
Abstract
Abstract
In recent years, rapid developments in several omics platforms and next generation sequencing technology have generated a huge amount of biological data about plants. Systems biology aims to develop and use well-organized and efficient algorithms, data structure, visualization, and communication tools for the integration of these biological data with the goal of computational modeling and simulation. It studies crop plant systems by systematically perturbing them, checking the gene, protein, and informational pathway responses; integrating these data; and finally, formulating mathematical models that describe the structure of system and its response to individual perturbations. Consequently, systems biology approaches, such as integrative and predictive ones, hold immense potential in understanding of molecular mechanism of agriculturally important complex traits linked to agricultural productivity. This has led to identification of some key genes and proteins involved in networks of pathways involved in input use efficiency, biotic and abiotic stress resistance, photosynthesis efficiency, root, stem and leaf architecture, and nutrient mobilization. The developments in the above fields have made it possible to design smart crops with superior agronomic traits through genetic manipulation of key candidate genes.
Introduction
A
Development of smart crops that produce more food in a short period, reduce our need on chemicals such as pesticides and fungicides, and add to environmental friendly agriculture has become important to ensure food and nutritional security in many parts of the worlds today. Anticipated changes in climate and its variability, mostly extreme temperatures and alterations in rainfall, are expected to make crop improvement even more crucial for food production (Atkinson and Urwin, 2012; Redden et al., 2013; Varshney et al., 2011).
Since there has been increasing interest in health-promoting qualities of food, the future holds growing opportunities for crop improvement with the objective of development of functional food and neutraceuticals. Advances in our understanding of functional genomics, systems biology, synthetic pathways and their regulation, and new approaches to use this knowledge to improve the level of quality constituents have the potential to add value to crop products and contribute to society's health.
