
Abstract
Abstract
Discovery of new antibacterials with novel mechanisms is important to counteract the ingenious resistance mechanisms of bacteria. In this connection, omics-guided drug discovery offers a rigorous method in the quest of new antibacterials. (-)-Roemerine is a plant alkaloid that has been reported to possess putative antibacterial activity against Escherichia coli, Bacillus subtilis, and Salmonella typhimurium. The aim of the present study was to characterize the activity of (-)-roemerine in Escherichia coli TB1 using proteomics tools. With (-)-roemerine treatment, we found limited permeability through the outer membrane and repression of transport proteins involved in carbohydrate metabolism, resulting in poor carbon source availability. The shortfall of intracellular carbon sources in turn led to impaired cell growth. The reduction in the abundance of proteins related to translational machinery, amino acid biosynthesis, and metabolism was accompanied by a nutrient-limited state. The latter finding could suggest a metabolic shutdown in E. coli cells. High osmolarity was clearly not one of the reasons of bacterial death by (-)-roemerine. These observations collectively attest to the promise of plant omics and profiling of putative drug candidates using proteomics tools. Omics-guided drug discovery deserves greater attention in mainstream pharmacology so as to better understand the plants' medicinal potentials.
Introduction
T
With the progress made in the analysis of biological systems using high-throughput technologies, investigation of the activities of medicinal plants has gained pace (Sahu et al., 2014). Medicinal plants are in the heart of search for new multi-target bioactive molecules since plant extracts or plant-derived compounds are found as natural antibacterials and resistance-modifying agents that impede bacterial resistance mechanisms (Abreu et al., 2012; Sibanda and Okoh, 2007). Alkaloids, flavonoids, and terpenoids are some of the families of plant secondary metabolites that are considered as attractive antibacterial candidates (Wink, 2008). Among these, alkaloids are known as nitrogenous compounds derived from amino acids such as tyrosine, tryptophan, phenylalanine, arginine, and lysine (Croteau et al., 2000). These molecules commonly impose their effects by forming hydrogen bonds with drug targets. Therefore, their scaffolds are found in various therapeutic drugs and lead compounds (Kittakoop et al., 2014).
Moreover, some alkaloids and their derivatives such as morphine, quinine, etoposide, colchicine, and reserpine possess significant medicinal values (Baldwin and Osheroff, 2005; Croteau et al., 2000; Doyle et al., 1955; Terkeltaub et al., 2010). Regarding their antibacterial mechanism, they are capable of binding to DNA and RNA (e.g., berberine, sanguinarine, palmatine, coralynena, and jatrorhizine) (Bhadra and Kumar, 2011), and inhibiting efflux pumps (e.g., reserpine) (Neyfakh et al., 1993), type II isomerase (e.g., quinolones) (Heeb et al., 2011), topoisomerase I and II (e.g., protoberberines) (Kobayashi et al., 1995), and bacterial division protein FtsZ (e.g., berberine) (Boberek et al., 2010). The variety in their intracellular binding partners suggests that they could be multi-target molecules.
(-)-Roemerine is an aporphine-type alkaloid possessing bioactive properties that includes antibacterial (Liou et al., 1979), antifungal (Agnihotri et al., 2008; Rao et al., 2009), antimalarial (Agnihotri et al., 2008; Baghdikian et al., 2013), anti-HIV (Kashiwada et al., 2005), and vasorelaxant activities (Chulia et al., 1995; Valiente et al., 2004). Despite its prevalent presence in the Papaveraceae family (Sariyar, 2002; Sariyar and Kalav, 1990; Sariyar and Oztekin, 1981), studies with this alkaloid are very limited.
In the current study, we used a proteomics approach to elucidate the affected pathways in Escherichia coli TB1 upon treatment with (-)-roemerine to propose hypotheses regarding the activity of this alkaloid in these cells. To test our hypotheses, genes found on related pathways were selected and analyzed by quantitative real-time PCR (qPCR).
Escherichia coli was chosen as the model organism since it is the most intensively studied microorganism from a genetic perspective (Blattner et al., 1997). Moreover, its metabolic pathways are reliably and comprehensively captured in several web-accessible databases (KEGG, MetaCyc, EcoCyc) that enables formation of a complete story in a biological context.
Materials and Methods
Chemicals and bacterial strain
All chemicals, solutions, and kits used in this study were purchased from Applichem (Germany), Merck (Germany), Molekula (Germany), Sigma (USA), Roche (Germany), Fermentas (USA), and Bio-Rad (Hercules, CA). Pure (-)-roemerine alkaloid from Papaver rhoeas (Sariyar, 2002) was a kind gift from G. Sariyar (Istanbul University, Faculty of Pharmacy). (-)-Roemerine was dissolved in dimethyl sulfoxide (DMSO). E. coli TB1 (F- ara Δ(lac-proAB) [Φ80dlac Δ(lacZ)M15] rpsL(StrR) thi hsdR) used as the model organism was from our laboratory stocks.
Cell growth and (-)-roemerine treatment
E. coli TB1 cells were grown in Luria-Bertani (LB) medium (per liter: 10 g tryptone, 5 g yeast extract, 10 g NaCl) at 37°C and 180 rpm. Cells were treated with 0-1×MIC (-)-roemerine when the OD600 of the bacterial culture reached 0.54±0.06. Control cells were treated with equal amount of DMSO. Growth was spectrophotometrically monitored at OD600 and the recordings were plotted as a function of time.
