
Abstract
Abstract
Mammalian milk is a key source of lipids, providing not only important calories but also essential fatty acids. Veterinary medicine and omics systems sciences intersection, termed as “veterinomics” here, has received little attention to date but stands to offer much promise for building bridges between human and animal health. We determined the changes in porcine mammary genes and proteomics expression associated with milk triacylglycerol (TAG) synthesis and secretion from late pregnancy to lactation. TAG content and fatty acid (FA) composition were determined in porcine colostrum (the 1st day of lactation) and milk (the 17th day of lactation). The mammary transcriptome for 70 genes and 13 proteins involved in TAG synthesis and secretion from six sows, each at d −17(late pregnancy), d 1(early lactation), and d 17 (peak lactation) relative to parturition were analyzed using quantitative real-time PCR and Western blot analyses. The TAG content and the concentrations of de novo synthesized FAs, saturated FAs, and monounsaturated FAs were higher in milk than in colostrum (p < 0.05). Robust upregulation with high relative mRNA abundance was evident during lactation for genes associated with FA uptake (VLDLR, LPL, CD36), FA activation (ACSS2, ACSL3), and intracellar transport (FABP3), de novo FA synthesis (ACACA, FASN), FA elongation (ELOVL1), FA desaturation (SCD, FADS1), TAG synthesis (GPAM, AGPAT1, LPIN1, DGAT1), lipid droplet formation (BTN2A1, XDH, PLIN2), and transcription factors and nuclear receptors (SREBP1, SCAP, INSIG1/2). In conclusion, a wide variety of lipogenic genes and proteins regulate the channeling of FAs towards milk TAG synthesis and secretion in porcine mammary gland tissue. These findings inform future omics strategies to increase milk fat production and lipid profile and attest to the rise of both veterinomics and lipidomics in postgenomics life sciences.
Introduction
L
The FAs in milk TAG are primarily long chain FAs (LCFAs) with 16 to 22 carbon atoms, while a small amount of medium chain FAs (MCFAs) with 8 to 14 carbon atoms are also present. Palmitic acid, oleic acid, and linoleic acid are the most abundant FAs found in mammalian milk TAG (Laws et al., 2009). The FAs that are used in mammary cell milk TAG synthesis have two main origins: 1) they are synthesized de novo in the mammary gland; 2) they are released into the circulation from chylomicrons by lipoprotein lipase (LPL) and from adipose tissue by hormone-sensitive lipase (HSL) (Scow et al., 1977).
Milk lipid synthesis and secretion in the lactating mammary gland is a complex process that requires the coordinated activity of a wide variety of pathways. These pathways involve many key proteins related to FA uptake (VLDLR, LPL, CD36, and SLC27A), FA activation and intracellar transport (ACSS, ACSL, FABP, and ACBP), FA synthesis (ACACA and FASN), and desaturation (SCD, ELOVL, and FADS), TAG synthesis (GPAM, AGPAT, LPIN, and DGAT), and lipid droplet formation (BTN, XDH, and PLIN) (Bionaz and Loor, 2008a; 2008b; Mohammad and Haymond, 2013; Rudolph et al., 2007; Yadav et al., 2015; Zhu et al., 2015).
Additionally, milk lipid synthesis is regulated by various transcription factors and nuclear receptors, including SREBPs, SCAP, INSIGs, THRSP, PPARs, and LXR (Bionaz and Loor, 2008b; Kadegowda et al., 2009; Li et al., 2015; Ma and Corl, 2012; Mohammad and Haymond, 2013; Oppi-Williams et al., 2013; Shi et al., 2014). It can be assumed that coordinated transcriptional regulation occurs during mammary gland lipid synthesis; however, the role of these transcriptional factors in lactating sow milk lipid production remains unclear.
We hypothesize that the genes and proteins involved in sow milk TAG synthesis may be regulated in a coordinated fashion during the period of physiological transition from late pregnancy to lactation. Therefore, the aim of this study was to use quantitative real-time PCR (qPCR) and Western blot analysis to establish the developmental pattern of key factors that participate in milk TAG synthesis and secretion in porcine mammary tissue from late pregnancy to peak lactation.
Materials and Methods
Animals and tissue preparation
Large White sows (n = 6, four to six parities) were purchased from the Huizhou Swine Breeding Center in South China and were managed by standard procedures. The diets (Supplementary Table S1; supplementary material is available online at www.liebertpub.com/omi) were corn and soybean meal-based and were formulated to meet or exceed nutrient concentrations recommended by the National Research Council (NRC, 1998).
Percutaneous biopsies were excised from the right or left anterior mammary gland of each sow at day −17 (± 2) (late pregnancy), day 1 (early lactation), and day 17 (± 2) (peak lactation) relative to parturition. Approximately 0.8 g of mammary tissue were excised by the surgical method described in Kirkwood et al. (2007). All procedures were conducted under protocols that had been approved by the Committee for the Care and Use of Experimental Animals at South China Agricultural University.
Measurement of the TAG content in mammary tissue
The TAG in porcine mammary tissue was determined by enzymatic colorimetric methods using commercial kits (Beijing Applygene Technology Co., Ltd., Beijing, China). Protein concentrations were determined using a Pierce BCA protein Assay kit (Thermo Scientific, IL, USA), and the TAG contents were normalized for protein in each well. Each experiment was performed in triplicate and repeated a minimum of three times.
Milk sampling and FA analysis
Colostrum samples were collected on the day of parturition, within 4 h of the birth of the first piglet. Milk samples were collected on d 17 post-farrowing following the intra-muscular administration of 2 mL oxytocin (10 IU/mL; NVS, UK). The samples were stored without preservative at −20°C until analyzed for FA composition. Briefly, the colostrum and milk lipids were extracted and transesterified as described previously (Christopherson and Glass, 1969). FA methyl esters were measured using a gas chromatograph (Agilent 6890N), and FA peaks were identified and quantified by using methyl ester standards (37-component FAME Mix, Sigma).
RNA extraction and real-time PCR
Total RNA was isolated from 100 mg tissue samples using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instructions. First strand cDNA synthesis was performed by using a PrimeScript RT reagent kit with gDNA eraser (Takara, Dalian, China). qPCR was performed in an ABI Prism 7500 Sequence Detection System using SYBR Green Real-Time PCR Master Mix (Toyobo, Osaka, Japan) in a final volume of 20 μL. The abundance of target gene and reference gene (GAPDH, MRPL39 and β-actin) mRNA transcripts was determined by qPCR. All the primer sequences for qPCR are shown in Supplementary Table S2 and additional information on the target genes is provided in Supplementary Table S3.
All samples mRNA levels were normalized by subtracting from each of the target gene cycle thresholds (Ct) the arithmetic means of the three selected reference gene Ct, and the results are expressed as the changes in the Ct value relative to d −17 using the 2-ΔΔCt method (Livak and Schmittgen, 2001). To estimate standard errors at d −17 and prevent biases in statistical analysis, normalized data were transformed to obtain a perfect mean of 1.0 at d −17, leaving the proportional difference between the biological replicates. The same proportional change was calculated at all other time points to obtain the change relative to d −17. PCR amplification efficiency and the mRNA abundance for each gene were calculated as described in detail previously (Bionaz and Loor, 2008a; de Jong et al., 2007). The overall gene abundance for porcine genes at each time point was calculated using the average value of ΔCt.
Western blot analysis
Mammary tissue samples were homogenized in RIPA lysis buffer (Beyotime, Nanjing, China). The homogenates were combined with equal volumes of SDS sample buffer, and the proteins were separated by electrophoresis on a 5%–12% polyacrylamide gel and transferred to nitrocellulose membranes. The membranes were blocked with 5% skim milk in Tris-buffered saline with Tween, followed by overnight probing with antibodies. β-Actin was intended to serve as a loading (internal) control. After washing, membranes were incubated with secondary antibody (Amersham Biosciences). The chemiluminescent signal was detected by using ECL reagents (Beyotime, Nanjing, China), and bands were quantified by Image Processing Software (Image Pro Plus6.0).