Over the last 30 years, advances in plant molecular biology and biotechnology have led to one of the great exciting and productive times in crop plant research. This has mostly been accomplished via a reductionist approach for biological insight. This approach has been greatly successful in identifying novel molecules that play key roles in complex biological activities; however, it requires a direct relationship between the gene and its function (Benfey et al., 2008; Westerhoff et al., 2004).
Significant development has been made in identification of traits/genes and its networks involved in central biological processes. Yet, how these genes and gene networks are coordinated over raising levels of biological complexity, beginning from cells to tissues and tissues to organs, remains unclear (Jaimie et al., 2009). Systems biology approaches are beneficial to solve complex biological problems that will be useful to develop new biotechnological approaches in crop improvement programs (Fig. 1).
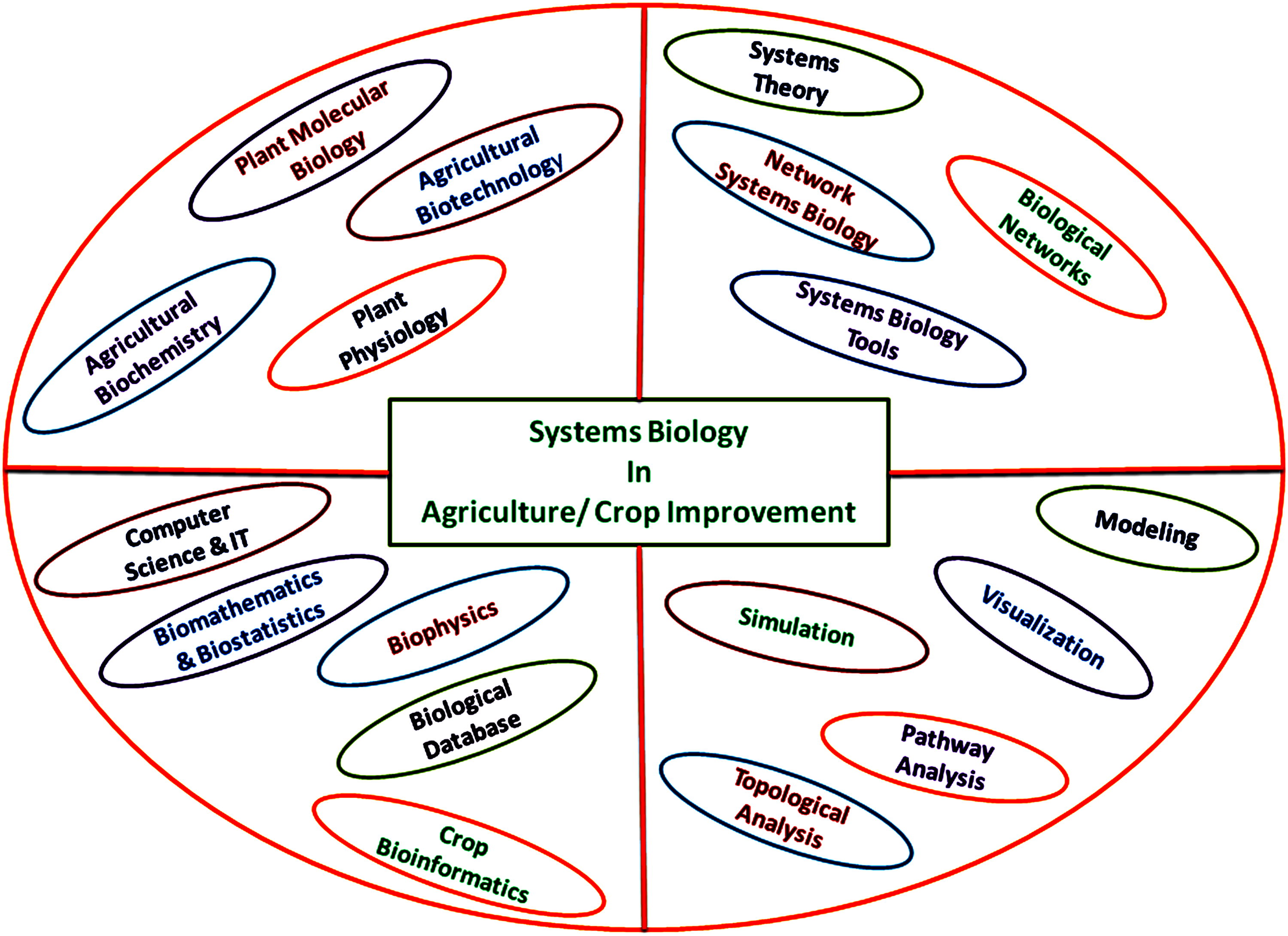
Systems Biology: An integration of interdisciplinary approaches for solving biological problem with respect to agriculture/crop improvements.
The term “systems biology” has come out recently to explain the frontier of cross-disciplinary research in life sciences (Likic et al., 2010; Stelling, 2004) (Fig. 1). Nearly a decade has passed since systems biology was established in the language of modern biology (Ideker et al., 2001, Kitano, 2002). Systems biology is a new field in biology that aims at system-level understanding of biological systems. That means that systems biology recognizes and understands complete biological systems by elucidating, modeling, and predicting the behavior of all components and interactions (gene, proteins, and metabolites) with respect to external stimuli (Junker et al., 2008).
At present, researchers have applied two system-based approaches for decoding the complexity of biological system, one is ‘Top-down’ approach and another is ‘Bottom-up’ approach. In ‘Top-down’ approach, researchers analyze large scale datasets to decode relationships between different levels of transcripts and proteins (Bassel et al., 2012) (Fig. 2). This approach starts with high-throughput experimental ‘omics’ data, followed by data analysis using bioinformatics and systems biology tool for identification of agriculturally important traits.
