Means, 1988) and seems to dephosphorylate tyrosine phosphorylated substrates in human sperm (Carrera et al., 1996).
Cellular health and vitality are dependent on the fine tuning of the protein phosphorylation components. Many disorders are associated with the abnormal phosphorylation of key proteins (e.g., cancer, diabetes, and neurodegenerative conditions) (Fardilha et al., 2010). Alterations in any factor that interferes with sperm motility may also lead to infertility. In fact, asthenozoospermia is one of the main male pathologies causing infertility (Curi et al., 2003). Indeed, a recent proteomic approach revealed differential protein phosphorylation levels between high and low motility sperm samples (Chan et al., 2009).
There are several studies using mouse models that demonstrate a clear association between a gene and sperm motility alterations or sperm morphology defects. PPP1C gene null male mice are infertile due to impaired spermatogenesis, leading to the absence of epididymal spermatozoa (Varmuza et al., 1999). Although PPP1CA expression was increased and its localization altered, it could not substitute for PPP1CC, which further indicates a specific role for PPP1CC2 in sperm differentiation and morphogenesis (Chakrabarti et al., 2007b).
Even though PPP1CC2 has been linked to sperm motility for about 2 decades, almost no data are available for the other STPPs. Here, the existence of several PPs in human sperm was established. Moreover, as alterations in PPs activity and/or expression levels are important to the pathology of male infertility, a comparison between the profiles of the different PPs in motile human sperm versus asthenozoospermic sperm was undertaken. Our findings permit the identification of the PPs present in human sperm and the subcellular localization of some STPPs in both normo and asthenozoospermic samples.
Materials and Methods
Biological material
Ejaculated human sperm samples were obtained from healthy donors (kindly provided by Clínica Dr. Alberto Barros, Porto, and Ferticentro, Coimbra) undergoing routine semen analysis. All donors signed informed consents, and all human samples were used in accordance with the appropriate ethical and Internal Review Board guidelines.
Sperm analyses
Semen analyses were performed according to the World Health Organization (WHO) guidelines. Sperm motility was evaluated by classification into three groups of motility, as recommended by WHO (WorldHealthOrganization, 1999). Briefly, this parameter was evaluated by using a wet preparation with an aliquot of sperm cells deposited onto a slide and further scoring of spermatozoa that were progressive (PM), nonprogressive (or in situ) (NPM), and immotile (IM). The concentrations of sperm samples were determined (WorldHealthOrganization, 1999), by counting the number of spermatozoa (spz) using an improved Nuebauer chamber,® expressed as ×106 spz/mL. Characterization of the samples used (n=11) is described in Table 1. Sperm samples were washed three times with 1× PBS, and sperm extracts were prepared as described below for each method used.
AZ, asthenozoospermic; NZ, normozoospermic; spz, spermatozoa.
Multi-immunoblotting analyses
The Kinetworks™ Protein Phosphatase Screen 1.2 (KPPS) provided semi-quantitative information on the expression levels of 29 phosphatases in total protein extracts (Appendix 1). The screen was performed by Kinexus Bioinformatics Corporation (Vancouver, BC, Canada), using sperm samples with different motility parameters
Preparation of sperm extracts
For the preparation of sperm extracts used in the Western blots, supernatant and pellet extract fractions were prepared, using homogenization buffer (10 mM Tris-HCl, pH 7.0, 1 mM EDTA, 1 mM EGTA) with protease inhibitors (10 mM benzamidine, 1 mM PMSF, and 0.1 mM TPCK). After centrifugation at 16,000 g for 20 min at 4°C, the supernatant fraction (soluble fraction) was collected to a new tube and the pellet was dissolved in 1% SDS (insoluble fraction). Sperm extracts were normalized to 5×105 or 2×105 spz/mL, boiled, and then sonicated for 15 sec.
Antibodies
The primary antibodies used in this work were the STPPs antibodies and the control antibodies. The STPPs antibodies included rabbit polyclonal CBC502 against the C-terminal of PPP1CC2 (da Cruz e Silva et al., 1995; Fardilha et al., 2011b), rabbit polyclonal anti-PPP1CB (Upstate Biotechnology Inc. #07-270, Lake Placid, NY), rabbit polyclonal anti-PPP1CA (da Cruz e Silva et al., 1995), rabbit polyclonal anti-PPP4C (Abcam #ab16475, Cambridge Science Park, UK), rabbit polyclonal anti-PPP6C (Sigma #P8734, St. Louis, MO), and mouse monoclonal anti-PPP2CA (Upstate Biotechnology Inc. #05-421). The epitopes to which the antibodies were raised are indicated in Appendix 2. Mouse monoclonal α-tubulin acetylated (Life Technologies S.A., Madrid, Spain) and mouse monoclonal β-tubulin (Life Technologies S.A.) were used as controls.