Statistical analysis
Comparisons between the time points were performed using the Wilcoxon Rank Sum tests analysis for categorical parameters and Tukey's test analysis for continuous variables. Bonferonni corrections were evaluated following bivariate analysis. Experimental data were tested for normality of distribution using the Shapiro-Wilk's test. Log transformations were performed on non-normative variables and then analyzed by one-way ANOVA followed by a Tukey's honestly significant difference test. Differences at p < 0.05 were considered statistically significant. Values are expressed as means ± SEMs.
Results
TAG content in mammary tissue
The TAG content in the porcine mammary tissue increased (two to three)-fold from late pregnancy (60.4 μg/mg) to early (195.8 μg/mg) and peak lactation (214.6 μg/mg protein), and the TAG content of milk was significantly increased (p < 0.05) relative to colostrum, indicating the developmental transition of TAG production from late pregnancy to peak lactation (Fig. 1A and B).
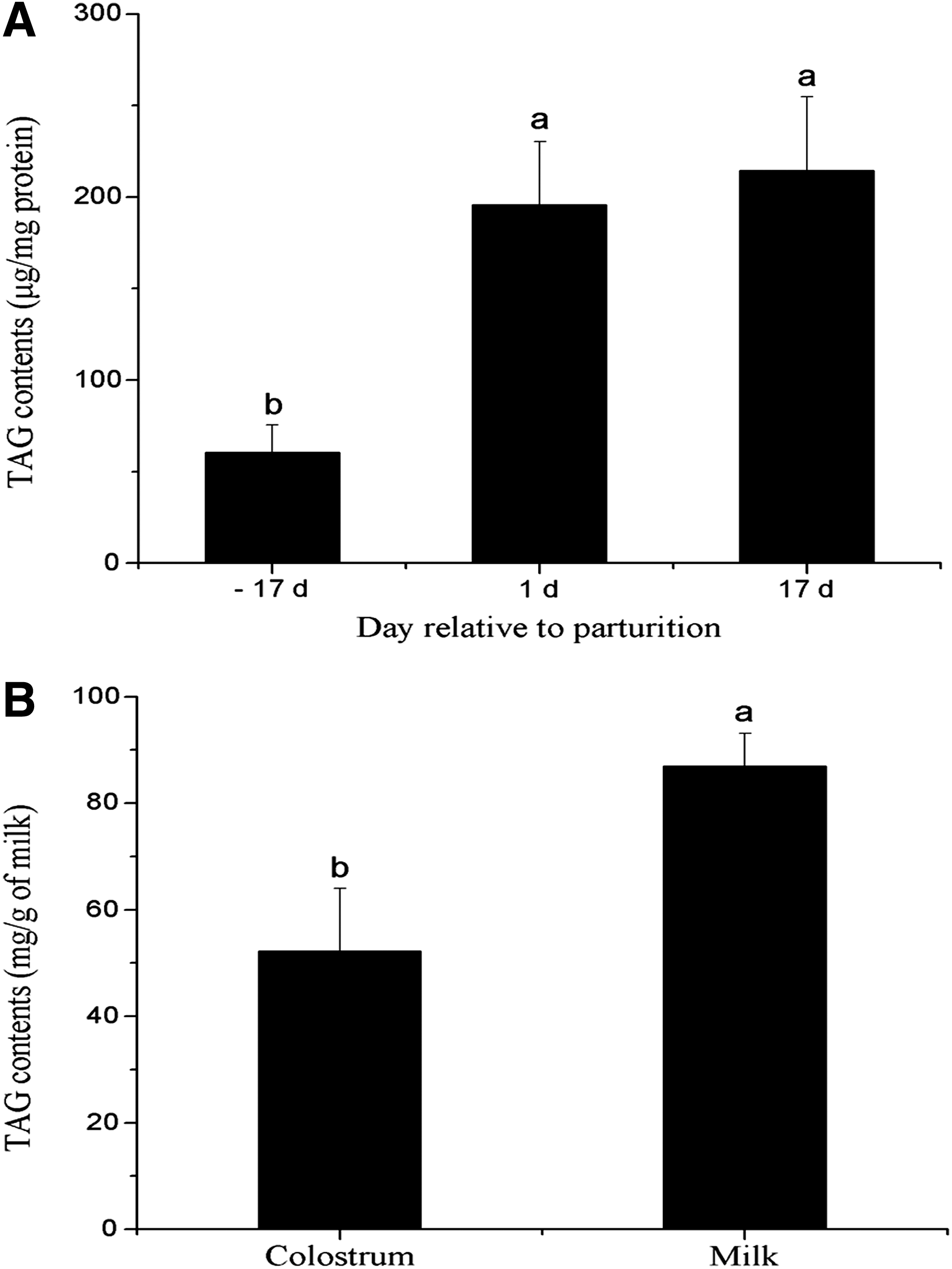
Concentration of TAG in the porcine mammary tissue
FA composition in milk
Total FAs were elevated in the sow milk relative to the sow colostrum (p < 0.05) (77.50 ± 4.73 vs. 47.34 ± 5.60, respectively) (Table 1). The C16:0, C18:1, C18:2(n-6) were the highest FAs concentrations in both colostrum and milk (Table 1). The concentrations of C12:0, C14:0, C14:1, C16:0, C16:1, C17:1, C24:1, C22:6(n-3) were higher in milk than in colostrum, but the concentration of C20:3(n-6) was lower in milk than in colostrums. No other differences were observed in the concentration of other FAs between colostrum and milk (Table 1). The concentrations of de novo synthesized FAs (DNSFAs), saturated FAs (SFAs), and monounsaturated FAs (MUFAs) were higher in milk than in colostrum (p < 0.05) (Table 1).
Values are means ± SEM, n = 6; 2DNSFA = sum of de novo synthesized fatty acids (FAs <16 carbons); 3SFA = sum of saturated fatty acids; 4MUFA = sum of monounsaturated fatty acids; 5n-3 PUFA = sum of n-3 polyunsaturated fatty acids; 6n-6 PUFA = sum of n-6 polyunsaturated fatty acids; 7PUFA = sum of polyunsaturated fatty acids; 8VLCFA = sum of very-long chain fatty acids (≥ 20 carbons).
Expression of genes or proteins related to FA uptake in lactating porcine mammary gland
The mRNA expression of VLDLR, LPL, CD36, SLC27A3, and SLC27A4 was upregulated throughout lactation and increased significantly to peak values at d 17 (1.8-fold, 24-fold, 26-fold, 2.6-fold, and 1.9-fold at d 17 vs. d −17, respectively, p < 0.05, Fig. 2A and B). Expression of the gene for CD36 is the highest in mRNA abundance and the degree of change within this category of genes (Table 2).
