
Abstract
Abstract
Dengue virus infects millions of people worldwide, and there is no vaccine or anti-dengue therapeutic available. Antimicrobial peptides have been shown to possess effective antiviral activity against various viruses. One of the main limitations of developing these peptides as potent antiviral drugs is the high cost of production. In this study, high yield production of biologically active plectasin peptide was inexpensively achieved by producing tandem plectasin peptides as inclusion bodies in E. coli. Antiviral activity of the recombinant peptide towards dengue serotype-2 NS2B-NS3 protease (DENV2 NS2B-NS3pro) was assessed as a target to inhibit dengue virus replication in Vero cells. Single units of recombinant plectasin were collected after applying consecutive steps of refolding, cleaving by Factor Xa, and nickel column purification to obtain recombinant proteins of high purity. The maximal nontoxic dose (MNTD) of the recombinant peptide against Vero cells was 20 μM (100 μg/mL). The reaction velocity of DENV2 NS2B-NS3pro decreased significantly after increasing concentrations of recombinant plectasin were applied to the reaction mixture. Plectasin peptide noncompetitively inhibited DENV2 NS2B-NS3pro at Ki value of 5.03±0.98 μM. The percentage of viral inhibition was more than 80% at the MNTD value of plectasin. In this study, biologically active recombinant plectasin which was able to inhibit dengue protease and viral replication in Vero cells was successfully produced in E. coli in a time- and cost- effective method. These findings are potentially important in the development of potent therapeutics against dengue infection.
Introduction
Cationic peptides have been shown to possess antiviral activity against HIV (Owen et al, 2004), avian influenzaH5N1 virus (Liang et al, 2010), herpes simplex virus (Yasin et al., 2004), and dengue virus (Rothan et al, 2012a, b). Most of the cationic peptides possess antibacterial activity mediated by permeabilization of bacterial membranes (Schneider et al., 2010). It could therefore be hypothesized that the lipid membrane of enveloped viruses such as the dengue virus may also be a potential target of antimicrobial peptides. In addition, these peptides have indeed been shown to block viral infection at the entry step by binding to heparan sulphate, thereby blocking viral attachment (Hazrati et al., 2006; Munk et al., 2003). Interestingly, some of these cationic peptides also showed potential inhibition against viral proteases (Rothan et al., 2012a, b).
Plectasin, an antimicrobial cationic peptide produced by the fungus Pseudoplectania nigrella, is the first fungus-derived defensin with therapeutic potential especially against gram positive bacteria (Schmitz and Holzgabe, 2013). This peptide has secondary structures similar to defensins found in other organisms (Mygind et al., 2005). It consists of 40 amino acids (NH2-MGFGCNGPWDEDDMQCHNHCKSIKGYKGGYCAKGGFVCKCY-COOH) with three intramolecular disulfide bridges between C1 and C5, C2 and C4, and C3 and C6 that stabilize its alpha helix-beta motif tertiary structure (Mandal et al., 2009; Mygind et al., 2005).
Plectasin could be considered as the best candidate for antimicrobial drug development for clinical application due to its low toxicity, immunogenicity, and high stability. It has been shown that plectasin has low ability to induce cytokine production such as interleukin 8 (IL-8), as well as low cytotoxicity against mammalian cells (Liang et al., 2010). Low in vivo toxicity was also observed on mice treated with these peptides (Mygind et al., 2005). Immunization studies were used to assess immunogenic properties of plectasin in mice. After 5 days of daily subcutaneous injections of plectasin, no antibody response, signs of hypersensitivity, or injection site reactions were observed (Brinch et al., 2009). Surprisingly, the antibacterial activity of plectasin was stable over a wide pH range (2.0–10.0). Plectasin also showed high thermal stability (stable at 100°C for 1 h), and remarkable resistance to papain and pepsin (Zhang et al., 2011).
One of the main limitations of using cationic peptides as antiviral drugs is the high cost of production by chemical synthesis. As an alternative, producing recombinant peptides in Escherichia coli may represents one of the most attractive approaches towards large-scale production. However, E. coli almost always produce short peptides in soluble form that require column purification (Hazrati et al., 2006; Jung et al., 2010), which could be considered a bottleneck for large-scale production. In this study, to overcome this problem, five plectasin peptides were designed to be expressed in tandem in E. coli as inclusion bodies. These were then purified by minimal washing steps, followed by solubilization, refolding, and cleaving to facilitate high yield production of single units of plectasin peptides. Subsequent assays on the recombinant plectasin peptide showed that it possessed inhibition activities against dengue NS2B-NS3pro and viral replication in Vero cells.