The secondary antibodies used were: horseradish peroxidase-conjugated anti-rabbit and anti-mouse IgGs (GE Healthcare, Amersham Biosciences Europe GmbH, Freiburg, Germany). Secondary fluorescent labeled antibodies anti-rabbit Texas-Red (Molecular Probes, Eugene, OR, USA), anti-mouse Texas Red (Molecular Probes), and anti-mouse Alexa Fluor 488 (Invitrogen, Eugene, OR, USA) were also used.
Western blot analysis
To follow up on the KPPS 1.2 screen results, the expression levels of identified STPPs were analyzed by SDS-PAGE, followed by Western blotting with the appropriate antibodies (Appendix 2), using human sperm samples with different characteristics (Table 1). Sperm extracts prepared as explained above were resolved by 10% SDS-PAGE and proteins were subsequently electrotransfered onto nitrocellulose membranes and immunodetected with the appropriate specific antibodies: polyclonal rabbit CBC502 (1:2000); rabbit polyclonal anti-PPP1CB/β (1:500); rabbit polyclonal anti-PPP1CA/α (1:1000); rabbit polyclonal anti-PPP4C (1:1000); rabbit polyclonal anti-PPP6C (1:1000); mouse monoclonal anti-PPP2CA (1:2000). Immunoreactive bands were detected by enhanced chemiluminescence (ECL®) using the secondary antibodies horseradish peroxidase-conjugated anti-rabbit (1:5000), or anti-mouse (1:5000) IgGs.
The control antibodies used included mouse monoclonal α-tubulin acetylated (1:2000) and mouse monoclonal β-tubulin (1:500). The latter was used after membrane striping. Briefly, the membrane was incubated for 30 min with stripping solution (2% SDS, 62.5 mM Tris-HCl, pH 6.7, 100 mM β-mercaptoethanol) at 50°C and 70 rpm, washed with TBST for 15 min with agitation, twice with water, and then left to dry. Immunoreactive protein bands were quantified using the Quantity One densitometry software (Bio-Rad, Portugal).
Immunocytochemistry
To validate the subcellular localization of the STPPs in human sperm, an aliquot of washed diluted sperm (25 μL) was placed onto a glass coverslip pre-coated with 100 μg/mL poly-L-ornithine, and dried at room temperature, in a 6-well plate containing one coverslip per well. For fixation, 1 mL of 4% paraformaldehyde in 1xPBS was gently added to each well and left to stand for 10 min. Subsequently, sperm were washed twice with 1 mL of 1xPBS for 10 min. For permeabilization, 1 mL of 1:1 methanol/acetone solution was added for 2 min, and then the specimens were washed twice with 1 mL of 1xPBS for 10 min, and blocked for 1 h with 3% BSA in 1xPBS, before incubation with primary antibodies for 2 h at room temperature. The STPPs primary antibodies (and dilutions) used in this study (see Antibodies section and Appendix 2) included rabbit anti-PPP1CC2 (1:1000), rabbit anti-PPP1CB (1:500), rabbit polyclonal anti-PPP4C (1:500), rabbit polyclonal anti-PPP6C (1:500), mouse monoclonal anti-PPP2CA (1:500), and mouse monoclonal α-tubulin acetylated (1:250). After three washes with 1xPBS, the fluorescent-labeled secondary antibodies were added and the coverslips incubated for 2 h. The secondary fluorescent-labelled antibodies used were: anti-rabbit Texas-Red (1:300), anti-mouse Texas Red (1:300), and anti-mouse Alexa Fluor 488 (1:50). Finally, three washes with 1xPBS were performed and coverslips were mounted on microscope glass slides with one drop of anti-fading reagent containing DAPI for nucleic acid staining (Vectashield, Vector Laboratories Burlingame, CA, USA). Fluorescence images were acquired by confocal microscopy in a LSM 510 META confocal microscope (Zeiss) using an Argon laser line of 488 nm (green channel), a 561 nm DPSS laser (red channel), and a Diode 405–430 nm laser (blue channel). Profiles of single sperm cells were acquired using the Zeiss LSM 510 4.0 software as previously described (Vieira et al., 2010). Semi-quantitative assessment of phosphatases nuclear co-localization (n=30) was performed with the Just Another Colocalization Plugin (JACoP) from NIH Image J software using the Mander's M1 and M2 coefficients (Dunn et al., 2011). Manders' coefficients indicate the level of co-localization between red and blue channels (and vice versa) and are independent of the intensity of fluorescent signals. The values of Manders' coefficients range from 0 to 1, with 1 indicating complete co-localization and 0 indicating no co-localization.