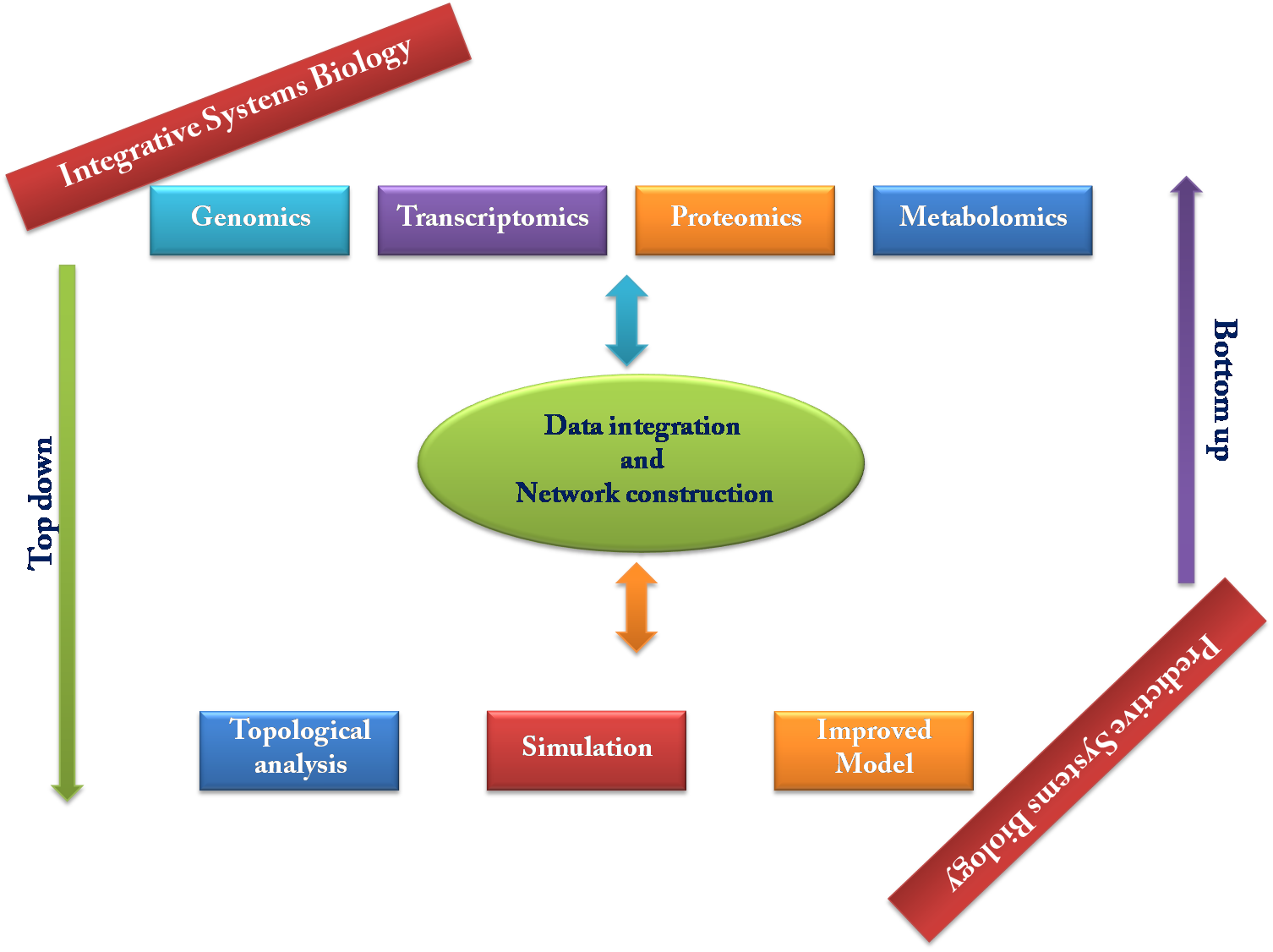
Top-down and bottom-up approaches for defining Systems Biology.