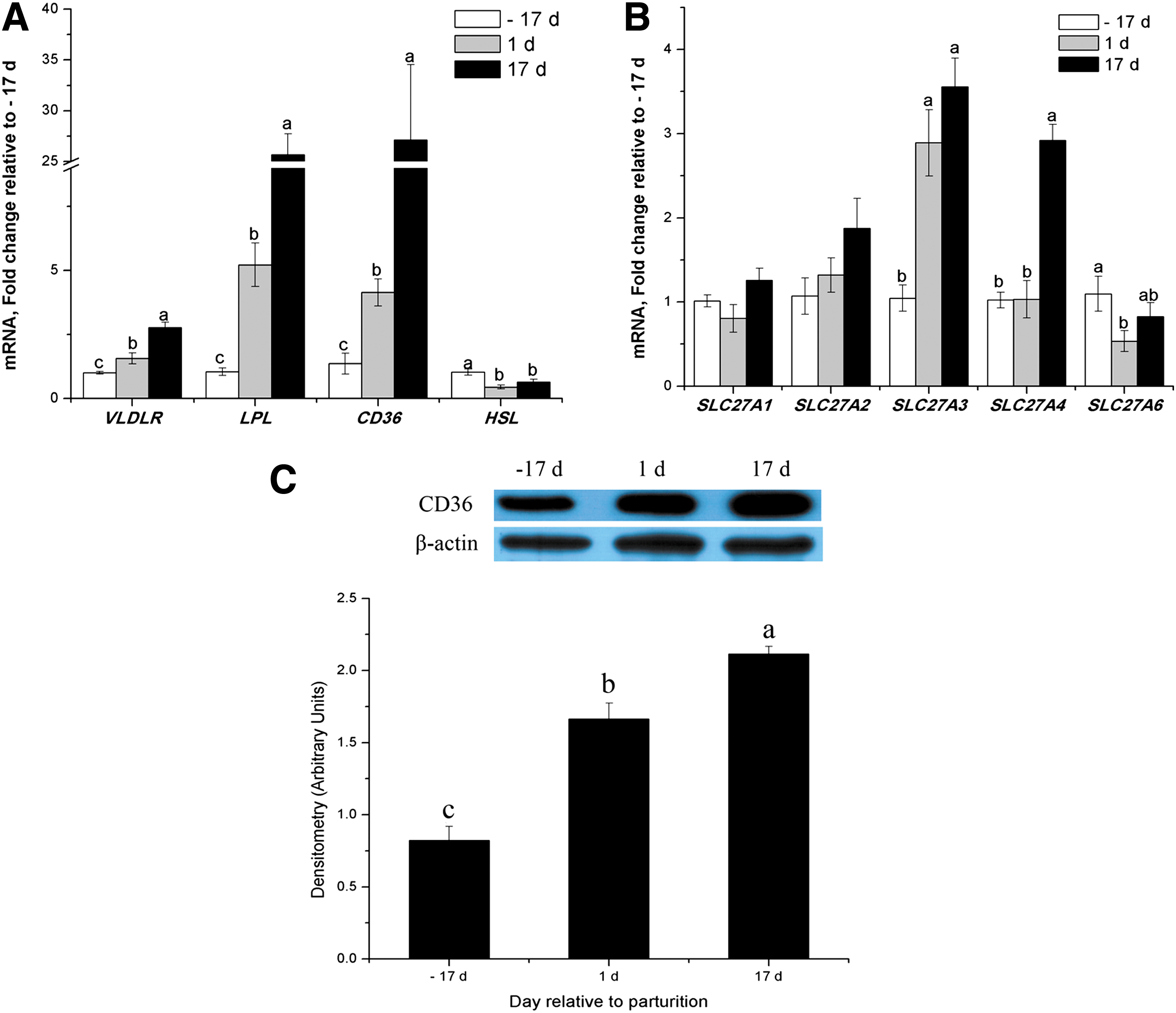
Expression of genes or proteins for FA uptake in porcine mammary tissue throughout the lactation cycle.
The % mRNA abundance among all genes investigated in the study was calculated as [((1/qPCR EfficiencyΔCt) specific gene/sum (1/qPCR EfficiencyΔCt) all genes measured) ×100].
Similarly, CD36 protein expression in porcine mammary tissue significantly increased throughout lactation (Fig. 2C). However, mRNA abundance for HSL and SLC27A6 were lower by 55% and 47%, respectively, during the transition from late pregnancy to lactation (p < 0.05, Fig. 2A and B). There were no differences in the mRNA abundance for SLC27A1 or SLC27A2 (Fig. 2B) between late pregnancy and lactation. During lactation, SLC27A3 mRNA was most abundant because it accounted for >70% of all SLC27A isoforms (Table 3). Other SLC27A isoforms had lower mRNA abundance but mRNA of SLC27A5 was undetectable during lactation (Table 3).
At each time, mRNA abundance of each isoform in the gene family was calculated as [((1/qPCR EfficiencyΔCt) specific gene/sum (1/qPCR EfficiencyΔCt) all genes of specific gene family) ×100], where ΔCt = median Ct gene – median Ct internal controls (Bionaz and Loor, 2008; de Jong et al., 2007).
Undetectable.
Expression of genes or proteins related to FA activation and intracellar transport in lactating porcine mammary gland
From late pregnancy to peak lactation, the transcript abundance of genes increased 4-fold for ACSS1, 8-fold for ACSS2, 3-fold for ACSS3, 59-fold for ACSL3, > 700-fold for ACSL6, 2.3-fold for FABP1, 9-fold for FABP3, and 2.3-fold for FABP5 (p < 0.05, Fig. 3A–C). Western blot analysis showed that FABP3 markedly increased during lactation compared with late pregnancy (p < 0.05, Fig. 3D). However, mRNA abundance for ACSL5 and FABP6 was 47% and 63% lower, respectively, for porcine mammary glands at peak lactation compared with late pregnancy (P < 0.05, Fig. 3B and C).
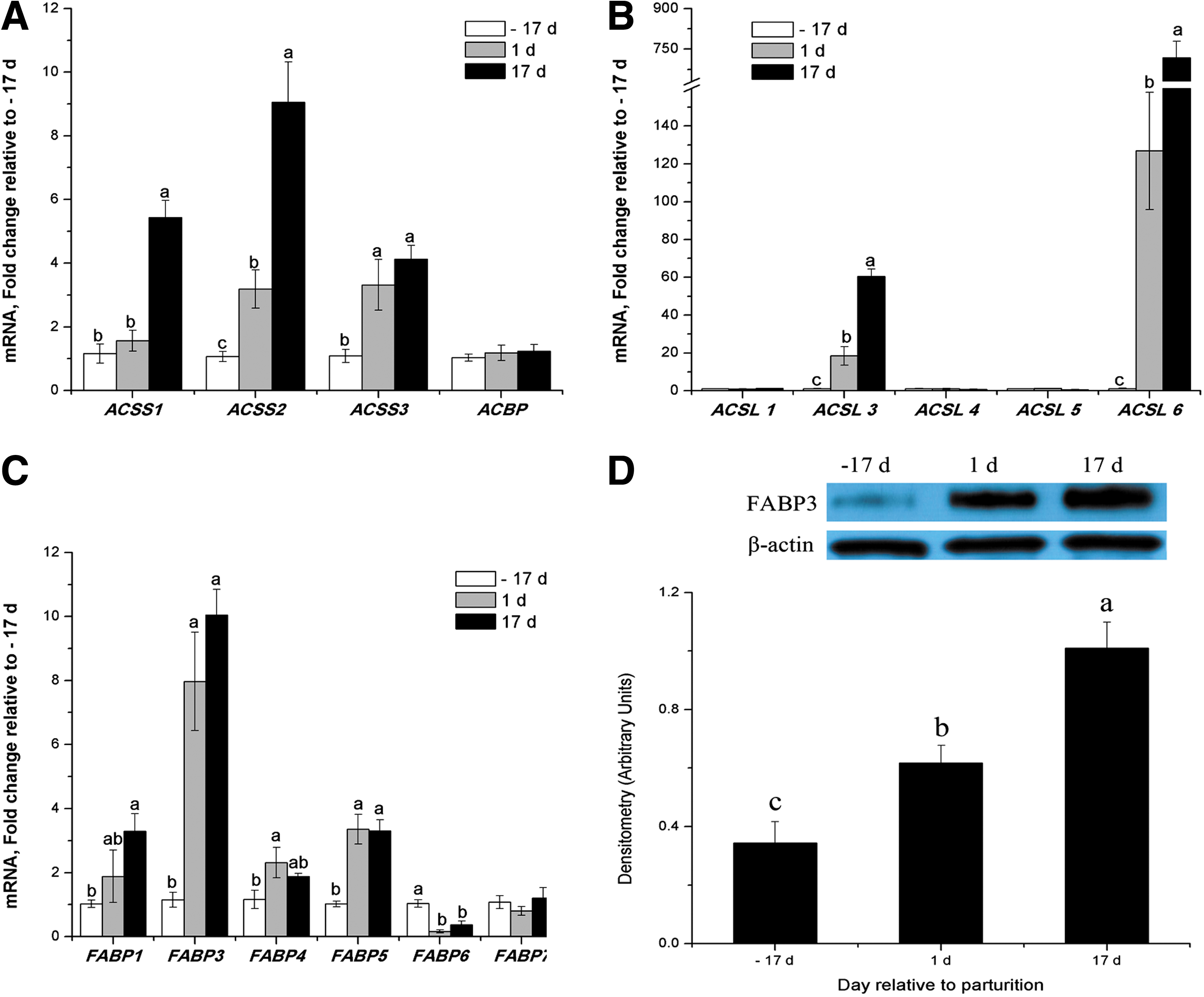
Expression of genes or proteins related to FA activation and intracellar transport in porcine mammary tissue throughout the lactation cycle.