Results
Sperm phosphatases high throughput screen
The antibody-based KPPS (Kinetworks™ Protein Phosphatase Screen 1.2) was initially employed to investigate the presence of PPs in pooled human sperm (NZ1, Table 1). Following this optimization protocol, a motile sample (NZ2, Table 1), that had spermatozoa with an intermediate motility, was compared to an immotile sperm sample (AZ1, Table 1). Screening of several PPs, which are signaling molecules that normally exist in low abundance, required the use of multi-immunoblotting techniques, such as the KPPS method. The affinities of individual antibodies used to screen the different PPs varied; therefore, this screening approach does not provide an absolute comparison of different protein levels but indicates the relative amounts of a target protein in two different samples. The KPPS screen permits detection of 29 PPs (Appendix 1), which include tyrosine-PPs (8), STPPs (13), threonine/tyrosine-PPs (7), and serine/tyrosine-PPs (1). Of these, only 14 were detected in the sperm pool (NZ1), 10 in the NZ2, and 7 PPs in the AZ1 sample (Table 2). The remainder of the PPs screened were undetectable, either due to their absence in the sperm samples tested or because their levels were below the detection limit.
AZ, asthenozoospermic; cpm, counts per minute; CS, catalytic subunit; NZ, normozoospermic; RS, regulatory subunit.
The levels of chemiluminescent signals for immunoreactive target phosphatases from normozoospermic (NZ) versus asthenozoospermic (AZ) sperm samples are expressed as counts per minute (cpm).
From the 16 PPs screened for dephosphorylate tyrosine residues, only four were detected: PTP1D, PTP1C, MKP1, and MKP2. The first two are tyrosine-PPs and the last two are dual specificity threonine/tyrosine-PPs. All the tyrosine-PPs were detected in the NZ1 sample except for MKP1 (Table 2), which was present in NZ2 and AZ1. For all the other PPs (i.e., the STPPs), they were always evident in the NZ1 sample. For comparison purposes, the differences between NZ2 and AZ1 were investigated. PTP1D, as well as two isoforms of the STPP, PPP6CB and PPP6CC, were only detected in NZ2 sperm sample (they are shown in Table 2 as an increment). The remaining five PPs were mainly expressed in NZ2 sperm sample: the catalytic subunit of PPP1CA, PPP1CB, PPP2CA, PPP4C, and PPP5C. However, the PPP2 regulatory subunits, PPP2R1A/B, were highly expressed in the AZ1 sample.
The PPP1CA and PPP1CB isoforms were, for the first time, identified in human mature sperm, as were PPP4C, PPP5C, PPP6CB, and PPP6CC. The PPP1CC2 isoform has been previously identified in mammalian sperm (Smith et al., 1996) and is considered to be the sperm-enriched PP (Fardilha et al., 2011b). Nevertheless, the KPPS-1.2 analysis failed to detect this isoform in both motile and immotile human sperm. As the PPP1 antibodies for the various PPP1 isoforms did not detected the presence of PPP1CC2 in the human motile or immotile sperm samples tested by this PP screen, this testis/sperm-enriched isoform was further searched in human sperm samples by Western blotting and immunocytochemistry (see below). Likewise, the other most sperm abundant class of PPs based on the KPPS-1.2 screen, the STPPs, were also further investigated and confirmed by these follow up studies.
Sperm phosphatases validation results
In this study, validation of the KPPS-1.2 screen results was focused on the serine/threonine PPs in human sperm samples, since they were the most abundant class of PPs detected by this method. For comparison purposes, the presence and the relative levels in sperm of the different PPs, PPP1 (PPP1CA, 37 kDa, PPP1CB, 38 kDa, and PPP1CC2, 39kDa) and PPP2 (PPP2CA, 36 kDa, PPP4C, 34 kDa, and PPP6C, 32 kDa) subfamilies, were examined in normozoospermic sperm sample soluble (left panel) and insoluble (right panel) fractions (Table 1 and Fig. 1; NZ3). β-Tubulin (present in the microtubules of the flagella axoneme) and α-tubulin acetylated (used as a marker for microtubules stability) were used as internal controls. In both fractions PPP2CA was the most abundant PP. In the soluble, PPP1CC2 was also detected. In the insoluble fraction, as expected, all the PPs were detected, except PPP1CA (Fig. 1), which indicates that the majority of the STPPs studied are tightly attached to the axoneme. After showing that the vast majority of the STPPs are present in the insoluble sperm fraction, we proceeded to confirm that STPPs were also expressed in asthenozoospermic human sperm samples insoluble fraction. Experiments were performed with two asthenozoospermic samples, AZ2 (data not shown) and AZ3 (Fig. 2). All the PPs analyzed from the PPP2 subfamily (PPP2AC, PPP4C, and PPP6C) are expressed, as well as PPP1CB and PPP1CC2, from the PPP1 subfamily. The absence of PPP1CA was confirmed, although it was evident with the KPSS-1.2 screen. This discrepancy may be due to the different sensitivity of the antibodies used or to variations amongst individuals, which could also explain the apparent absence of PPP1CC2 with the KPSS-1.2 screen.