Methods
Construction of plectasin-coding expression cassette
The DNA sequence was constructed using the Klenow-Pfu polymerase method as described previously (Holowachuk and Ruhhoff, 1995). A five plectasin coding sequence was designed using E. coli optimized DNA sequence and was separated by inserting His tag-linker-FactorXa recognition site, totaling to approximately 763 bp. Long oligonucleotides (60 mer) and amplification primers containing sequences of cleaved XbaI and XhoI restriction sites (22–25 mer each) were designed spanning the length of the synthetic gene with overlapping 3' ends (15 nucleotides) using gene designer DNA 2.0 software. The long oligonucleotides were annealed and the mixture of complementary overlapping oligonucleotides was extended and filled-in with dNTP and Klenow DNA polymerase (Invitrogen, USA). The full-sized DNA product was subsequently amplified by PCR and cloned into pET303/CT E. coli expression vector. PCR conditions through 32 cycles were 95°C for 45 sec as denaturing steps, 60°C for 45 sec as annealing steps, and 72°C for 1 min as elongation steps. After cloning, recombinant plasmids were isolated, purified, and sequenced to verify that the sequences were correct and without unwanted mutations. The desired double-stranded DNA fragment was digested with XbaI and XhoI restriction enzymes (Promega, USA), purified using the gel extraction kit (Qiagen, USA), and cloned directly into pET303/CT His vector. Then, transformation into E. coli BL21 competent cells was performed.
Protein expression
Single colonies of cells containing recombinant plasmid pET 303/CT His-plectasin were used to inoculate 10 mL of LB broth. After overnight incubation at 37°C, the primary culture was used to inoculate 1 L of expression medium containing 100 μg/mL ampicillin, 10 g/L tryptone, 5 g/L yeast extract, 5 g/L NaCl, and 2 g/L glucose. The subculture was grown at 37°C with good aeration (250 rpm with shaking) until OD600 was approximately 0.9–1.0. A sample of 1 mL was taken as non-induced cells and centrifuged for 2 min; the cell pellet was then resuspended in 50 μL of SDS-PAGE sample buffer and frozen at −20°C. For induction, isopropylthiogalactoside (IPTG) was added to the remaining culture to a final concentration of 0.8 mM and the culture was incubated at 37°C with shaking at 250 rpm for 24 h. Samples of 1 mL were taken and prepared for SDS-PAGE analysis.
Inclusion bodies isolation and solubilization
Bacterial cells were harvested and resuspended in lysis buffer (Tris-HCl 50 mM, NaCl 100 mM, lysozyme 500 mg, 1% Triton x-100, DDT 10 mM, 1 mM EDTA). The cells were lysed by sonication at 400 W for 15 min on ice. The lysed cells suspension was centrifuged for 20 min, 6000 rpm at room temperature. The precipitated inclusion bodies were washed three times with washing buffer (Tris-HCl 50 mM, NaCl 100 mM, DDT 10 mM, 1 mM EDTA), followed by two times with dH2O. One gram of pure inclusion bodies was resuspended into 250 mL dH2O by continuous short sonication. Then, inclusion bodies were solubilized by adding 0.2 mL of 10 N NaOH and incubated for 30 min at room temperature.
Refolding of recombinant plectasin
Recombinant plectasin polypeptide was refolded as previously described Sijwali et al., (2001). In brief, the pH of solubilized polypeptide was adjusted to 12.5. Then, disulfide bonds were reduced by 5 mM of β-mercaptoethanol and incubated at room temperature for 30 min with stirring. The denatured and reduced peptide was diluted to a final concentration of 0.01 mg/mL with ice-cold refolding buffer (100 mM Tris-HCl, 1 mM EDTA, 10% glycerol, 250 mM L-arginine, 1 mM reduced glutathione, and 0.5 mM oxidized glutathione [pH 12.5]). Protein sample was loaded into a dialysis tube and dialysis was carried out overnight against 200 volumes of 100 mM Tris-HCl, pH 10 to eliminate the residuals of β-mercaptoethanol. Overnight incubation of peptide samples at 4°C was performed before the refolded polypeptide was concentrated to 1 mg/mL by Vivaspin 50 mL concentrating tubes with a 3000 MW cut-off membrane (Sartorius, Germany). Then, the concentrated polypeptide was incubated with Factor Xa for 4 h at room temperature to release single units of plectasin by cleaving polypeptide construct.
Column purification
Nickel column purification was applied according to the manufacturer's instructions (His GraviTrap™ Flow precharged Ni Sepharose™ 6 Fast column, Amersham Biosciences, USA). In brief, the column was equilibrated with phosphate buffer (20 mM sodium phosphate buffer and 500 mM NaCl, pH 7.4). The sample was loaded into the column, which was washed with binding buffer (phosphate buffer containing 20 mM imidazole, pH 7.4). The recombinant protein was eluted with elution buffer (phosphate buffer containing 200 mM imidazole, pH 7.4).