There were no differences in mRNA abundance for ACSL1, ACSL4, FABP7, and ACBP (Fig. 3A-C). ACSS2, ACSL3, and FABP3 are the major isoforms within each gene family in porcine mammary tissue (Table 3). Other ACSS, ACSL isoforms and FABP4, FABP5 exhibited lower mRNA abundance, while FABP1, FABP6 and FABP7 were barely detectable, and FABP2 was undetectable (Table 3).
Expression of genes or proteins related to FA synthesis and desaturation in lactating porcine mammary gland
ACACA, FASN, and SCD were markedly upregulated throughout lactation at both the mRNA and protein levels (p < 0.05, Fig. 4A, C–E). The mRNA expressions of FADS3, ELOVL1, ELOVL5, and ELOVL6 were upregulated throughout lactation and increased 1.4-fold, 6-fold, 1.5-fold, and 3.8-fold, respectively (p < 0.05, Fig. 4A and B). FADS1 mRNA expression was only increased significantly at early lactation (p < 0.05, Fig. 4A).
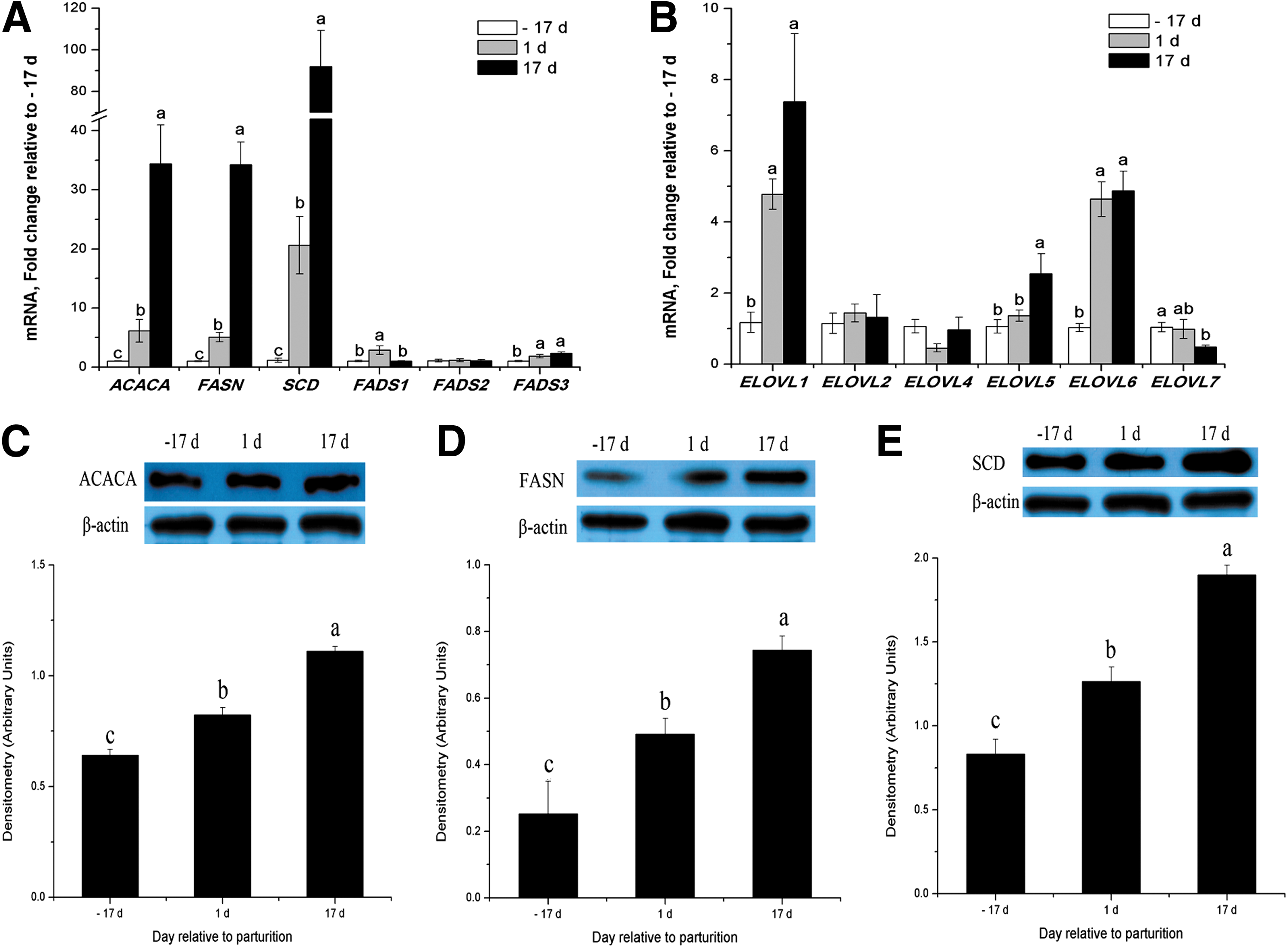
Expression of genes or proteins related to FA synthesis and desaturation in porcine mammary tissue throughout the lactation cycle.
However, mRNA abundance for ELOVL7 was 56% lower for the porcine mammary gland at peak lactation compared with late pregnancy (p < 0.05, Fig. 4B). There were no differences in the mRNA abundance for ELOVL2, ELOVL4, and FADS2 (Fig. 4A and B). FADS1 and ELOVL1 are the major isoforms within each gene family in porcine mammary tissue (Table 3). Other FADS isoforms and ELOVL5, ELOVL6, and ELOVL7 exhibited lower mRNA abundance, while ELOVL2 and ELOVL4 were barely detectable, and ELOVL3 was undetectable (Table 3).
Expression of genes or proteins related to TAG synthesis in lactating porcine mammary gland
Most of the genes associated with TAG synthesis were upregulated throughout lactation and increased significantly to peak values at d 17, including GPAM (6.6-fold), AGPAT1 (11.4-fold), AGPAT2 (0.9-fold), AGPAT3 (1.8-fold), AGPAT5 (0.6-fold), AGPAT6 (9.8-fold), AGPAT8 (3.2-fold), AGPAT9 (2.5-fold), LPIN1 (6.6-fold), and DGAT1 (4.8-fold) (p < 0.05) (Fig. 5 A-C). Western blot analysis of GPAM, AGPAT1, LPIN1, and DGAT1 proteins in porcine mammary tissue confirmed their increases at peak lactation compared with late pregnancy (Fig. 5D). However, LPIN3 mRNA expression was significantly downregulated (p < 0.05) during lactation (Fig. 5C). There were no differences in mRNA abundance for AGPAT4, AGPAT7, and DGAT2 (Fig. 5A-C).