The number of viable cells, as colony-forming units (CFU), were estimated by plating the (-)-roemerine treated and control cells with serial dilutions on LB agar media. Colonies formed were counted after an overnight incubation at 37°C and the mean values from triplicate experiments were calculated.
Minimum inhibitory concentration (MIC) of (-)-roemerine for E. coli TB1 was determined using broth microdilution method (Amsterdam, 1996). MIC was defined as the lowest concentration of the alkaloid that inhibited cell growth following an overnight incubation at 37°C.
Protein extraction, analysis, and identification
Protein extraction was achieved as previously described (Ozbalci et al., 2010). Cells treated with DMSO or 1X MIC (-)-roemerine for 1 h were harvested by centrifugation for 20 min at 6000 g and 4°C. Cell pellets were rinsed twice with 50 mM Tris buffer (pH 7.8) and re-centrifuged under the same conditions. Cellular proteins were extracted using ProteoPrep® Sample Extraction Kit (Sigma, PROT-TOT). Obtained protein samples were cleaned with Zeba Spin Desalting columns, 7K MWCO (ThermoScientific). Bradford dye protocol was used to determine the protein concentration (Bradford, 1976). Bovine serum albumin was used as the standard.
Proteins extracted from three biological replicates were separated by 2-dimensional gel electrophoresis (2-DGE) using immobilized pH gradient (IPG) strips of 17 cm (pH of 4-7, Bio-Rad) in the first dimension and SDS-PAGE in the second dimension (Bjellqvist et al., 1982). The required reagents for 2-DGE were from ReadyPrep™ 2-D Starter Kit (Bio-Rad). Protein spots on 2-D gels were visualized using CBB-G250 dye method (Neuhoff et al., 1988). Stained gels were scanned by GE HEALTCARE® Image Scanner II system and then gel images were analyzed by REDFIN software (Bio-Rad) to find the changes in the intensity of the protein spots. The list of spots was statistically ordered by p-value from the one-way ANOVA analysis. By going through the spot rank table, top ranked protein spots that had a threshold≥1.8 fold change with a p value<0.05 were regarded as significant.
Identification of selected protein spots was performed at the University of York, UK (Proteomics Laboratory of Department of Biology) based on MS/MS fragmentation data, which filtered matches to accept only peptides with a score of 0.05 or lower. The obtained spectral data was then searched against a subset of the EcoProt database (4284 sequences; 1351035 residues).
Total RNA extraction, cDNA synthesis, and qPCR
Total RNA was extracted with High Pure RNA Isolation Kit (Roche). RNA concentration was measured by Qubit® 2.0 Fluorometer (Invitrogen). cDNA was synthesized with Transcriptor High Fidelity cDNA synthesis kit (Roche).
SYBR Green I (LightCycler® FastStart DNA MasterPLUS SYBR Green I, Roche) based qPCR analysis was performed by LightCycler 1.5 Instrument (Roche). The primers used for each target gene amplification are listed in Table 1. qPCR was carried out in a 20 μL reaction volume with 2–4 ng cDNA, 4 μl of 5X SYBR Green I mix, and 125 nM of primers. The reaction conditions were 95°C for 10 min for denaturation followed by 40 cycles of 95°C for 10 sec, 57°C for 10 sec for annealing, and 72°C for 45 sec for extension. Melting curve analysis was performed to analyze primer dimer formation and other artefacts. Data normalization was performed using 16S rRNA as the housekeeping gene. The fold change in each gene was calculated based on the 2-ΔΔCT method (Livak and Schmittgen, 2001). Averages from at least duplicate analysis were taken.
F designates forward; R, reverse primer.
Functional interaction network analysis of proteins
The functional interaction network integrating proteomics and qPCR results was generated by the aid of STRING 9.1 database's active prediction methods with medium confidence score (http://string-db.org/) (Franceschini et al., 2013). In brief, each protein or gene was used as an input and E. coli was selected as the organism. Output was restricted to “no more than 10 interactors.”
Results
Growth profiles of E. coli cells exposed to (-)-roemerine
(-)-Roemerine was found to inhibit the growth of E. coli TB1 with a MIC value of 100 μg/mL. As spectrophotometrically measured, the growth rate of E. coli was not appreciably affected by DMSO, the solvent used to dissolve (-)-roemerine (Fig. 1).

The effect of 1% DMSO (v/v) on cellular growth. Symbols ( Δ ) and (◯) denote growth profiles of control and DMSO-treated cells, respectively.
Taking the MIC value as the upper limit, logarithmic phase cells were treated with different concentrations of (-)-roemerine (Fig. 2). Comparative analysis of control and (-)-roemerine-treated cells revealed that cell growth was subtly affected in the presence of 0.25X MIC of alkaloid. Increasing (-)-roemerine concentration to 0.5X MIC retarded cell growth, however, cell growth almost entirely ceased 1 h after treatment with 1X MIC. These findings are compatible with the growth rates calculated after treatment with DMSO and (-)-roemerine (Table 2). The growth rates of control and 0.25×MIC (-)-roemerine-treated cells were similar. However, increasing (-)-roemerine concentration to 0.5X MIC and 1X MIC reduced growth rates approximately by 30% and 60%, respectively.