Antimicrobial activity
The antimicrobial activity of recombinant plectasin was assessed using inhibition zone assays (Zhang et al., 2011). In brief, recombinant plectasin (1 mg/mL) were dropped onto a filter paper discs and left to dry at room temperature. Filter paper discs saturated with column elusion buffer were used as a negative control. The filter paper discs were then placed on an S. aureus- inoculated Mueller-Hinton agar using two separated plates which were then incubated 24 h at 37°C.
Maximal nontoxic dose (MNTD)
To determine the MNTD value of recombinant plectasin, Vero cells were cultured at 0.5×105 cell/well in 96-well plates for 24 h at optimal conditions (37°C, 5% CO2 in humidified incubator). Then, the growth media (10% FBS) were removed and the cells were washed two times with PBS. New maintenance DMEM medium (2% FBS) containing 0.0, 5.0, 10, 20, 40, and 80 μM of recombinant plectasin were added and the cells were incubated for 24, 48, and 72 h. Triplicate wells were analyzed for each concentration, and column elusion buffer was used as control. A ten microliter solution of freshly prepared 5 mg/mL MTT in PBS was added to each well and allowed to incubate for an additional 2 h. Afterwards, the media was removed and DMSO was added at 100 μL/well. Plates were then swirled gently to facilitate formazan crystal solubilization. The absorbance was measured at 570 nm using a microplate reader (Tecan Infinite M200 Pro, Switzerland). Percentage of cell viability was calculated as follows: 100-(Absorbance of peptide treated cells/Absorbance of untreated cells)×100.
In vitro Dengue protease assay (NS2B-NS3pro)
The ability of plectasin to inhibit DENV2 dengue serine protease (NS2B-NS3pro) was examined as described previously (Rothan et al., 2012a, b). DENV2 NS2B-NS3pro was produced in E. coli as a single chain containing 48 amino acids of NS2B and 185 amino acids of NS3 (Yon et al., 2005; Yusof et al., 2000). A standard reaction mixture (200 μL) was prepared in black 96-well plates containing 200 mM Tris-HCl (pH 8.5), 2 μM recombinant NS2B-NS3pro, 5 mM NaCl, 15% glycerol, and varying concentrations of plectasin peptide. Incubation for 30 min at 37°C was performed before starting the reactions by adding 100 μM of fluorogenic peptide substrate ([Boc-Gly-Arg-Arg-AMC] Osaka, Japan, Lot no: 580907). Further incubation was performed for 30 min at 37°C and the release of free AMC was measured using Tecan Infinite M200 Pro fluorescence spectrophotometer (Tecan Group Ltd., Switzerland). Substrate cleavage was normalized against buffer only (control) at the emission of 440 nm upon excitation at 350 nm. Fluorescence values obtained with the no-inhibitor control was taken as 100%, and values in the presence of inhibitors was calculated as the percentage of inhibition of the control using GraphPad Prism 5 software under non-linear regression.
Treatment of DENV2- infected cells with recombinant plectasin
Vero cells were grown in a 24-well tissue culture plate (1×105 cells/well), incubated 24 h under optimal conditions (37°C and 5% CO2). DENV2 was added to the wells (MOI of 2) followed by incubation for 1 h with gentle shaking every 10 min for optimal virus to cell contact. The virus supernatant was removed, and the cells were washed twice with fresh serum free DMEM media to remove residual virus. New complete DMEM media containing 5, 10, and 20 μM of peptide were added and the cultures were incubated for 24, 48, and 72 h. Afterwards, cellular supernatants were collected. For quantification of DENV2 copies, a standard curve was generated by 10-fold serial dilution of known copies of DENV2 RNA. Viral RNA was extracted from culture supernatant using QIAmp viral RNA mini kit (Qiagen, Germany), according to the manufacturer's instructions. A fragment located at the 5'UTR region of the virus genome was used as reference to design the primers. One-step RT-PCR using SYBR Green Master Mix (Qiagen, Germany) was used to perform absolute quantification using ABI7500 machine from Applied Biosystems (Foster City, CA). The PCR programme included 1 cycle of 50°C for 2 min, 1 cycle of 95°C for 10 min, and 40 cycles of 95°C for 15 sec and 60°C for 1 min. Dissociation curve analysis was added at the end of each run. Results were analyzed using Sequence Detection Software Version 1.3.
Statistical analysis
Statistical analyses were performed using GraphPad Prism version 5.01 (GraphPad Software, San Diego, CA). P values of <0.05 were considered significant. Error bars are expressed as ±SD.
Results
Production of recombinant plectasin in E. coli
The DNA coding for plectasin polypeptide expression cassette was constructed in vitro using Klenow-pfu DNA polymerase method. The molecular weight of plectasin peptide is 4.4 kDa, while the whole expression cassette that comprises 5 units of the peptide, 5X His tags, and the linkers was approximately 27 kD (Fig. 1A and 1B). Plectasin polypeptide was produced insolubly as inclusion bodies at high level reaching approximately 85% of the total bacteria protein. The refolded polypeptide was incubated with Factor Xa to release recombinant plectasin in the form of single units (Fig. 1C). In order to eliminate the residual components of Factor Xa, the cleaved polypeptides were purified by Ni-affinity chromatography (Fig. 1D). The biological activity of the recombinant peptide was confirmed by showing high inhibition potential against S. aureus (Fig. 2). Using the above strategies, we managed to produce approximately 75 g of recombinant plectasin per liter of E.coli cell culture.