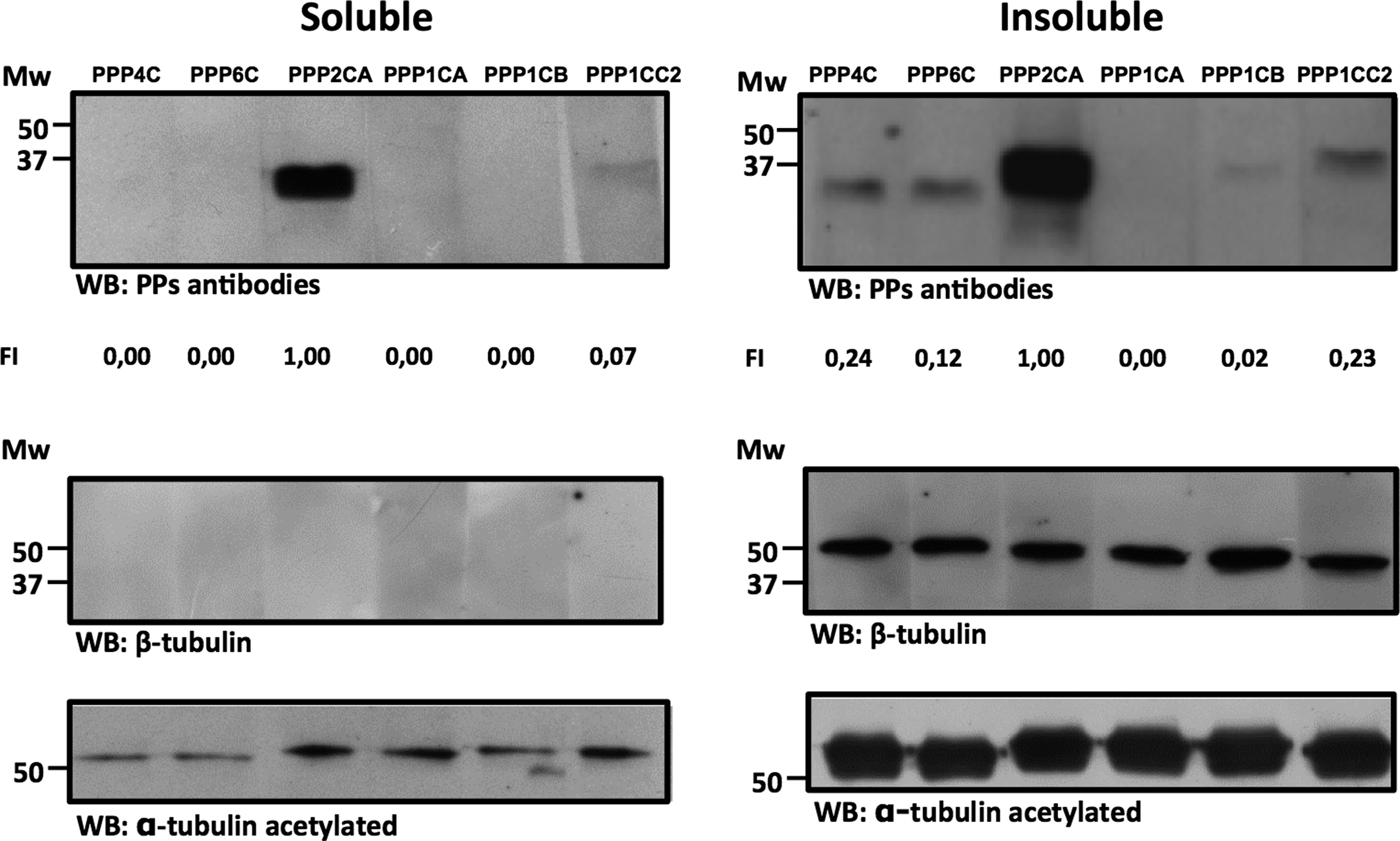
Expression of STPPs in normozoospermic human semen samples. Semen samples were separated into soluble and insoluble fractions, and the same concentration of spermatozoa, corresponding to 5×105 spz/mL, were loaded per lane. β-Tubulin and α-tubulin acetylated were used as internal controls. Normalization of the expression values was performed as the ratio to β-tubulin and the fold increase (FI) relatively to PPP2CA is presented (NZ3). The molecular mass is in kDa.