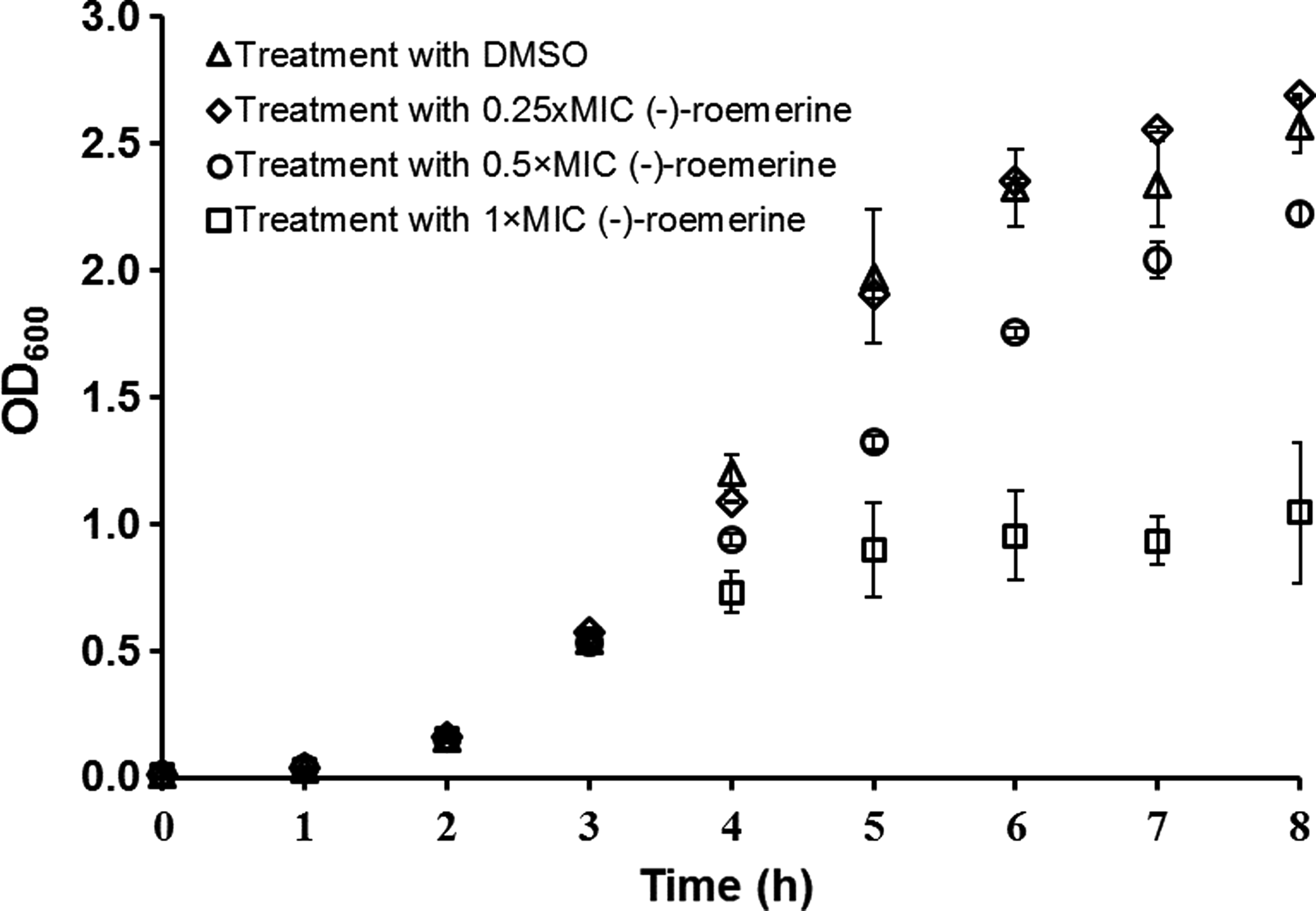
The effect of different concentrations of (-)-roemerine on cellular growth. Arrow indicates time of (-)-roemerine treatment.
The substantial reduction in cell growth with 1×MIC (-)-roemerine was also demonstrated by the drop in the number viable cells (Fig. 3). The number of viable cells dropped to one-third of its value, from 5.2×108 to 1.7×108, 1 h after treatment with 1X MIC (-)-roemerine. The presence of live cells under this condition urged us to analyze the antibacterial mechanism of (-)-roemerine with these cells.

CFU of cells treated with DMSO (dark bars) and 1×MIC of (-)-roemerine (light bars). Treatment was achieved at the third hour when OD600 of cells was 0.54±0.06.
Protein profiles of E. coli TB1 exposed to (-)-roemerine
The comparative analysis of protein samples separated by 2-DGE returned a total of 174 protein spots with a change in their intensities. Of these, 16 with a cut-off value of ≥1.8 and with significant spot intensities were analyzed for identification and the obtained results are summarized in Table 3. Two protein spots with similar abundances (marked as ‘R1 and R2’) were randomly selected as control proteins. Consistent results found for the control proteins (rpIL) confirmed the reliability of the analysis.
C, Control protein.
Among the 16 identified proteins, the abundance of nine spots decreased, while the intensity of the seven spots increased in the presence of (-)-roemerine (See Supplementary Figs S1 and S2; supplementary material is available online at www.liebertpub.com/omi). These proteins were found to function in diverse metabolic pathways such as amino acid biosynthesis, transport, and metabolism (L-asparaginase II, yjgF, and glutamine binding protein, glnH), protein biosynthesis (EF-Ts, EF-Tu), bacteriosin transport (TolB), outer membrane integrity (OmpX), cell redox homeostasis (DsbA), cell adhesion (FimA), D-glucarate catabolic process (GarR), and carbohydrate transport (MalE, MalM and MglB).