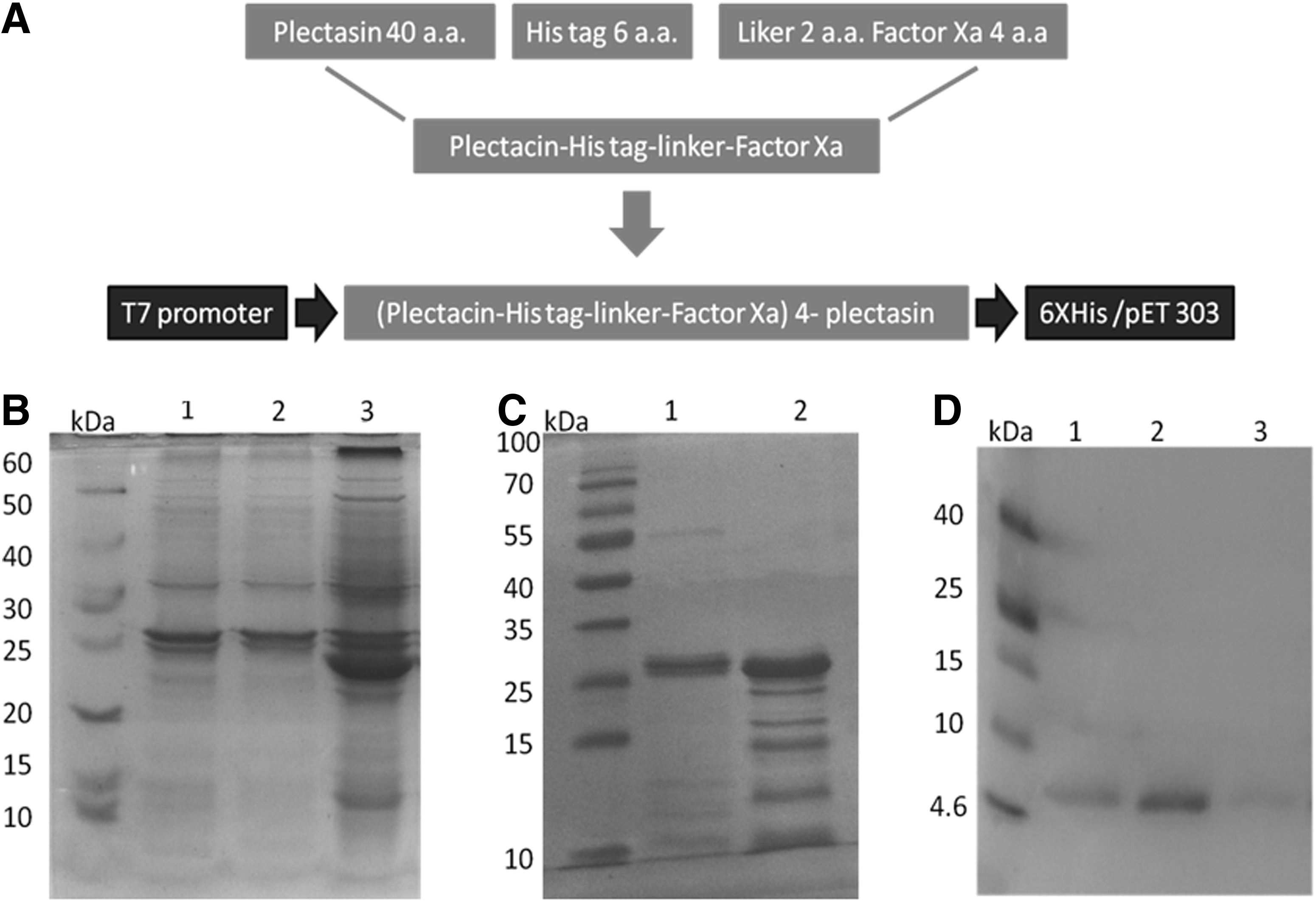
Production of recombinant plectasin in E coli.