Bottom-up approach is an integrative one in which properties of genes/proteins with available quantitative information such as kinetics data and transcription rates are used to construct a model of well-characterized components (gene/protein, etc) that can be simulated computationally to identify and predict the behavior of system in different conditions. Systems biology attempts to study living systems by determining all of the components and the associations between those components in the perspective of the entire systems.
The term “Network” is a most important keyword for systems biology (Chen et al., 2009; Danning et al., 2012). Networks organize the complexity of biological systems as components (nodes) and interactions (edges) between them. Biological processes are often represented in the form of networks such as protein–protein interaction, signal transduction, gene regulatory, and metabolic networks (Fig. 3). Such research efforts in which novel biological networks are designed from a model will eventually converge and couple with efforts to study existing biological systems. In view of this scenario, one would not only possess predictive models but would also have the power to use these models to re-engineer cells.

Networks in Systems Biology for modeling, analysis, prediction, and identification of complex traits.
A range of potential modifications could be rigorously evaluated through model simulations that later can be verified directly in the biological system. The first stage in a systems-based approach is to identify all of the components involved in systems functionalities (e.g., genomics, transcripts, proteome, and metabolites), followed by perturbation of the systems and monitoring of its response in defined conditions. The investigated information can then be incorporated into models of system function (Gutierrez et al., 2005; Rhee et al., 2006). New hypotheses generated by these models can be tested experimentally, which may lead to revised models and novel testable hypotheses.
Today, there is a growing need of systems biology for successful identification and introduction of the desirable traits such as disease resistance, insect resistance, herbicide tolerance, chilling tolerance, delayed fruit ripening, as well as prolonged shelf-life for the development of smart crops with national and international importance. This review provides recent developments and advances in the practical applications of systems biology in agriculture. The principal approaches are generally as follows.
Integrative systems biology
Integration of ‘omics’ data sets; computer modeling of disease or disease condition level information available in the literature and databases, give novel insight into biological processes or systems at molecular and physiological levels and provide an integrative view of these omics data. This approach, which can decipher existing biological information and large-scale omics data to improve our understanding with respect to unknown traits of a system of interest (Auffray et al., 2010; Greene et al., 2010), is also known as integrative systems biology or Top-down approach.
Predictive systems biology
Predictive systems biology, also known as the Bottom-up approach, provides a good approximation of the quantitative behavior of crop plant systems in various climates, as well as abiotic and biotic conditions, by using integrative model simulation, analysis, and interpretations (Hood et al., 2004; Liu, 2005). Integrative and predictive systems biology approaches explain biological phenomenon not on a gene by gene or trait basis but through the net interactions in cell or whole systems for all cellular and biochemical components (Wynn et al., 2012). These advanced complementary approaches, which must ultimately be integrated to understand the complex plant systems, are already having an impact on the crop improvement programme.
Important Agricultural Traits Linked with Agricultural Productivity
Sustainable agriculture relies on conserved action of agricultural and wild biodiversity management of soil and water with minimization of external inputs usage. Systems biology has a potential for development of sustainable agriculture; it can be used to model and analyze complex traits linked with agricultural productivity such as nitrogen use efficiency, photosynthetic efficiency, water use efficiency, and other essential traits associated with plant architecture and plant physiology for future biotechnological applications in crop plants that provide new dimension with context of improvement, growth, and productivity.
Plant growth and development
An increase in the mass or size during the life of a plant due to cell division and cell expansion is termed growth and development. Cell division in meristems is responsible for increasing cell number; it increases the potential of growth; however, cell expansion accounts for the actual increase in plant mass. Plant hormones are recognized as the regulators that play crucial roles in growth and development.
Band et al. (2012) used a mathematical model associated with measurements of root growth and developmental dynamics to investigate the distribution of gibberellic acid within the root elongation zone through simulation analysis. The integration of the gibberellic acid signaling network facilitated an in silico simulation of how GA would affect the various downstream components. The ultimate challenge of plant biology is characterization of not only but also single molecules and cells (Araújo and Fernie, 2012). This will require a systems biology approach for better understanding of biological component involved in plant growth and development.