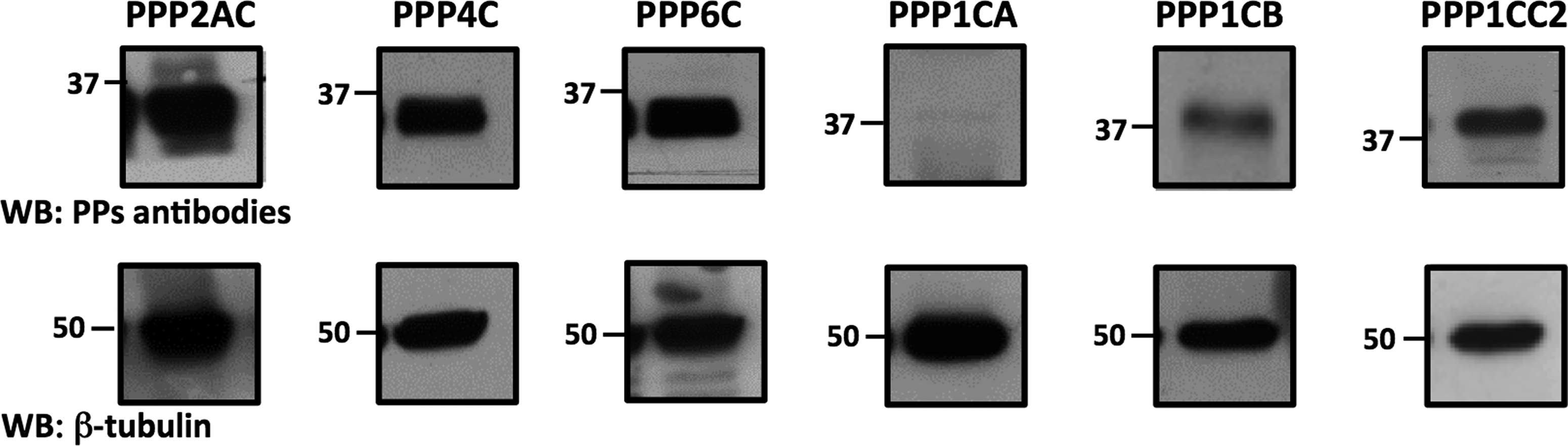
Expression of STPPs in the insoluble fraction (see Methods section) of asthenozoospermic human semen sample (AZ3). Equal loading based on the concentration of spermatozoa (2×105 spz/mL) was performed. β-Tubulin was used as an internal control. Numbers (37 and 50) correspond to the molecular mass markers (kDa).
The presence of STPPs in human spermatozoa was also addressed by immunocytochemistry (the specificity of the antibodies used was previously shown, see Methods section). Images representative of the results obtained are shown in Figure 3 and 4. All of the PPPs analyzed were present, either in the head or in the tail of the spermatozoa, both in NZ and AZ human samples. For all the STPPs tested, a co-localization with α-tubulin acetylated was performed except for PPP2CA, since the antibodies were both raised in mice (Fig. 4). For PPP1CA, results were not obtained, as the antibody was not suitable for immunocytochemistry. DAPI (blue) stains the nucleus in the sperm head. The tail is clearly delimited by the α-tubulin acetylated staining. There was an overall decrease in the intensity of the PPP2 subfamily expression from NZ to AZ sperm samples. This finding was in accordance with the KPPS-1.2 screen results. A profile for PPP4C was made (Fig. 4) and this confirmed that PPP4C expression intensity along a predefined axis decreased from NZ to AZ samples. To further quantify the results, a semi-quantitative co-localization analysis was performed with the JACoP plugin, using the Mander's M1 and M2 coefficients, to access the relative distribution of each PP (pixels) independently in the sperm head and tail (Table 3). In general, from NZ to AZ, PPs relative distribution increased in the head. PPP1CB and PPP2CA have a similar distribution in head and tail in NZ sperm, although PPP2CA tended to accumulate in the head (p=0.015) of AZ sperm. PPP1CC2, PPP4C, and PPP6C were highly abundant in sperm tail in NZ sperm. Nevertheless, the relative distribution in the head for the three PPP isoforms increased in asthenozoospermia (Table 3). Apparently, for PPP4C and PPP6C, the relative increase in the sperm head was due to a decrease in the middle-piece (Figs. 3 and 4). The increase was statistically significant only for PPP4C and PPP6C (p<0.001).

Immunolocalization of PPP1CB and PPP1CC2 in normozoospermic (NZ) and asthenozoospermic (AZ) human semen samples.