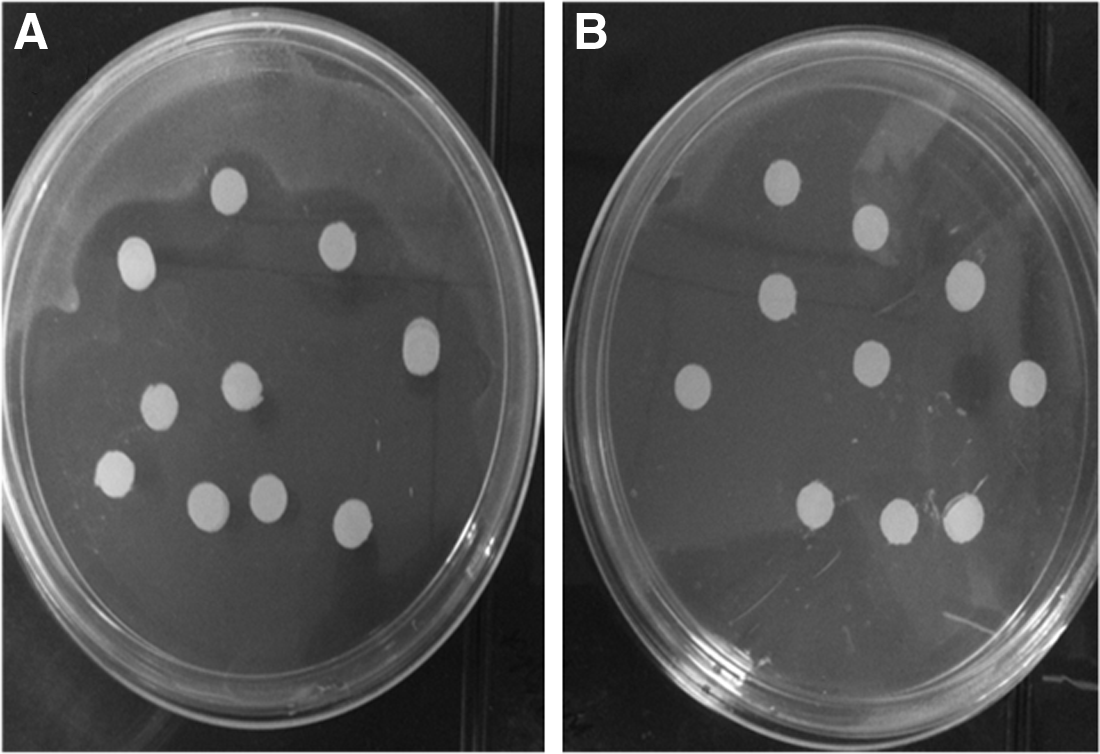
Antimicrobial activity of recombinant plectasin using inhibition zone assays. (A) Inhibition of S. aureus growth by 1 mg/mL of recombinant plectasin dropped onto filter paper discs.
Inhibition of dengue NS2B-NS3pro by recombinant Plectasin
DENV2 NS2B-NS3pro was produced in E. coli as a single chain by joining NS2B unit with NS3 by 9 amino acids linker. The activity of the recombinant enzyme was tested using fluorogenic peptide substrate. Increasing concentrations of recombinant plectasin were added to the reaction mixture that contained the NS2B-NS3pro and its substrate. After incubation at 37°C, the release of free AMC was measured by fluorescence spectrophotometry. The results of this study showed that the recombinant plectasin inhibited dengue NS2B-NS3pro at Ki value of 5.03±0.98 μM. The mode of inhibition could be concluded using the parameters of plectasin inhibition that are represented in Table 1. A kinetic analysis was carried out to determine the ability of plectasin to inhibit dengue NS2B-NS3pro at different concentrations. Increasing concentrations of the inhibitor were not able to change the apparent Michaelis–Menten constants (Km value, 96.7±5.8 μM). This finding indicates that the inhibitor was able to reduce enzyme activity by disrupting its stability but not reducing the affinity between the enzyme and the substrate. This type of inhibition is a characteristic feature of a noncompetitive mode of inhibition (Fig. 3).

Kinetic assay plot for the inhibition of dengue NS2B-NS3pro by plectasin peptide. This assay was carried out using increasing concentration of inhibitor while all other conditions were kept constant. The data were analyzed by Michaelis-Menten model under nonlinear regression curve fit in GraphPad Prism 5 software. The peptide concentrations were 0, 2, 4, and 8 μM. There is marked decrease in the maximum enzyme velocity when plectasin concentration was 2.0 μM, and more than 80% reduction was observed at 8.0 μM of plectasin peptide.
Alpha, Constant that determines mechanism; Ki, Inhibition constant (μM); Km, Michaelis-Menten constant (μM); Vmax, Maximum enzyme velocity (μM/min). If Alpha=1, this is the same as noncompetitive. If Alpha is very high, then the model approaches a competitive model. If alpha is very low (but greater than zero), the model approaches an uncompetitive model. All parameters were calculated by GraphPad Prism 5.0 software.
Toxicity of recombinant plectasin
The present study showed that the MNTD value of recombinant plectasin peptide was approximately 20 μM (100 μg/mL) that was observed at 24, 48, and 72 h (Fig. 4). Therefore, concentrations that are equal and lower than the MNTD value were used to test the ability of recombinant plectasin to inhibit DENV2 replication in Vero cells.