Identification results for three different but closely spaced spots (S1, S2, and S3) returned the same protein, MalE. This could probably be due to the presence of three distinct isoforms, as found and suggested elsewhere (Gooley and Packer, 1997; Karaosmanoglu et al., 2014; Stancik et al., 2002). Two equally possible results were found for one spot with increasing intensity (TolB or TufA/B for S50). High scores obtained for both findings made it impossible to eliminate either one. Nevertheless, the location of the TufA on the 2-D gels was not consistent with the findings of Mee-Jung et al. (2012) which led us to assume that this result could be misleading.
Transcriptional analysis of selected genes in E. coli TB1 exposed to (-)-roemerine
Based on the finding from proteomic analysis, it was possible to propose two hypotheses on the effect of (-)-roemerine; one regarding the change in the osmolarity of the medium, and the other regarding the integrity of the cell membranes. To test these hypotheses, several groups of genes were selected for analysis with qPCR. In Table 4, the selected genes are listed. Among the osmolarity related genes were the components of the EnvZ/OmpR osmo-regulatory system and the genes of its target proteins OmpF and OmpC. The integrity of the outer membrane was sought through the fate of the TolB-PAL complex. In addition to these genes, three proteins common with the proteomic analysis, TolB, OmpX, MalE, were specifically selected for validation of proteomic findings. The gene rrsG was used as the housekeeping gene for quantitative analysis since the transcriptional level of this gene was found to remain constant upon treatment with (-)-roemerine.
Post-amplification melting curve analysis confirmed that the primers were specific and there was no dimer formation. The fold changes of all target genes calculated by the 2-ΔΔCT method of Livak and Schmittgen (2001) are presented in Figure 4. A cut-off value of≥1.8 was accepted as significant. Figure 4 revealed that ompX and ompR were upregulated, while ompF, ompC, tolB, pal, and, otsB were downregulated after treatment with (-)-roemerine. Under the same conditions, the transcript level of envZ gene remained unchanged. The fold changes of the two genes, malE and otsA, could not be calculated since their expressions were too low to be detected.
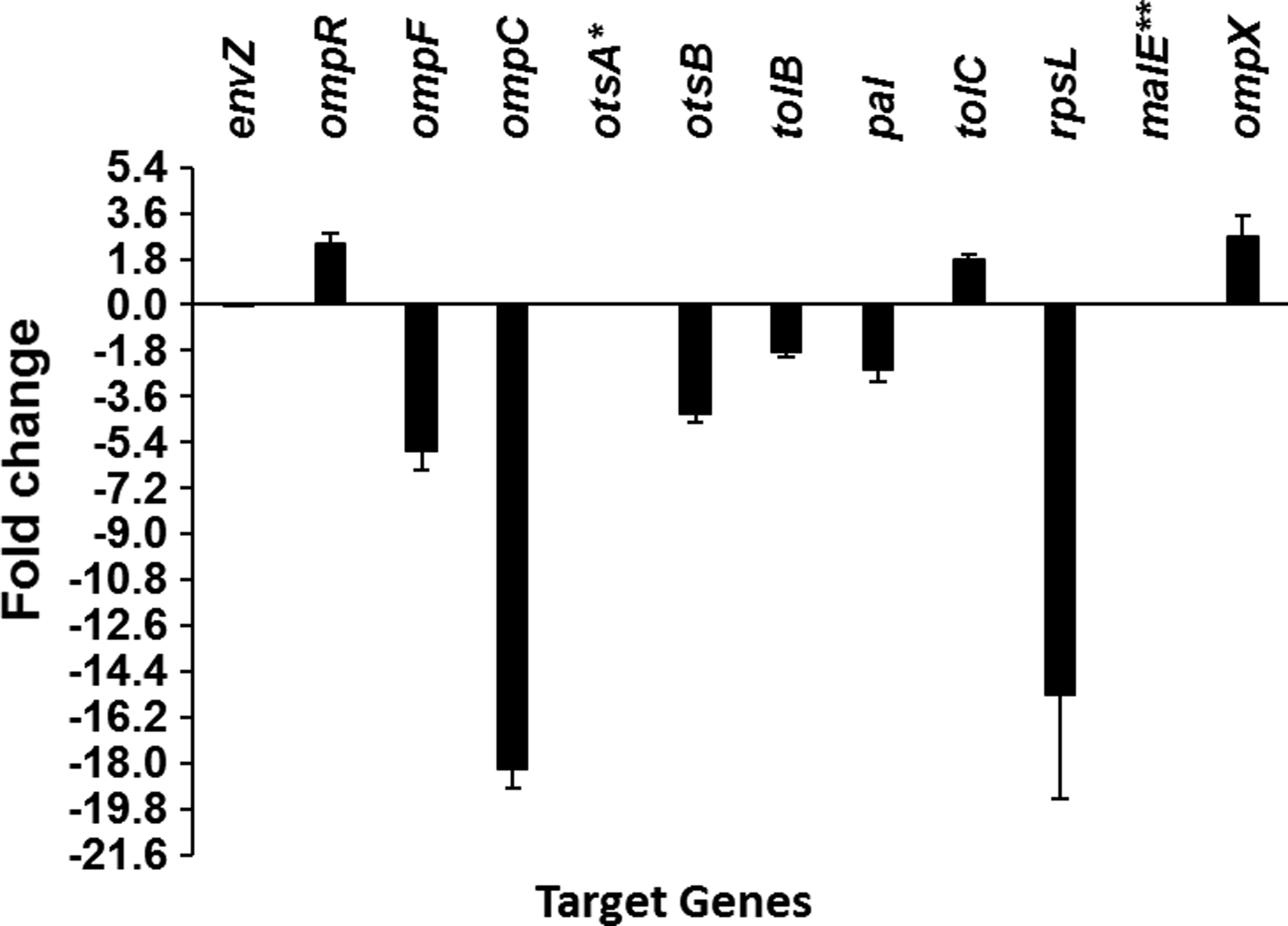
Transcriptional analysis of selected genes subjected (-)-roemerine treatment. *There was no otsA expression in both control and (-)-roemerine-treated cells. **The expression of malE in (-)-roemerine-treated cells was too low to calculate change in expression
The results obtained from proteomic and qPCR analyses for MalE and OmpX were consistent (Table 3 and Fig. 4), however the result obtained for TolB was contradictory. The spot intensity of TolB increased, whereas the transcriptional analysis showed that tolB was downregulated.