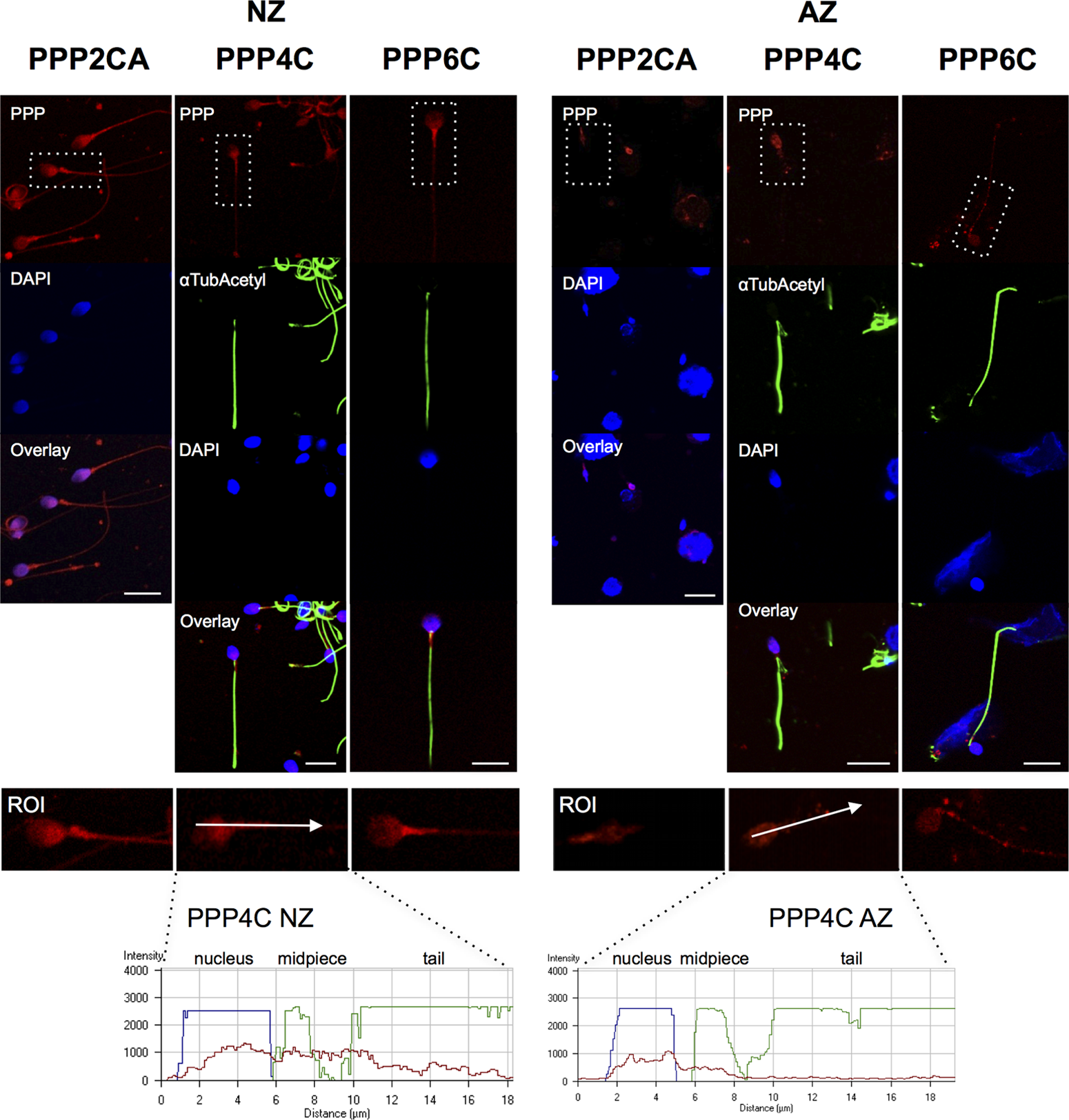
Immunolocalization of PPP2CA, PPP4C, and PPP6C in normozoospermic (NZ) and asthenozoospermic (AZ) human semen samples. Representative images are shown from NZ samples (NZ3 and NZ4) and AZ samples (AZ2 and AZ3). α-Tubulin acetylated was used as an internal control, except with the anti-PPP2A antibody, also of mouse origin. Nucleus was stained with DAPI. The fluorescence intensity profiles of PPP4C represent the voxels through the white arrowed lines indicated in the ROI image.
Gray boxes indicate PPs that are here described for the first time as being present in human sperm. SEM, standard error of the mean; STDV, standard deviation.
Discussion
Many reports use mass spectrometry based techniques to identify nonspecific proteins from human sperm (Gilany et al., 2011). Particularly, a recent work from Amaral et al. (2013) detected in NZ human sperm tail some PP. Here, a broad antibody-based PP screen was used to search specifically for PPs in human sperm, which permitted the identification of a human sperm partial phosphatome. The identified PPs might be responsible for the dephosphorylation of diverse protein substrates in sperm cells. Further, an in-depth characterization was made with the STPPs identified due to their participation in the acquisition of the epididymal motility and also to the fact of being highly abundant in the initial screening. Thus, several STPPs were further validated by Western blotting and immunocytochemistry, in particular the members from the PPP2 (PPP4, PPP6, and PPP2AC) and PPP1 (PPP1CB and PPP1CC2) subfamilies.
Beside the STPPs identified, four tyrosine-PPs were detected with the KPPS-1.2 screen: PTP1D, PTP1C, MKP1 (or DUSP1), and MKP2 (or DUSP4). Recently, Gonzalez-Fernandez et al. (2009) performed a systematic immunoblotting screen in mammalian sperm lysates (pig, horse, and dog), which was designed to identify 24 classical tyrosine-PPs, and discovered only four in boar and stallion sperm (i.e., PTPRB, PTP1D, DUSP3, and DUSP4) and three in dog (i.e., PTP1D, DUSP3, and DUSP4) (Gonzalez-Fernandez et al., 2009). In the same study, they showed that PTP1D was mainly located to the post-acrosomal region of the head and in the tail middle-piece. Furthermore, treatment with tyrosine-PPs inhibitors (e.g., pervanadate) reduced motility in a concentration-dependent manner. Sperm motility could be recovered after inhibitor removal, reinforcing the idea that one or more vanadate-sensitive tyrosine-PPs play a critical function in the regulation of sperm motility.