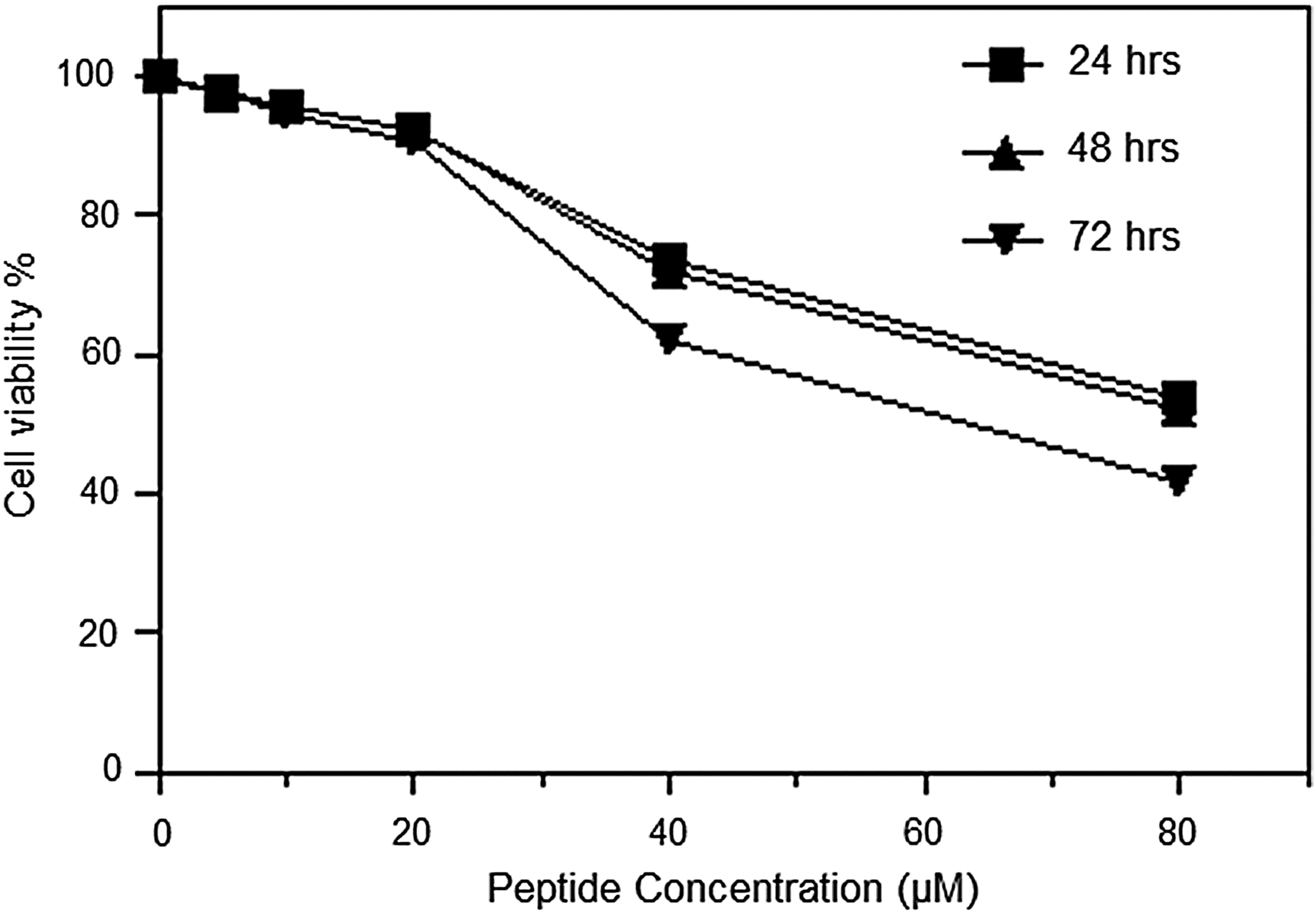
Maximum nontoxic dose (MNTD) of plectasin peptide. Vero cells were seeded at 0.5×105 cells/well in triplicate at optimal conditions (37°C, 5% CO2 in humidified incubator) in 96-well plates. Plectasin was diluted to serial concentrations 5, 10, 20, 40, and 80 μM with DMEM media supplemented with 2% FBS. The cell culture was analyzed at 24, 48, and 72 h using MTT assay. The toxic dose of Vero cells was identified to be greater than 20.0±0.12 μM.