Nitrogen use efficiency (NUE)
Nitrogen (N) is the constituent of numerous important compounds, including amino acids, proteins (enzyme), nucleic acid, chlorophyll, and several plant hormones. It comprises 1.5%–2% of plant dry matter and approximately 16% of total plant protein. It is not only the most important mineral nutrient for plants but also the single most important factor in deciding crop yield and grain protein content. To meet ever increasing demands of food due to burgeoning world population, nitrogenous fertilizers have been excessively used to enhance production of cereal crops such as rice, wheat, and maize, which are staple food crops for mankind.
Nitrogenous compounds used are usually available in nitrate and ammonium form and are extremely mobile in the soils. Plants are capable of using only 30%–40% of the applied N, because more than 60% of the soil N is destroyed by a combination of leaching, microbial consumption, surface run-off, de-nitrification, and volatilization. Further, it has now been realized that this un-utilized nitrogen has become a potential environmental threat. Continuous increments of nitrates in the ground water have now been reported to cause cancer, methemoglobinemia, or the blue baby syndrome, which is a disease of bottle-fed infants. This occurs when the nitrate level increases beyond 22.6 mg/I Nitrate-N in drinking water.
The overall NUE of plants comprises both uptake and utilization efficiencies, which can be calculated as UpE × UtE = Nt/Ns × Gw/Nt = Gw/Ns (Gw, total grain (seed) weight; Ns, total N provided to the plant; Nt, total N transported to the seeds; UpE, N uptake efficiency; UtE, N utilization efficiency). It is approximated that 1% enrichment in NUE could save $1.1 billion annually. Thus, it is significant to increase NUE of crops to minimize the loss of N and decrease environmental pollution with reduction input cost. NUE of cereal crops could be enhanced by employing better agricultural practices, as well as through genetic engineering techniques. Modern agricultural practices such as precision farming and nano-delivery methods have been put to practice, but with limited success because newer crop genotypes generated through genetic engineering need to be developed that respond well to these agricultural practices.
However, the molecular information governing genetic variation among crop plant varieties, as well as hybrids for NUE, is poorly known. In order to apply molecular breeding and biotechnology approaches to improve crops for the important and complex trait like NUE, it is crucial to have a broad knowledge of the gene regulatory mechanisms controlling N use, mainly when the amount of nitrogen is limited in the environment.
There are two common stages for N use in the crop plant life cycle. The first stage is the quantity of N uptake and storage, as well as assimilation into amino acid and other important nitrogenous compounds during biomass production. The second stage is the quantity of nitrogen that is partitioned into the seed, ensuing in the final yield. Therefore, understanding the mechanisms for N-update, assimilation, translocation, distribution, and remobilization during the life cycle of crop plant is essential for increasing NUE.
Until now, several strategies such as identification of quantitative trait locus (QTLs) through molecular marker technology, isolation of candidate traits or genes, understanding and identification of regulatory proteins and C/N sensors, and genetic transformation events have been carried with partial success due to the complex nature of traits. In addition to this, nonavailability of a model plant and limited gene pools for this trait in common cereals makes it difficult to achieve the target of achieving high NUE under low N conditions. Therefore, key genes (GS, GOGAT, PEP, AS, GDH, etc.) and model plants (Arabidopsis, finger millets, etc.) need to be identified, which can be utilized for both understanding the complex traits of NUE and to develop crop genotypes that have high NUE.
Systems-based analyses have identified genes and pathways (e.g., nitrate assimilation, pentose phosphate pathway, and glycolysis) that respond to nitrate under a variety of conditions (Gutiérrez et al., 2007). To identify the link between carbon- and/or light-regulated genes in wild-type and the misregulated genes in carbon and light-insensitive mutants (cli186), the microarray data is interpreted in the context of metabolic and regulatory networks. The network constructed from the 966 light/carbon-regulated genes in wild-type, show that cli186 is affected in the light and/or carbon regulation of a network of 60 connected genes, including six transcription factors.