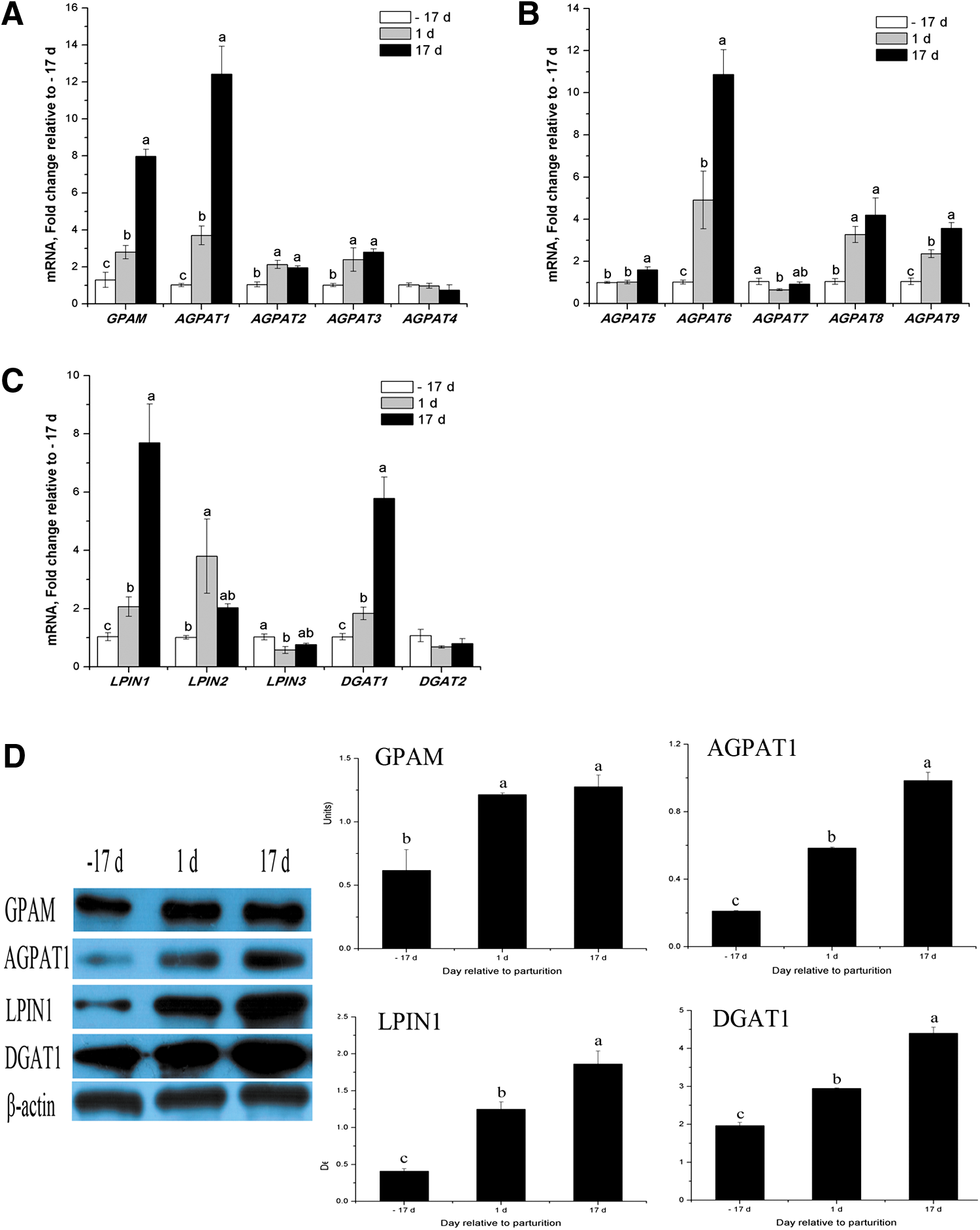
Expression of genes or proteins related to TAG synthesis in porcine mammary tissue throughout the lactation cycle.
The mammary gland expressed all AGPAT isoforms measured, with mRNA for AGPAT1 being predominant (43%–60% of AGPAT isoforms), followed by AGPAT6 (26%–34% of AGPAT isoforms; Table 3). Other AGPAT isoforms had lower mRNA abundance but all were detectable (Table 3). LPIN2 was the most abundant isoform from late pregnancy to early lactation, accounting for 54%–76% of all LPIN mRNA, with LPIN1 being the next most abundant (55%) during peak lactation (Table 3).
Expression of genes or proteins related to lipid droplet formation in lactating porcine mammary gland
Four genes (BTN2A1, XDH, PLIN2, and PLIN3) were upregulated throughout lactation and increased significantly to peak values at d 17 (4.6-fold, 187-fold, 5.4-fold, and 4.5-fold, respectively, p < 0.05, Fig. 6A). Western blot analysis showed that PLIN2 increased only at peak lactation compared with late pregnancy (p < 0.05, Fig. 6B). The mRNA abundances for PLIN1 and PLIN4 were 95% and 80% lower, respectively, for the porcine mammary gland at peak lactation compared with late pregnancy (p < 0.05, Fig. 6A). In the PLIN family, four isoforms were discovered in the porcine mammary gland, with mRNA of PLIN2 being predominant (> 90% of PLIN isoforms) while PLIN1 and PLIN4 were almost undetectable (Table 3).

Expression of genes or proteins related to lipid droplet formation in porcine mammary tissue throughout the lactation cycle.
Expression of genes or proteins related to regulation of transcription in lactating porcine mammary gland
Compared with late pregnancy, mRNA abundance for most of the genes associated with the regulation of transcription were upregulated throughout lactation and increased at peak lactation, including INSIG1 (72-fold), INSIG2 (48-fold), SREBP1 (2.2-fold), SREBP2 (2.2-fold), SCAP (3.4-fold), THRSP (17-fold), PPARβ/δ (5.9-fold), and LXR (2.4-fold) (p < 0.05) (Fig. 7A and B). Western blot analysis showed that SREBP1 increased only at peak lactation, compared to late pregnancy (p < 0.05, Fig. 7C).
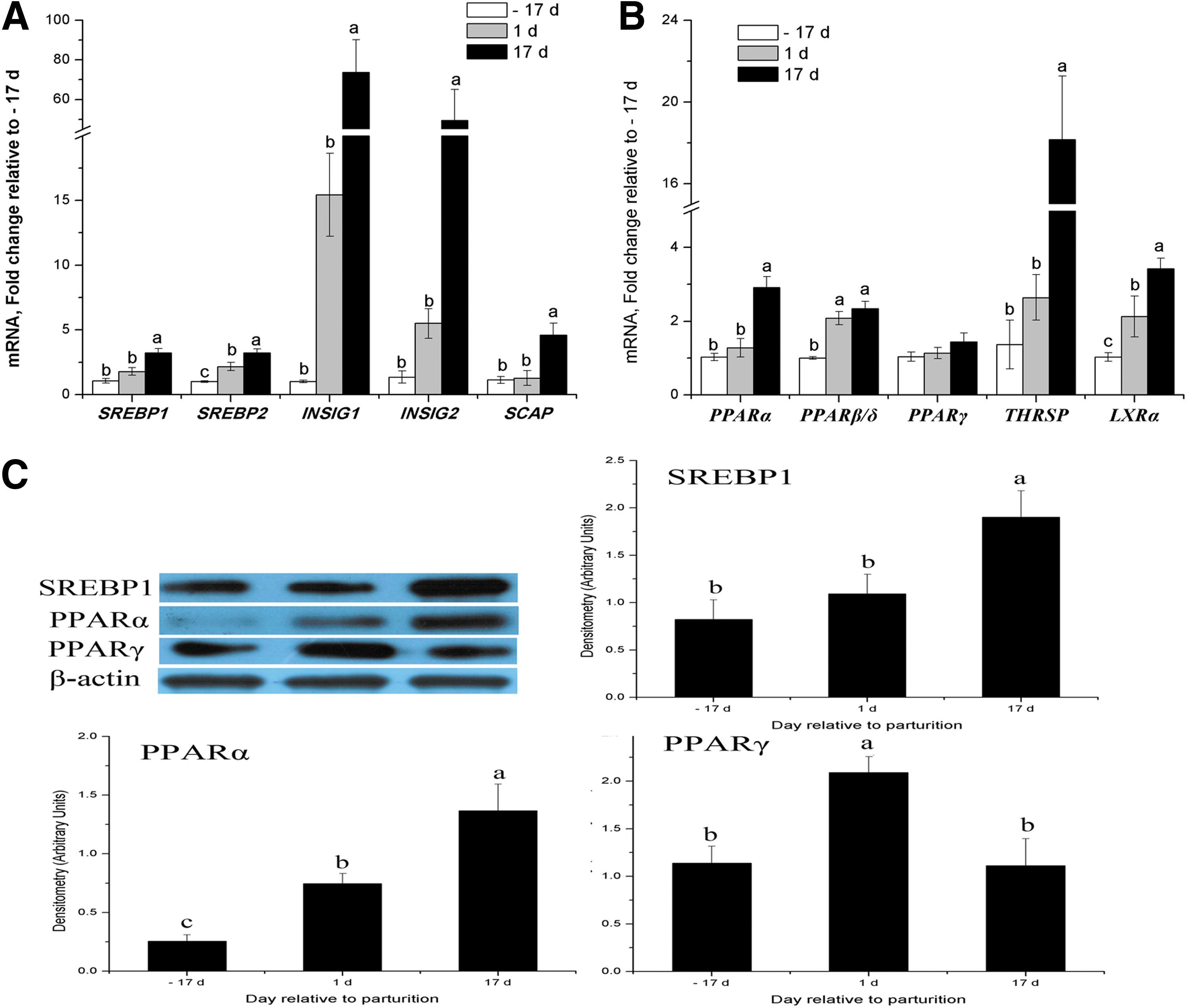
Expression of transcription factors and nuclear receptors at the mRNA and protein levels in porcine mammary tissue throughout the lactation cycle.