Interaction network of differential expression
The regulation patterns of the proteins and genes are illustratively summarized in Figure 5. The target points were then integratively analyzed using the STRING 9.1 database to extract interactions among these targets. The interaction network constructed is depicted in Figure 6. Among the targets identified, 15 were found to be connected with each other. In short, interaction was found among the proteins of maltose and galactose transport systems encoded by malE, malM, and mglB. Simultaneously, interactions were found between MalE and the outer membrane proteins encoded by ompF and ompC. The interaction between the periplasmic protein of the cell envelope Tol complex, TolB, and the peptidoglycan-associated outer membrane lipoprotein, PAL, was also visualized in the network analysis.
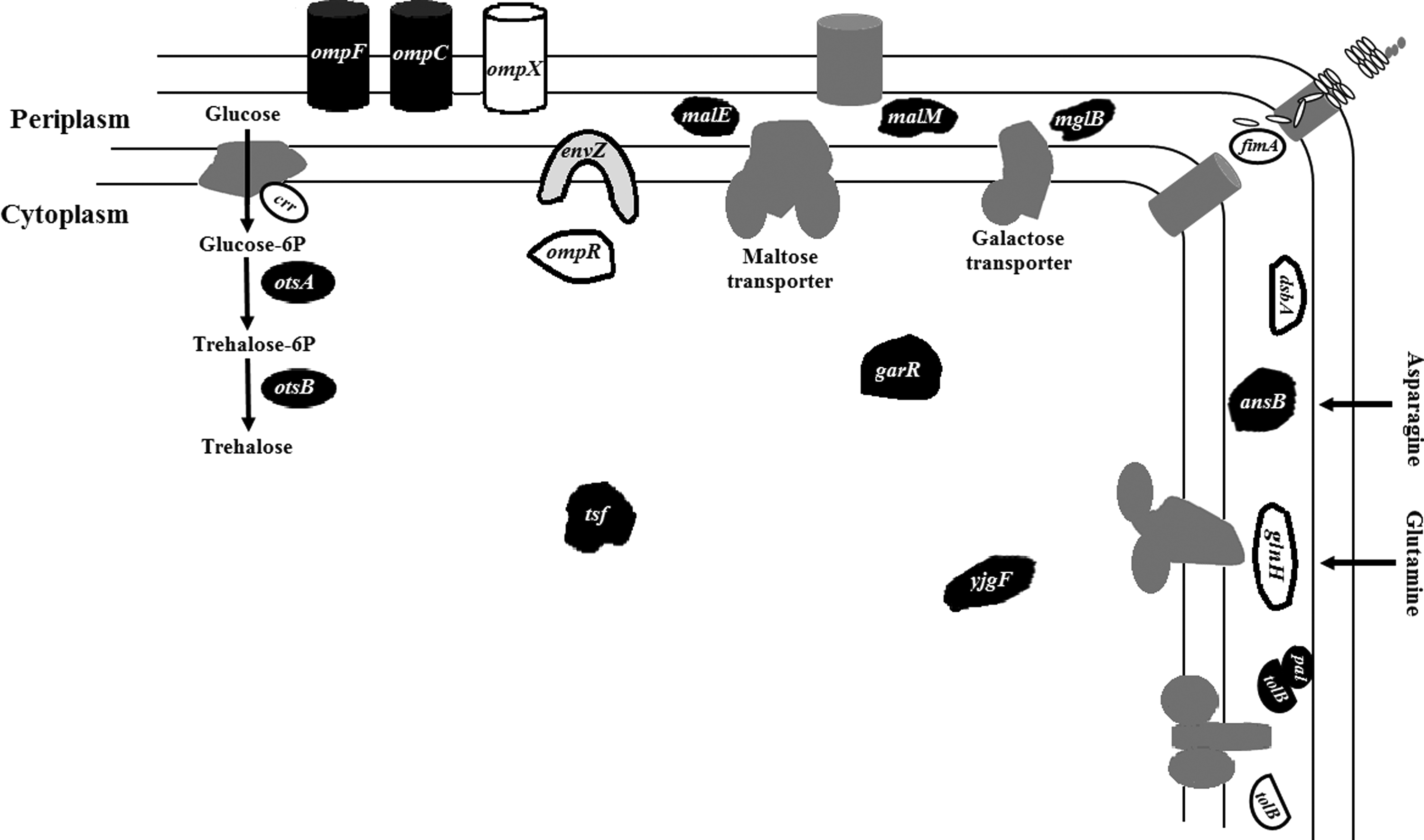
A schematic summary of affected gene products following 1-hour (-)-roemerine treatment. Downregulated gene products are shown in black, upregulated gene products are shown in white, and the unaffected gene product is shown in light gray with a black border. Dark gray components symbolize the overall systems.