One of the transcription factors, HAT22, is a regulatory “hub” in the cli186 network as it shows regulatory relations connecting to a metabolic network of genes involved in amino acid metabolism, C-compound/carbohydrate metabolism, and glycolysis/gluconeogenesis (Thum et al., 2008). Literature studies show that genetically distinct systemic signaling underlying plant N economy: (i) N supply, corresponding to a long-distance systemic signaling triggered by nitrate sensing; and (ii) N demand, experimental support for the transitive closure of a previously inferred nitrate–cytokinin shoot–root relay system that reports the nitrate demand of the complete plant systems, promoting a compensatory root growth in nitrate-rich patches of heterogeneous soil (Ruffel et al., 2011).
The cell-specific regulation of a transcriptional circuit mediates lateral root out growth in response to nitrogen via microRNA167, linking small RNAs to nitrogen responses (Gifford et al., 2008). The integrated systems biology approach will certainly play a pivotal role in understanding the complex traits of NUE and related issues. It will address such complex issues and provide modalities in the form of products, process, practices, and new knowledge for enhancing agricultural productivity (Bi et al., 2014; Kant et al., 2011, 2012).
Photosynthetic efficiency
Photosynthesis is a biological process by which plants, algae, and photosynthetic microbes use light energy to derive the synthesis of organic compounds as a food, governed by complex photosynthetic traits for growth and development. In the last decades, increasing crop productivity of some major crops such as wheat, rice, and Brassica have shown slight improvement. In this regard, improvement in photosynthetic efficiency of crop plant will prove instrumental in increasing yield of these crops.
The major challenges in improving photosynthetic efficiency of crop plants is to know how alternations should be made to photosynthetic process. In addition, the process of photosynthesis is strongly influenced by various abiotic and biotic stresses, as well as water transport (Zhu et al., 2010). The C4 photosynthetic cycle supercharges photosynthesis by concentrating CO2 around ribulose-1,5-bisphosphate carboxylase and significantly reduces the oxygenation reaction. Therefore, engineering C4 features into C3 plants has been suggested as a feasible way to increase photosynthesis and yield of C3 plants such as rice, wheat, and potato.
Systematic comparison of C3 and C4 metabolic networks using the improved constraint-based models for Arabidopsis and maize show that C3 network exhibits more dense topology structure than C4. The simulation of enzyme knockouts demonstrated that both C3 and C4 networks are very robust, especially when optimizing CO2 fixation. Moreover, a C4 plant has better robustness no matter if the objective function is biomass synthesis or CO2 fixation. In addition, all the essential reactions in C3 networks are also essential for C4, while there are some other reactions specifically essential for C4, which validate that the basic metabolism of C4 plant is similar to C3, but C4 is more complex.
Research studies identified more correlated reaction sets in C4, and demonstrated that C4 plants have better modularity with complex mechanism coordinates the reactions and pathways than that of C3 plants. The increase of both biomass production and CO2 fixation with light intensity and CO2 concentration in C4 is faster than that in C3, which reflects more efficient use of light and CO2 in C4 plant (Wang et al., 2012).
Targeted omics-based research can generate huge amounts of proteomics and genomics data with respect to photosynthesis, integrated approach using systems biology will model these huge data to identify agriculturally important traits for improvement of photosynthetic efficiency in crop plants (Bonardi et al., 2005; Bräutigam et al., 2014; Kanwal et al., 2014).
Water use efficiency
Water use efficiency at the physiological level is defined as the measure of a cropping system's capacity to convert water into plant biomass. It comprises the use of water stored in the soil as well as rainfall during the growing period (de Fraiture and Wichelns, 2010). The basic physiological definition of water use efficiency (WUE) equates to the ratio of photosynthesis (A) to transpiration (T), also known as transpiration efficiency.