PPARα was upregulated both at the gene and protein expression levels from late pregnancy to peak lactation (p < 0.05, Fig. 7B and C). PPARγ mRNA expression was not affected by lactation (Fig. 7B), but its protein expression was increased at early lactation compared with late pregnancy (p < 0.05, Fig. 7C). Overall, total SREBP isoform mRNA abundance represented ∼2.9% of the genes measured (Table 2). SREBP1 was the most abundant isoform, accounting for ∼86% of all SREBP mRNA (Table 3). In the INSIG family, mRNA abundances for INSIG1 and INSIG2 were essentially equivalent in the lactating porcine mammary gland (Table 3).
Discussion
In the present study, we found that the TAG content within porcine mammary tissue increased 2-fold from late pregnancy to early and peak lactation (Fig. 1) and that the concentrations of DNSFAs, SFAs, and MUFAs in TAG was markedly higher in milk than in colostrum (p < 0.05). Enhanced TAG or FA synthesis is likely attributable to upregulation of many key genes or proteins in the mammary gland, including those involved in FA uptake from the blood, intracellular FA activation and transport, de novo FA synthesis, FA elongation and desaturation, TAG synthesis, lipid droplet formation, and transcription regulation. These results suggest a developmental transition in TAG production in sows from late pregnancy to peak lactation.
FA uptake in the mammary gland
The LCFA substrate that is available for milk TAG synthesis in mammary cells is derived from plasma chylomicrons by LPL activity and from adipose tissue by HSL activity (Scow et al., 1977). VLDLR could increase LPL and TAG-rich lipoprotein interactions at the capillary surface and help the transcytosis of LPL (Takahashi et al., 2003). In this study, we confirmed that LPL and VLDLR had higher mRNA abundances as early as the onset of milk synthesis in the lactating porcine mammary gland, similar to what has been observed in bovine (Bionaz and Loor, 2008b) and human models (Mohammad and Haymond, 2013).
SLC27A represents a key family of genes for exogenous LCFA trans-membrane transport and activation (Anderson and Stahl, 2013). Overall, SLC27A isoform mRNA abundance represented 0.1% of all genes measured in this study (Table 2). The expression of SLC27A3 increased to a peak value at d 17 (2.6-fold at d 17 vs. d −17, p < 0.05, Fig. 2B), while other isoforms exhibited smaller degrees of change with low mRNA abundance. Likewise, low SLC27A mRNA abundance with relatively small changes have been demonstrated in bovine (Bionaz and Loor, 2008a; 2008b) and human (Mohammad and Haymond, 2013) mammary gland tissue. The results suggest some involvement of SLC27A3 in the uptake of FAs from the circulating blood FA pool.
Besides SLC27A, CD36 was also the main protein involved in FA uptake by increasing the rate of intracellular esterification (Xu et al., 2013). Previous reports have demonstrated that in both cattle (Bionaz and Loor, 2008b) and humans (Mohammad and Haymond, 2013), CD36 mRNA abundance is the highest among all FA transporter genes in mammary tissue. Consistent with these reports, we confirmed that CD36 had the highest mRNA abundance among all FA transporter genes and that both the protein and mRNA levels for CD36 were upregulated in porcine mammary tissue at the onset of lactation (Fig. 2A and C).
FA activation and intracellar transport in mammary cells
Once LCFAs enter cells, ACSLs convert them to acyl-CoAs in an ATP-dependent reaction (Mashek et al., 2006). The ACSL isoforms differ in their substrate preferences, enzyme kinetics, and intracellular locations, and then each ACSL isoform channels FA toward separate metabolic fates (Quesada et al., 2011). In the present study, ACSL3 was the most abundant isoform in the porcine mammary gland, accounting for ∼75% of all ACSL mRNA measured (Table 3), although it has been reported that ACSL1 is predominant among ACSL isoforms in lactating bovine (Bionaz and Loor, 2008a) and human (Mohammad and Haymond, 2013) mammary tissue.
This inconsistency in our results with previous reports probably indicates that species differences exist in ACSL mRNA isoform expression in lactating mammary cells. Fujino et al. (1996) reported that rat ACSL3 prefer C16–C20 unsaturated FAs, which are major constituents of FAs in sow milk (∼60%). Considering that ACSL3 mRNA expression was markedly upregulated throughout lactation, we speculate that ACSL3 mainly channeled LCFA towards TAG synthesis during lactation in porcine mammary tissue.
Among the enzymes involved in the activation of short chain FAs (SCFAs), three ACSS isoforms (ACSS1, ACSS2, ACSS3) were detected, and their expressions were increased several-fold in the porcine mammary gland throughout lactation (Table 3; Fig. 3A). ACSS2 had greater mRNA abundance and expression upregulation than did ACSS1 or ACSS3 (Table 3; Fig. 3A). ACSS2 has been reported to channel acetate towards FA synthesis (Luong et al., 2000) and correlate with mammary de novo FA synthesis (Mellenberger et al., 2009; Yadav et al., 2015). Thus, the observation that ACSS isoforms increase at the onset of lactation suggests that the proteins encoded by these genes provide activated acetate (acyl-CoA) for de novo FA synthesis and a carbon source for energy generation in the mammary gland (Bionaz and Loor, 2008b; Mohammad and Haymond, 2013).
FABP and ACBP are the main intracellular FA transporters in the mammary gland in many species (Bionaz and Loor, 2008a; 2008b; Mohammad and Haymond, 2013). FABP has high affinity for LCFA-CoA, but it can also bind acyl-CoA (Frolov et al., 1997). We showed here that FABP3 is the most abundant isoform in this gene family (∼ 96% of all FABP isoforms, Table 3) and that its expression at the mRNA and protein levels is markedly upregulated throughout lactation (Fig. 3C and D), while other isoforms exhibit only small changes, with low mRNA abundance. These results are in accordance with previous reports that showed that FABP3 was the predominant isoform in the mammary glands of other species (Bionaz and Loor, 2008a; Mohammad and Haymond, 2013).
De novo FA synthesis and desaturation in mammary gland cells
FASN and ACACA are considered crucial enzymes to mammary gland cellular de novo FA synthesis, and their mRNA expression is strongly associated with mammary de novo lipogenesis (Bauman et al., 2011). In this study, the marked upregulated mRNA and protein levels of ACACA and FASN throughout the lactation period (p < 0.05, Fig. 4A, C, and D) could explain, at least in part, the increased DNSFAs in milk (Table 1). In agreement with our results, upregulation of mammary ACACA and FASN mRNA during lactation has also been reported in humans (Mohammad and Haymond, 2013) and bovines (Bionaz and Loor, 2008b).