Inhibition of dengue replication in Vero cells by recombinant plectasin
Based on the MNTD value, recombinant plectasin at concentrations of 5.0, 10.0 and 20.0 μM were used to treat Vero cells infected with DENV2. The viral copy number was significantly reduced after treatment with recombinant plectasin at 24, 48, and 72 h (Fig. 5A). The highest reduction in dengue virus replication was observed at 24, 48, and 72 h when plectasin concentration was 20.0 μM (Fig. 5B). Plectasin showed sustainable inhibitory effects against dengue replication in Vero cells throughout the time points of study. DENV2-infected cells were characterized by the occurrence of cytopathic effects (CPE) that includes irregular, round, spindle shaped cells and apoptotic cells, as well as the failure of treated cells to maintain the monolayer sheet. DENV2-infected cells treated with 20.0 μM of recombinant plectasin showed an ability to maintain its monolayer with notable reduction of viral CPE and low amount of apoptotic cells (Fig. 5C).
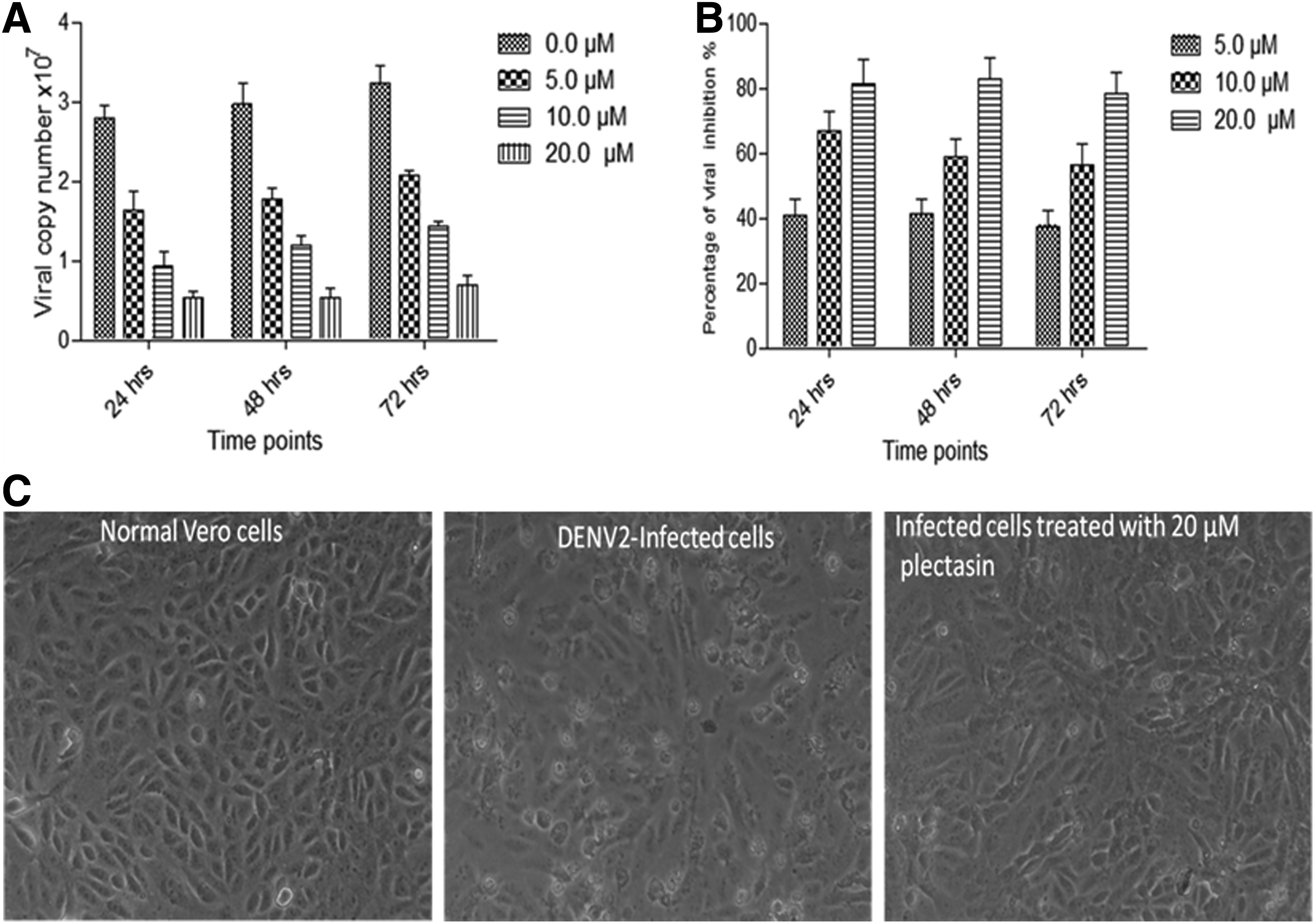
Inhibition of DENV2 replication in Vero cells after applying different concentrations of recombinant plectasin. Vero cells were infected with DENV2 and treated with 5, 10, and 20 μM of plectasin peptide for 24, 48, and 72 h. Cellular supernatants were collected and DENV2 copies were quantified using quantitative RT-PCR.