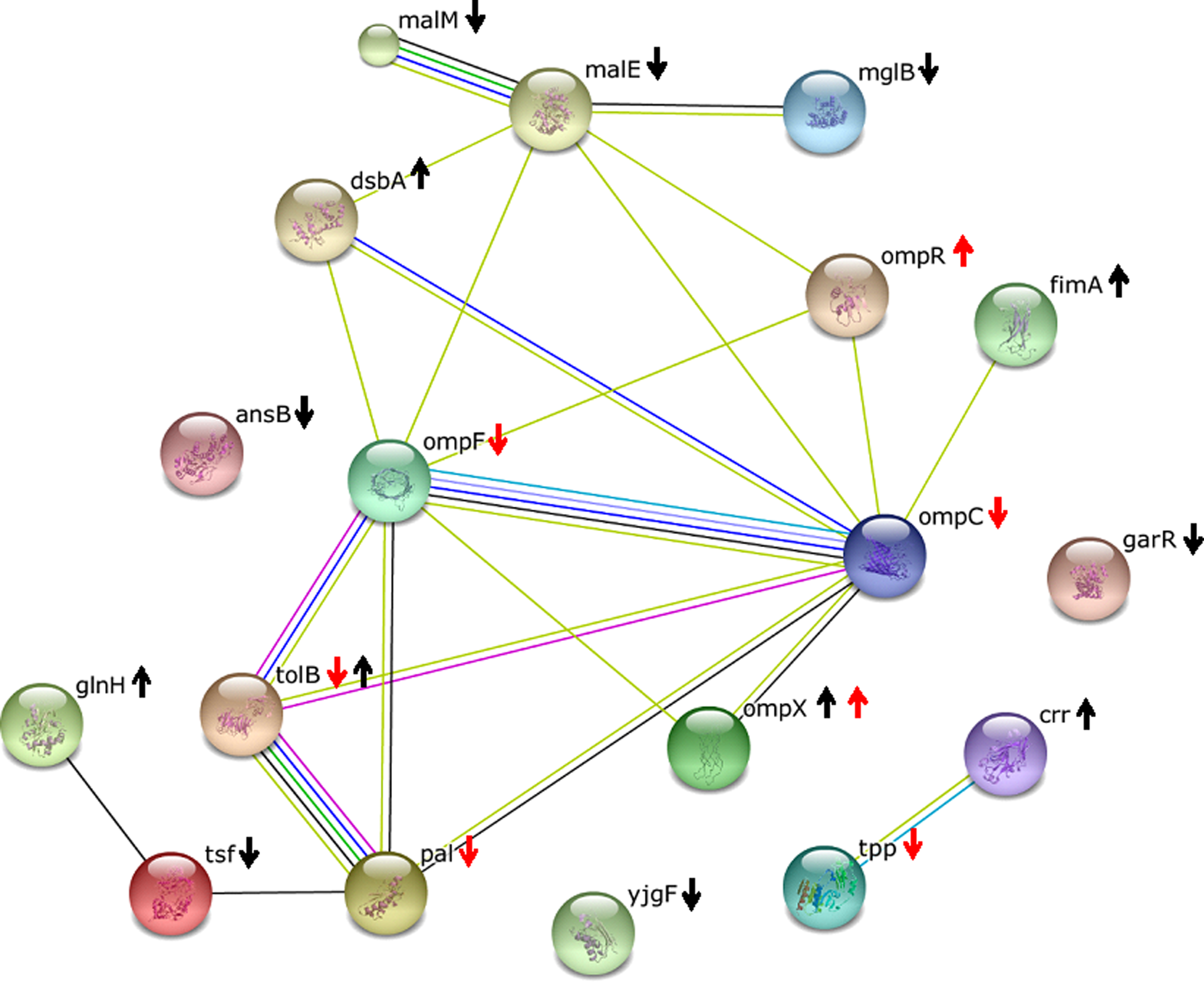
The functional interaction network of proteins in E. coli TB1 following 1-hour treatment with (-)-roemerine. Line colors indicate the methods used for predicting the interaction: black (coexpression), yellow (text mining), blue (databases), green (neighborhood), light blue (homology), red (gene fusion), deep blue (co-occurrence), and purple (experimental results). Black arrows show findings of proteomics analysis; red arrows show findings of qPCR.
Discussion
Omics-guided drug discovery efforts have substantially increased the pace in screening medicinal plants for active compounds (Lin et al., 2012; Reddy et al., 2015; Sahu et al., 2014). In this sense, proteomics is among the most attractive techniques since it is a rigorous method that allows identification of drug-targets and elucidation of action mechanisms of drug candidates. Furthermore, it reflects the dynamic response of the cells upon exposure to the drug candidates (Bandow et al., 2003; Brötz-Oesterheld et al., 2005). In this technology, qPCR is usually coupled to proteomic tools for data validation (Rao et al., 2014; Reddy et al., 2015). The robustness of the method in elucidating drug mechanisms revealed that proteomic tools are not restricted to the investigation of antibacterial activities but can effectively be used in the screening and analysis of other biological activities (Binita et al., 2014; Donnelly and Ciborowski, 2015; Hoehamer et al., 2010; Petri 2003).
The isolation of (-)-roemerine from medicinal plants dates back to 1939 (Marion and Grassie, 1944); interestingly, studies regarding its activity are only few. Present information regarding its activities hints that it possesses different biological activities in different cells. Liou et al. (1979) was the first to demonstrate the antimicrobial activity of (-)-romerine found in Formosan plants. They have reported that Escherichia coli, Bacillus subtilis, and Salmonella typhimurium were among the microorganisms that were affected by this alkaloid. Later, Deng et al. (2006) showed that (l)-roemerine had inhibitory effects against plant pathogenic fungi and bacteria. Then in 2009, Rao et al. (2009) reported that (-)-roemerine was active against Candida species.