Long-chain polyunsaturated fatty acids (LC-PUFAs) are essential to the normal growth and development of the brain and retina of the newborn. The maternal organism needs to synthesize LC-PUFAs when the required amounts are not met by the dietary supply (Makrides and Gibson, 2000). LC-PUFAs in the rat mammary gland can be derived endogenously from precursor EFAs, such as linoleic acid and α-linolenic acid, by consecutive desaturation (FADS1, FADS2) and chain-elongation (ELOVL5) (Brenna et al., 2010; Lattka et al., 2010; Rodriguez-Cruz et al., 2006; 2011). Upregulation of mammary FADS1 and FADS2 is affected by the stage of lactation in bovine species (Bionaz and Loor, 2008b), the rat (Rodriguez-Cruz et al., 2011), and the mouse (Han et al., 2010), with FADS1 mRNA showing a more pronounced and significant increase compared with the mRNA for FADS2.
But in the present study, we found that mRNA expression for FADS1 was increased significantly only at early lactation, while the mRNA for FADS2 remained unchanged during lactation (Fig. 4A). The expression of mammary FADS3 was upregulated during lactation, although its mRNA expression was the least abundant isoform during lactation among the FADS isoforms present. The results indicate that FADS1 plays a major role in the biosynthesis of LC-PUFAs in the lactating porcine mammary gland and that FADS3 may take part in this process as well.
The mRNA expressions of ELOVL1, ELOVL5, and ELOVL6 were all upregulated (Fig. 4B) during lactation in the porcine mammary gland, similar to what has been observed in humans (Mohammad and Haymond, 2013). ELOVL1 had the highest increase (6-fold) among the ELOVL genes monitored during this study (Fig. 4B). Our results suggest that the mammary gland participates in LC-PUFAs synthesis in sows from late pregnancy and throughout lactation by expressing desaturase (FADS1, FADS3) and elongase enzymes (ELOVL1, ELOVL5, ELOVL6).
SCD is an important enzyme in the mammary gland, as it converts SFA into MUFA by introducing a double bond at the Δ9 location of primarily myristoyl-, palmitoyl-, and stearoyl-CoA (Conte et al., 2010). In this study, the concentrations of MUFA nearly doubled from colostrum to milk (Table 1), which was probably associated with the dramatically increased mRNA level (Fig. 4A) and protein expression (Fig. 4C) of mammary SCD throughout lactation. Our results are consistent with those from a previous report that demonstrated that SCD mRNA plays a crucial role in TAG synthesis in the lactating bovine mammary gland based on its mRNA abundance (23% of total) and upregulation (40-fold) during lactation (Bionaz and Loor, 2008b).
TAG synthesis in the lactating mammary gland
Once synthesized and activated, FAs are esterified to glycerol-3-phosphate to produce triglycerides. The enzymes responsible for triglyceride esterification, including GPAM, AGPAT1, LPIN1, and DGAT1, were the major isoforms within each gene family detected in the lactating porcine mammary tissue (Table 3) in this study. Additionally, the mRNA for GPAM was the most abundant among this group of genes (GPAM > AGPAT1 > LPIN1 > DGAT1) (Table 2), and the expression of GPAM, AGPAT1, LPIN1, and DGAT1 both at the mRNA and at the protein level were markedly up-regulated at peak lactation (Fig. 5A,C, and D).
The results are consistent with those observed in lactating bovine (Bionaz and Loor, 2008b). In contrast, LPIN1 was reported to be the most abundant gene in the human model (LPIN1 > AGPAT1 > DGAT1 > GPAM) with respective mRNA expressions upregulations of 4-, 2.5-, 1.2-, and 9.0-fold during the onset of lactation (Mohammad and Haymond, 2013).
It has been reported that LPIN protein function and glycerolipid synthesis are influenced by intricate functional interactions between LPIN family members because they act in an interdependent manner to optimize lipid homeostasis in various tissues (Csaki et al., 2013; Dwyer et al., 2012). For example, LPIN1 and LPIN2 cooperate to maintain glycerolipid homeostasis in the liver and cerebellum of young mice (Dwyer et al., 2012). Additionally, tissue-specific relationships among LPIN family members have been demonstrated in eight different tissues in mice (Csaki et al., 2013).
The LPIN1 mRNA in mammary tissue was reported to be the most abundant among LPIN isoforms in human (Mohammad and Haymond, 2013), mouse (Han et al., 2010), and bovine tissue (Bionaz and Loor, 2008a), and it showed a significant increase during lactation. In this study, LPIN1 was predominant (55%) at peak lactation, and it increased 6.7-fold (p < 0.05) at d 17 of lactation, while the level of LPIN2 doubled (p < 0.05) (Table 3; Fig. 5C). LPIN3 mRNA expression was significantly downregulated during lactation (p < 0.05; Fig. 5C). We speculate that, a major role of LPIN1 and a minor role of LPIN2 may be played in TAG synthesis in the porcine mammary gland during lactation, while LPIN3 may play a negligible role.
Milk lipid droplet formation in mammary cells
Milk fat is secreted as a globule surrounded by a globular membrane and proteins such as PLIN, BTN, and XDH (McManaman et al., 2007; Reinhardt and Lippolis, 2006; Robenek et al., 2006). In the present study, the mRNA of PLIN2 was the most abundant among this group of genes (PLIN2 > XDH > BTN2A1) (Table 2), and the respective mRNA expressions were upregulated 5.5-, 187-, and 4.6- fold at peak lactation (Fig. 6A). A similar pattern has been seen in the bovine mammary gland, where PLIN2 was also the most abundant gene (PLIN2 > XDH > BTN1A1), with respective increases of 3-, 8-, and 14- fold by day 60 postpartum (Bionaz and Loor, 2008b). XDH was demonstrated to be the most abundant gene (XDH > PLIN2 > BTN1A1) in the human model and their respective mRNA expressions were up-regulated 8-, 1.3-, and 7.5-fold during the onset of lactation (Mohammad and Haymond, 2013).
PLIN2 localizes on the cytoplasmic lipid droplets (CLDs) in mammary epithelial cells and milk fat globules (MFGs) (Russell et al., 2007; 2008). Increased PLIN2 expression was reported to initiate the sequentially ordered process of CLD accumulation in mammary epithelial cells (Russell et al., 2007). Microarray analysis indicated that PLIN2 transcripts were among the most abundant mRNAs in the lactating mouse mammary gland, equivalent in level to that of other secreted milk proteins such as the caseins (Rudolph et al., 2007).The PLIN2 transcript and protein levels in milk-secreting cells dramatically increased as the mammary gland underwent differentiation into a secretory organ over the course of pregnancy (Russell et al., 2007).
In this study, we confirmed the predominance of PLIN2 mRNA in the porcine mammary gland, which is consistent to what was observed in human (Mohammad and Haymond, 2013), bovine (Bionaz and Loor, 2008b), and mouse models (Russell et al., 2008). As well, we showed here that both the protein and the mRNA levels of porcine mammary PLIN2 were increased at peak lactation compared with late pregnancy (p < 0.05, Fig. 6A and B). The results suggest a major role for PLIN2 in porcine mammary lipid droplet formation.
Transcription factors and nuclear receptors for the regulation of TAG synthesis in the mammary gland
Previous studies have shown that milk lipid synthesis is regulated by many transcription factors and nuclear receptors, including SREBPs, SCAP, INSIGs, THRSP, PPARs, and LXR through the activation of lipogenic genes (Bionaz and Loor, 2008b; Kadegowda et al., 2009; Li et al., 2015; Ma and Corl, 2012; Oppi-Williams et al., 2013; Shi et al., 2013; 2014). Many of these genes were expressed at high levels in the lactating mammary gland (Bionaz and Loor, 2008b; Mohammad and Haymond, 2013; Yadav et al., 2015). Among these genes, SREBP1 plays a central role in mammary gland lipid synthesis during lactation (Bionaz and Loor, 2008b; Mohammad and Haymond, 2013).