By contrast, inhibitors of STPPs have an opposite behaviour with respect to sperm motility. Treatment of NZ sperm with tyrosine-PPs inhibitors reduced motility while treating AZ sperm with STPPs inhibitors induced motility (Vijayaraghavan et al., 1996). Thus, to have a functional sperm cell, STPPs such as PPP1CC2, need to be inhibited, while tyrosine-PPs have to be active. This is in agreement with the result of the KPSS-1.2 screen for PTP1D, where the tyrosine-PPs were expressed only in the NZ sample.
MKP1 and PTP1C, to our knowledge, are described here for the first time as being present in human sperm, although previously their presence was tested in pig, horse, and dog sperm (Gonzalez-Fernandez et al., 2009). MKP1 is one of several dual-specificity PPs that dephosphorylate the regulatory threonine and tyrosine residues of its target MAPK, inhibiting immune responses. The expression of MKP1 is induced by a number of growth factors and stress factors and conditions in multiple cell types. For example, in macrophages, the induction of MKP1 correlates with decline in the activities of JNK and p38MAPK (which we also observed in human sperm, data not shown). This is consistent with a role of MKP1 in inhibiting these MAPKs as a feedback mechanism to control inflammation (Chi and Flavell, 2008). PTP1C is a tyrosine-PP predominantly expressed in cells of hematopoietic origin. Mutations in this PP have been associated with multiple hematopoietic defects, which indicates that tyrosine phosphorylation is crucial to modulation of cell signalling pathways in proliferation and differentiation (Bignon and Siminovitch, 1994). The precise role of PTP1C in sperm still needs to be elucidated. Here, PTP1B was not detected, in contrast with a previous study that showed the presence of this PP in human sperm (Tomes et al., 2004). This apparent contradictory result may reflect the fact that the protein in our samples may have been below the detection limit of the antibody used in the KPPS-1.2 screen.
More recently, STPPs have caught researchers' attention for their involvement in signaling pathways as diverse as sperm motility, sperm morphogenesis, and capacitation (Adachi et al., 2008; Chakrabarti et al., 2007a; Fardilha et al., 2011a; Vijayaraghavan et al., 1996). STPPs functions in sperm have mainly been attributed indiscriminately to PPP2CA or PPP1C, without any isoform specificity. Here, we used isoform-specific antibodies to characterize the expression of PPP1 and PPP2 subfamily members in human sperm and their subcellular localization. The specific PP isoforms should have particular functions in the different spermatozoa compartments by associating with different partners (Fardilha et al., 2011a). We confirmed the presence of PPP2CA, PPP1CC2, and PPP1CA and showed, for the first time, the presence of PPP4C, PPP6C and PPP1CB (Figs. 1 and 2 and Table 2). All PPP2 members are present in all spermatozoa compartments, although at different intensities (Figs. 3A). From the PPP1 subfamily members, PPP1CC2 is the highly enriched member (Figs. 3B), prevalent in sperm tail as previously reported (Fardilha et al., 2011b).
Several research groups have addressed the differences between NZ and AZ men, according to the WHO criteria (1999) (Martinez-Heredia et al., 2008; WorldHealthOrganization, 1999; Zhao et al., 2007). Essentially, differences have been seen in proteins related to energy metabolism, structure, and movement. Here, we first looked for differences in the levels of PPs expression by Western blotting (Figs. 1 and 2) and validated the KPPS-1.2 screen to confirm the presence of several PPs in the sperm samples. With subsequent immunocytochemistry studies using confocal microscopy, we were able to detect the expression of the PPs in a single sperm cell (Figs. 3 and 4) and their subcellular localizations. Comparisons between NZ and AZ samples were also performed. Expression levels of all PPP2 subfamily members decreased when comparing NZ versus AZ samples (Table 2). Differences in expression do not necessarily translate directly to differences in enzymatic activity. Thus, it would be of extreme importance to determine PPP2 phosphatase activity in a single sperm cell model. To better quantify the subcellular localization of the different PPs, a semi-quantitative analysis was performed and the results revealed that all the PPs tested were present in both the tail and the head of the sperm cells. Their nuclear localization changed when comparing samples from NZ to AZ in that PPs seem to be relatively more abundant (more number of pixels) in the head, and this was particularly evident for PPP1CC2, PPP4C and PPP6C. Moreover, the relative distribution was statistically different between NZ and AZ sperm samples only for the PPP2 subfamily members. Thus, PPs from the PPP2 subfamily may also have a role in asthenozoospermia in the regulation of sperm motility events.