The interaction of roemerine with P-glycoprotein (also known as multidrug resistance protein 1), reported by You et al. (1995), could constitute an important finding towards its utilization as an antimicrobial agent. P-glycoprotein is an important eukaryotic membrane protein that functions to pump out foreign substances, including drug molecules. Its inhibition by (-)-roemerine has been reported to reverse multidrug resistance and increase the cytotoxicity of the chemotherapeutic agent, vinblastine, in various cell lines normally insensitive to this drug (You et al., 1995).
Protein tyrosine phosphatase (PTP), CD45, has been reported to be another important target of this alkaloid (Miski et al., 1995). PTPs, which regulate many vital cellular processes, constitute important drug targets since they are found to be involved in unfavorable conditions and disease states (Heneberg et al., 2009). In this respect, search for PTP inhibitors poses an important topic in drug discovery. Miski et al. (1995) was the first to show that (-)-roemerine from Rolinia ulei was among the most potent aporphine alkaloids to inhibit CD45 with an IC50 of 107 μM.
Furthermore, Tran et al. (2010) demonstrated that (-)-roemerine passes ‘rule of five’ (Lipinski et al., 2001) and the two requirements proposed by Veber et al. (2002) for oral bioavailability of drug-like molecules. Unfortunately it shows cytotoxicity to some extent, which decreases in its N-oxide derivatives (Tran et al., 2010). It has also shown to possess mutagenicity (Nozaka et al., 1990) and clastogenicity (Tadaki et al., 1992) for some cell lines but their mechanisms need clarification.
Despite an early report on the antibacterial activity of (-)-roemerine (Liou et al., 1979), information regarding the dynamic response of the cells to (-)-roemerine that leads to retardation in cell growth has not been solved yet. In this work, investigation of the changes in protein abundances, complemented with changes in gene expression levels upon treatment with (-)-roemerine altogether enabled the evaluation of two hypotheses proposed regarding the activity of (-)-roemerine in E. coli TB1. Does this alkaloid change the osmolarity of the growth medium, or does it act by altering the integrity of the cell membranes?
Maintenance of water balance, osmoregulation, is a fundamental requirement for many cells. In the absence of bacterial osmoregulation, under high osmolarity cells tend to lose water, whereas under low osmolarity the cytoplasmic water content increases. In either case, homeostasis is upset and cells eventually die (Wood, 2011). Therefore, osmotic balance should be finely regulated. Under high osmolarity, an increase in the abundance of OmpX in E. coli (Dupont et al., 2007) and Enterobacter aerogenes (Dupont et al., 2004), as well as the repression of maltose regulon in the absence of its external inducer (Bukau et al., 1986) has been reported. With this information we hypothesized that the presence of (-)-roemerine could lead to the disturbance of homeostasis through increasing medium osmolarity, since the abundance of OmpX increased and the abundances of maltose transport system components, MalE and MalM, decreased with (-)-roemerine treatment.
To test this hypothesis, the transcription of the two-component system of E. coli, EnvZ/OmpR, was investigated. The EnvZ/OmpR regulatory system is responsible for sensing changing osmolarity in E. coli. EnvZ is the osmosensor protein that phosphorylates its response regulator OmpR, which reciprocally activates OmpF and OmpC. At high osmolarity, OmpR represses OmpF and induces OmpC (Forst et al., 1989). In addition, the genes involved in trehalose synthesis were analyzed since, at high osmolarity, trehalose is internally synthesized as an osmoprotectant through the activity of otsA and otsB genes (Hengge-Aronis et al., 1991).
Our findings suggested that high osmolarity should not be the reason for retardation in cell growth under (-)-roemerine treatment. Since envZ expression remained unchanged, the EnvZ/OmpR regulatory system was not active to reciprocally regulate ompF and ompC expressions. On the contrary, the expression of both outer membrane porins decreased. The increase found in the transcription of ompR should be independent of the transcription of envZ. Furthermore, trehalose was not synthesized, at all, as an osmoprotectant.
Cell membranes provide structural integrity, protect bacteria against harsh environments, and fulfill a number of tasks that are crucial to the bacterial cell, such as solute and protein transport, with the help of embedded proteins (Koebnik et al., 2000). It is for these reasons that they constitute an important target for antimicrobial agents. The variation in the abundances of OmpX, type-1 fimbriae, and TolB implied a reorganization in the structure of the membrane.
The transcriptional changes found for ompX, ompF, and ompC supported this hypothesis. Membrane modifications under (-)-roemerine treatment should not be unexpected since the porins encoded by ompC and ompF are entry routes of different antibacterials into E. coli cells (Mahendran et al., 2010; Tran et al., 2014) and the gene encoded by ompX interferes with the regulation of cell surface structures in E. coli cells (Otto and Hermansson, 2004). Dupont et al. (2004) reported decreased vulnerability to β-lactams with increased ompX expression. They also demonstrated that ompX overexpression was accompanied with ompF reduction upon exposure to toxic compounds. This type of regulation in OmpX and OmpF was suggested to be an initial response to stress conditions to change the permeability of the membrane (Dupont et al., 2007). In an early report by Stoorvogel et al. (1987), cloning of ompX in a multicopy plasmid in E. coli resulted in a decrease in the level of OmpF and OmpC proteins.