Discussion
Dengue virus infects people in more than 100 tropical countries, causing severe syndromes that lead to serious economic burden (Gubler, 2002). Elevated global interest in developing new antipoverty vaccines and drugs would dramatically increase the current support for new neglected tropical diseases technologies (Hotez, 2011). In this study, we describe an innovative strategy for low cost production of natural cationic peptide that showed significant inhibition against dengue virus. Natural cationic peptides have been considered as best candidates for designing and developing new anti-dengue therapeutics due to their low toxicity, immunogenicity, and high stability (Brinch et al., 2009; Hara et al., 2008; Mygind et al., 2005; Zhang et al., 2011).
In this study we show that high yield production of biologically active peptide was inexpensively achieved by producing tandem plectasin peptides as inclusion bodies in E. coli. The inclusion bodies of plectasin polypeptide were solubilized by alkaline shock and refolded in an alkaline and diluted environment in the presence of redox agents. The refolding strategy adopted in this study was based closely on a previously described cloning effort by Sijwali et al. (2001). The authors successfully produced cysteine-rich protease falcipain-2 in E. coli; the importance of an alkaline and diluted environment was emphasized as it reduced the possibility of inter-molecule disulfide bond formation.
Previous reports have also shown that recombinant plectasin can be produced in E. coli as fusion peptide with Trx protein (Jing et al., 2010) and as a secreted protein in Pichia pastoris (Jing et al., 2010; Mao et al., 2010). These methods reported production of milligrams of recombinant plectasin in one liter of cell culture. In the present study, a significant increase in recombinant plectasin yield was obtained (75 g/L of E. coli cell culture) by strategically expressing repeated units of the peptide as inclusion bodies in E. coli, followed by simple purification. Perhaps more importantly, this was achieved at relatively low cost and would be attractive in support of the development of antipoverty drug initiatives (Hotez, 2011).
The current study shows that the recombinant plectasin peptide significantly inhibited dengue NS2B-NS3pro and subsequently viral replication in vitro. It has indeed been shown that the dengue NS2B-NS3pro function is essential for viral replication. Dengue NS2B-NS3pro cleaves viral polyprotein to structural and nonstructural proteins after translation of viral RNA using host cell ribosomes (Geiss et al., 2009; Stadler et al., 1997). The results of this study show that recombinant plectasin inhibited dengue NS2B-NS3pro at Ki value of approximately 5.0 μM, which is more potent than some of the other low molecular weight inhibitors that showed a range of inhibitory activities against dengue NS2B-NS3pro (Tomlinson et al., 2009). It is known that the binding between the dengue protease subunits depends on the interaction between negatively charged amino acids in NS2B and positively charged amino acids in NS3 (Erbel et al., 2006). Therefore, plectasin as a cationic peptide may inhibit the activity of dengue protease by disrupting the binding between enzyme subunits that, in turn, lead to significant reduction in its activity. A variety of different flavivirus protease inhibitors have been previously reported. Most of these inhibitors have charged moieties such as peptide-aldehyde inhibitors (Knox et al., 2006), short cationic peptides inhibitors (Stoermer et al., 2008), and nonpeptidic guanidine compounds (Ganesh et al., 2005). Furthermore, a series of noncharged 8-hydroxyquinoline compounds have also been identified (Mueller et al., 2008). However, most of these compounds have a wide range of cytotoxicity, stability, and other concerns related to the cost of synthesis and large scale production. On the other hand, recent findings have shown that the use of plectasin has several advantageous. Studies have reported low cytotoxic effect of plectasin against mammalian cells (Hara et al., 2008), as well as low in vivo toxicity using mice as an animal model (Mygind et al., 2005). In addition, the cationic plectasin peptide has high stability at different pH values and temperatures, and is less susceptible to proteases such as papain and pepsin (Zhang et al., 2011). Our results corroborated these observations and also showed that concentrations at maximal nontoxic dose (MNTD value) of plectasin significantly reduced viral replication in Vero cells. The observed effect is likely mediated by the inhibition of the viral NS2B-NS3 protease. Further work would be necessary to understand the exact mechanism(s) responsible for the inhibitory effect of plectasin against viral replication.
In conclusion, this is the first study to describe the inexpensive high-yield production of recombinant plectasin through the tandem-protein expression strategy, and the characterization of its antiviral activity. We successfully produced high quantities of biologically active recombinant plectasin in E. coli using a time- and cost-effective method that can be developed and adopted for large-scale production toward the development of therapeutic agents against dengue infection.
Footnotes
Acknowledgments
This project was funded by the University of Malaya and Ministry of Science, Technology and Innovation (IPharm Grant 53-02-03-1049).
Authors' contributions; HAR design performed the experiments and drafted the manuscript. ZM and MAS participated in the analysis and preparation of the manuscript. NSR and RY participated in the design and drafted the manuscript. All authors approved the final manuscript.
Author Disclosure Statement
The authors declare that they have no competing interests.