In this study, SREBP1 mRNA abundance represented ∼ 2.5% of all the genes measured in the lactating porcine mammary gland, which was much more than the other transcription factors measured (Table 2). Both the protein and mRNA levels of porcine mammary SREBP1 were increased at peak lactation compared with late pregnancy (p < 0.05, Fig. 7A and C). It has been reported that SREBP1c at parturition is the most important isoform in secretory activation in the mouse and that it preferentially promotes the expression of FA biosynthesis related genes (Rudolph et al., 2010), but that disruption of this gene results in a 41% decrease in milk fat concentration (Rudolph et al., 2007). Generally, most of the key enzymes involved in lipogenesis are transcriptionally regulated by SREBP1, which is highly expressed in the lactating bovine mammary gland (Bionaz and Loor, 2008b). Therefore, we speculate that SREBP1 is one of the most important transcription factors regulating TAG synthesis in the lactating porcine mammary gland.
In the lactating mammary gland, INSIG1 is another important transcription regulator that activates the genes that drive the lipid synthesizing machinery (Bionaz and Loor, 2008b). It is reported that INSIGs block the processing of SREBPs by binding to SCAP in a sterol-regulated fashion, thus preventing it from escorting SREBPs to the Golgi apparatus and altering the rate of lipogenesis (Engelking et al., 2004; Li et al., 2003).
In a previous study, it was suggested that high levels of INSIG1 create a situation in which low levels of endogenous sterols can trigger SCAP binding to INSIG1 without the necessity for exogenous sterols (Espenshade and Hughes, 2007). In this study, mRNA expression for INSIG1, INSIG2, and SCAP were increased during lactation; ∼73-fold for INSIG1, ∼48-fold for INSIG2, and 3.5-fold for SCAP at peak lactation (Fig. 7A). The results suggest that INSIG1 and INSIG2 play an important role in orchestrating lipid metabolism in the porcine mammary gland during lactation.
PPARγ is reported to play a crucial role in regulating TAG synthesis and secretion by upregulating the PPARγ lipogenic target genes that are related to TAG synthesis and secretion in bovine (Kadegowda et al., 2009) and goat (Shi et al., 2013) mammary cells. Previous bovine data indicated that PPARγ mRNA predominated and was upregulated in mammary gland tissue during lactation (Bionaz and Loor, 2008b). In this study, despite being the main PPAR isoform, PPARγ mRNA expression was not affected by the stage of lactation (Fig. 7B), while its protein expression was increased at early lactation compared with late pregnancy (p < 0.05, Fig. 7C).
Another PPAR isoform, PPARα, was upregulated both at its gene and protein expression levels from late pregnancy to peak lactation (p < 0.05, Fig. 7B and C), while the percent PPARα abundance in the PPAR isoforms was increased from 29% (d −17) to 38% (d 17), and then its abundance reached equivalence with PPARγ) (Table 3). Similarly, PPARα target genes involved in FA transport, such as CD36, ACSL3, and FABP3, were also upregulated significantly in the lactating porcine mammary gland, as had been observed in lactating mouse (Rudolph et al., 2007) and bovine models (Bionaz and Loor, 2008a; 2008b). Therefore, we speculate that activation of PPARα promotes FA transport by upregulating to various extents the lipogenic target genes in the porcine mammary epithelial cells that are involved in FA transport, such as CD36, ACSL3, and FABP3.
Conclusions and Perspective
Taken together, these findings inform future omics strategies to increase milk fat production and milk lipid profiles and further attest to the rise of veterinomics in postgenomics life sciences. Notably, the content of each FA as well as the TAG content of milk increased during porcine lactation and that regulation of lipid synthesis and secretion in the porcine mammary gland occurs at the level of mRNA and protein expression.
Transcripts of SLC27A3, ACSS2, ACSL3, FABP3, FADS1, ELOVL1, DGAT1, AGPAT1, LPIN1, PLIN2, SREBP1, INSIG1, INSIG2, and PPARγ were the most abundant within each specific gene family, and these isoforms, except for FADS1 and PPARγ, were upregulated from late pregnancy to peak lactation.
Compared with late pregnancy, marked upregulation with high relative mRNA abundance during lactation were observed for those genes involved in milk TAG synthesis and secretion including those related to FA uptake (VLDLR, LPL, CD36), FA activation (ACSS2, ACSL3) and intracellar transport (FABP3), de novo FA synthesis (ACACA, FASN), FA elongation (ELOVL1), FA desaturation (SCD, FADS1), TAG synthesis (GPAM, AGPAT1, LPIN1, DGAT1), lipid droplet formation (BTN2A1, XDH, PLIN2), and transcription factors and nuclear receptors (SREBP1, SCAP, INSIG1/2). Western blot analysis of CD36, FABP3, ACACA, SCD, FASN, GPAM, AGPAT1, LPIN1, DGAT1, PLIN2, and SREBP1 proteins confirmed their increase in lactating porcine mammary tissue compared with late pregnancy.
The milk TAG synthesis and secretion in the porcine mammary gland are supported by an increase in a wide variety of lipogenic genes and the expression of their respective proteins during lactation. The lipidomics method used in this study could be applied to develop effective biomarkers for important traits as target for genomic selection or animal management in the livestock industry. For example, the polymorphisms in AGPAT1 can be used to select for animals producing healthier milk with higher concentration of PUFA and linoleic acid as the AGPAT1 haplotypes in Holstein dairy cattle were reported to be significantly associated with higher concentrations of PUFA and linoleic acid in milk (Nafikov et al., 2014).
The dietary composition, and especially the sources of energy, for lactating animals should also be considered as varied nutrients are potentially important in promoting (Jacobs et al., 2015; Liu et al., 2013) or inhibiting (Hussein et al., 2013; Wang et al., 2014) lipogenesis. In practical production, to increase the feed intake of lactating sow or to supplement functional nutrients, such as folic acid or conjugated linoleic acid during late gestation, are potential nutritional strategies to increase sow milk production (Krogh et al., 2012; Wang et al., 2011).
Metabolic diseases, such as laminitis, metritis, fatty liver, and ketosis, are common in lactating sows and dairy cows. The pathogenesis of metabolic diseases is associated with the disturbance of one or more blood metabolites in the sick animals. The complex interaction between metabolic disorders makes a single treatment for metabolic diseases less effective, which suggests that a highly integrated research strategy is required to more fully understand the biological complexities of metabolic diseases.
Recent advances in high throughput ‘omics’ technologies (multi-omics), which involves the combination of genomics, transcriptomics, proteomics, lipidomics, and metabolomics provides a comprehensive understanding of cell metabolism as one ‘integrated system’ (Günther et al., 2014; Kollmus et al., 2014; Longuespée et al., 2014; Montague et al., 2014;). Multi-omics characterizes the molecular and metabolic pathway derangements that might play a role in the pathogenesis and progression of complex diseases (Civelek and Lusis, 2014; Li et al., 2015).
Therefore, muti-omics studies could be applied to understand, predict, diagnose, and monitor diseases (Civelek and Lusis, 2014; Li et al., 2015; Stanberry et al., 2013), and to identify drug targets (Kurland et al., 2013). Accordingly, in the future, this integrated approach to veterinary medicine may provide an innovative way to monitor the efficacy of therapy.
Footnotes
Acknowledgments
This study was supported by the National Research Program of China (No. 2013BAD10B01-4). The authors thank Dr. Y.Z. Chen and his colleagues from the Animal Hospital of South China Agricultural University for technical assistance with the biopsies. The authors also thank Dr. W.W Riley for his help in the presentation of this manuscript.
Author Disclosure Statement
The authors declare that they have no financial conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.