More recently, Karaosmanoglu et al. (2014) reported a reduction in the abundance of OmpC under berberine-chloride treatment. Thus, it was suggested that a change in membrane permeability should be a result of antibiotic resistance development. Given this information, we hypothesized that the upregulation of OmpX and the downregulation of OmpF and OmpC would be a result of modulation of the membrane permeability to impede (-)-roemerine entry.
We believe that the limitation in the permeability of the membrane through downregulation of the outer membrane porins, OmpF and OmpC, has led to a shortfall in intracellular carbon availability. The decreases in the abundance of proteins belonging maltose (MalE, MalM) and galactose transport (MglB) also support this notion. This condition could force cells to alternative carbon sources.
Alternatively, this finding could imply a shift in the cell physiology towards a state of metabolic shutdown. The increase in the PTS component of glucose, EIIAGlc (crr), a key regulator protein in carbon metabolism, hints at an internal struggle in this respect. The decrease in the abundance of MalE could be a “proteomic signature” for the presence of alkaloids, since it has also been reported with E. coli cells exposed to the isoquinoline alkaloid berberine (Karaosmanoglu et al., 2014; Ozbalci et al., 2010; Van Bogelen et al., 1999).
The small outer-membrane OmpX, was reported to possess similarity to outer-membrane proteins involved in virulence (Mecsas et al., 1995). The oligomeric protein, Type-1 fimbriae (fimA), is attached to bacterial outer membrane and mediates bacterial adhesion to the surfaces. Since it enables adhesion, it acts in pathogenicity (Beachey et al., 1988). Interestingly, a reciprocal regulation has been reported between OmpX and type-1 fimbrial protein. During virulence, adhesion-induced phenotypic changes in type 1 fimbriated E. coli accompany reductions in OmpX (Otto and Hermansson, 2004). Although simultaneous regulation of OmpX and fimA brings pathogenicity to mind, the simultaneous increase in their abundances indicates a requirement for attribution of different roles to both of these proteins under (-)-roemerine treatment. For example, diverse functions, such as the interference of OmpX in the formation of cell surface structures and the bacterial attachment to surfaces through the fimbrial protein to obtain nutrients, strengthen the idea that these two proteins could have unrelated functions.
Disulfide bond formation in FimA has crucial importance in fimbriae assembly. Crespo et al. (2012) reported that this process in Type-1 fimbriae (fimA) was catalyzed by the periplasmic protein DsbA. Moreover, dsbA gene deletion in E. coli rendered the disappearance of Type-1 fimbriae (Bringer et al., 2007; Heras et al., 2009). The increased abundance in DsbA, along with Type-1 fimbriae, correlates well with the correct assembly of the fimbriae. DsbA can also contribute to correct protein folding by catalyzing disulfide bond formation in different polypeptide chains for the dynamic folding of membrane bound and surface exposed proteins (Denoncin and Collet, 2013; Vivian et al., 2009).
TolB is a component of the Tol system, which acts in outer membrane integrity, and interacts with peptidoglycan-associated lipoprotein (PAL). Tol mutants of E. coli were found to be highly sensitive to antibiotics (Bouveret et al., 1995). The increase in the abundance of TolB in response to (-)-roemerine treatment gave an impression that the integrity of the outer membrane was affected. Hence tolB and pal genes were selected for transcriptional analysis. Although the abundance of TolB increased, transcriptional analysis of tolB and pal showed a reduced expression for both genes. The conflicting result for TolB requires further analysis.
The decrease in the abundances of L-asparaginase II (ansB), a protein in asparagine metabolism and yjgF, a protein reported to be related to isoleucine biosynthesis in Salmonella enterica (Christopherson et al., 2008) clued that the presence of (-)-roemerine has also declined the protein levels involved in amino acid biosynthesis and metabolism. This deficiency in intracellular amino acid supply in turn triggered glutamine uptake by increasing the abundance of glutamine binding protein, encoded by glnH. This amino acid is essential in the biosynthesis of both proteins and nitrogen-containing compounds in bacteria (Forchhammer, 2007).
The minimization in the abundances of proteins involved in amino acid biosynthesis and metabolism impaired protein biosynthesis. This is reflected by the low abundance in the highly conserved protein Elongation Factor-Translation, stable (EF-Ts), which acts in the elongation cycle in protein biosynthesis. The observed decline in protein levels involved in biosynthesis and metabolism could be related to the state of metabolic shutdown.
Conclusions
Limitation in the permeability of the outer membrane and the reduction of transport proteins of carbohydrate utilization in the presence of (-)-roemerine resulted in poor carbon source availability. The shortfall of intracellular carbon sources in turn led to impaired cell growth. The decrease in the abundances of proteins related to translational machinery, and amino acid biosynthesis and metabolism was accompanied by a nutrient-limited state. This finding could suggest a metabolic shutdown in E. coli cells. High osmolarity was clearly not one of the reasons of bacterial death by (-)-roemerine. These observations collectively attest to the promise of plant omics and profiling of putative drug candidates using proteomics tools. Omics-guided drug discovery deserves greater attention in mainstream pharmacology, so as to better understand the plants' medicinal potentials.
Footnotes
Acknowledgments
We would like to thank Bulent Mertoglu, Ayse Ogan, and Ahu Altınkut Uncuoğlu for their help. Funding by TUBITAK (113M052) and Marmara University, Scientific Research Projects Committee (FEN-A-050614-0238) are gratefully acknowledged. NBG was supported by TUBITAK-BIDEB 2211 fellowship.
Author Disclosure Statement
The authors declare that there are no conflicting financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.