Conclusion
In this study we report, for the first time, the identification in human sperm of three new STPPs—PPP1CB, PPP4C, and PPP6C—a tyrosine-PP–PTP1C and a dual specificity-PP–MKP1. Moreover, we reconfirm the existence of the so-called sperm-specific PP, PPP1CC2, and of PPP1CA. Also, the relative distribution of the STPPs PPP1 and PPP2 subfamilies members was addressed in NZ and AZ sperm samples. Overall, the relative distribution of the PPs increased in the sperm head, in comparison to the tail, from NZ to AZ sperm samples. Evaluation of protein expression levels and subcellular localization is proving to be a useful tool to validate molecular biomarkers for diagnostic and prognostic purposes in male infertility. Unravelling the relevance of (de)phosphorylation events may contribute to the development of novel male contraceptives (Fardilha et al., 2011a). It is clear from the results presented that different samples (NZ and AZ) have different STPPs relative distribution profiles. Results such as those presented here may permit the development of a platform based on PPs expression, activities, or both, that could serve for diagnostic and prognostic purposes, in addition to basic semen analysis. Such a PP platform, in the near future, would be useful and extremely practical in a clinical setting by providing valuable information with respect to the molecular basis of male infertility.
Footnotes
Acknowledgments
This work was supported by the Center for Cell Biology of the University of Aveiro, and by grants from FCT of the Portuguese Ministry of Science and Higher Education to MFe (SFRH/BD/73000/2010) and LKG (SFRH/BD/42334/2007). We thank Dr. Rui Miguel Conde and Dr. Carl Smith (University of Sheffield) for providing PPP4C, PPP6C, and PPP2CA antibodies. This article is dedicated to the memory of Edgar da Cruz e Silva who was a visionary scientist and was never afraid of thinking “outside the box”. His scientific enthusiasm and curiosity will be forever remembered.
Author Disclosure Statement
Dr. Steven Pelech and his family are majority owners of Kinexus Bioinformatics Corporation. No other competing interests exist.
Abbreviations Used
| Abbreviation | Full name of protein |
|---|---|
| Protein-tyrosine phosphatases | |
| CD45 | Leukocyte common antigen CD45 receptor-tyrosine phosphatase (LCA, T200) |
| LAR | LCA antigen-related (LAR) receptor tyrosine phosphatase |
| PTP1B | Protein-tyrosine phosphatase 1B |
| PTP1C | Protein-tyrosine phosphatase 1C (SHP1, SHPTP1) |
| PTP1D | Protein-tyrosine phosphatase 1D (SHP2, SHPTP2, Syp, PTP2C) |
| PTP | PEST Protein-tyrosine phosphatase with PEST sequences (PTPG1, PTPN12) |
| VHR | Dual specificity protein phosphatase 3 |
| SIRP1 | Signal regulatory protein substrate of PTP1D phosphatase (SHPS1) |
| Protein-serine/threonine phosphatase | |
| PPP1CA/PP1Cα | Protein-serine phosphatase 1–catalytic subunit–alpha isoforms |
| PPP1CB/PP1Cβ | Protein-serine phosphatase 1–catalytic subunit–beta isoforms |
| PPP1CC/ PP1Cγ | Protein-serine phosphatase 1–catalytic subunit–gamma isoforms |
| PPP2R1A/PP2AαPPP2R1B/PP2Aβ | Protein-serine phosphatase 2A–A regulatory subunit–alpha and beta isoforms |
| PPP2CA/PP2ACα | Protein-serine phosphatase 2A–catalytic subunit alpha isoforms |
| PPP2CB/PP2ACβ | Protein-serine phosphatase 2A–catalytic subunit beta isoforms |
| PPM1A/PP2Cα | Protein-serine phosphatase 2C–catalytic subunit–alpha isoforms |
| PPM1B/PP2Cβ | Protein-serine phosphatase 2C–catalytic subunit–beta isoforms |
| PPM1D/PP2Cδ | Protein-serine phosphatase 2C–catalytic subunit–delta isoforms |
| PPP4R1/PP4/A′2 | Protein-serine phosphatase 4–regulatory subunit (PPX/A′2) |
| PPP4C/PP4C | Protein-serine phosphatase X–catalytic subunit (PPX/C) |
| PPP5C/PP5C | Protein-serine phosphatase 5–catalytic subunit (PPT) |
| PPP6CB/PP6CbPPP6CC/PP6Cc | Protein-serine phosphatase 6–catalytic subunit (PPVC)–b and c isoforms |
| Protein-threonine/tyrosine phosphatases | |
| MKP1/DUSP1 | MAP kinase phosphatase 1 (CL100, VH1) |
| MKP2/DUSP4 | MAP kinase phosphatase 1 (VH2) |
| PAC1/DUSP2 | Dual specificity MAP kinase protein phosphatase |
| CDC25B | Cell division cycle 25B phosphatase |
| CDC25C | Cell division cycle 25C phosphatase |
| PPP3CA/PP2B/Aα | Protein-serine phosphatase 2B–catalytic subunit–alpha isoforms |
| PTEN | Phosphatidylinositol-3,4,5-trisphosphate 3-phosphatase and proteinphosphatase and tensin homolog deleted on chromosome 10 |
| Protein-serine/tyrosine phosphatases | |
| KAP | Cyclin-dependent kinase associated phosphatase (CDK inhibitor 3, CIP2) |