High water use efficiency plays a vital role in growing crops in drought-prone areas. WUE can be improved by increasing carbon assimilation while keeping the transpiration rate constant, or by reducing the transpiration rate while the carbon assimilation is kept (Yoo et al., 2009). Improving water use efficiency in crop plants is a difficult task; previous research demonstrated that the photosynthetic and chloroplast genes like ERECTA and HRD played essential role in water use efficiency (Nieto et al., 2014).
OnGuard software and models of the stomatal guard cell have recently been developed for exploring stomatal physiology. The analysis suggests that manipulations of single transporter populations are likely to have unforeseen consequences. Channel gates, especially the dominant K+ channels, appear to be the most favorable target for experimental manipulation (Wang et al., 2014). Systems biology approaches can be applied on photosynthetic and chloroplast systems for identification of novel traits that will help in future development of drought-resistant crop plants that could utilize water efficiently for growth and development in various environmental conditions.
Plant architecture
The architecture of plants is commonly referred to as three-dimensional organization systems, which include branching pattern, size, shape, as well as position of leaves and flower organs. It is species specific, indicating that it is under strict genetic control, though it is also influenced by climate conditions such as temperature, light, humidity, and nutrient status, as well as pathogen attack (Reinhardt and Kuhlemeier, 2002).
The molecular and genetic programming that regulates plant architecture is very complex. There are numerous traits involved in plant growth and development, including height, branching degree, branching angles, pattern of flowering, root and shoot growth orientation. Although a majority of the developmental processes involved in plant architecture are known in a model plant, various important traits are still unknown in economically and agriculturally important crop plants.
Research studies indicate that some plant hormones and other signals, such as sugars, are involved in these processes. Therefore genetic and genomic changes that alter hormone synthesis, regulation, and signaling have the potential to induce changes in plant architecture. The identification of specific genes such as MADS-box that are involved in branch orientation, degree of branching, plant size, plant height, and flowering should aid the development of crop plants with improved productivity, Systems biology has become very crucial in understanding plant architecture and essential traits with agricultural importance for benefit of human society (Hollender and Dardick, 2015).
Stress tolerance
Characterization of stress tolerances to various biotic and abiotic stimuli in the plant systems is challenging due to the inherent complexity in molecular mechanisms. Molecular studies help us in deciphering the information at the bio-molecular interaction level, which is not enough to correlate the physiological response (Rouxel et al., 2013; Somvanshi et al., 2014; Yang et al., 2010). Although the conventional techniques are being used in systems biology, their uses to biological complexity draws up new challenges and provides new openings (Pathak et al., 2013).
Abiotic and biotic stress conditions such as drought, heat, salinity, cold, pest, insect, fungus, and bacteria can have a harmful impact on crop plant growth, development, and yield under field conditions (Suzuki et al., 2014). Plant responses to various abiotic and biotic stresses are very complex and involve variations or changes at the transcriptome, proteome, cellular, and physiological levels (Atkinson and Urwin, 2012). The main objective of plant stress biology research is to identify important agricultural traits or genes linked with stress responses that can be useful in the development of stress tolerance crop plants. Identification and functional validation of these traits that act in both abiotic and biotic stress response are excellent candidates and play vital roles in altering stress tolerance (Baena Gonzalez and Sheen, 2008; Miller et al., 2010).
Biotic stress tolerance
It is important to understand plant–pathogen interactions so as to prevent the attack of pest and pathogen to develop disease resistance in plant systems. Systems-based studies on model plants Arabidopsis identified genes such as CIR1, RPS2, RPM1, WRKY31, MPK9, WRKY33, FLS2, RPS4, and RPP13 that play important roles in biotic stress tolerance (Carstens et al., 2014; Chinchilla et al., 2007; Leal et al., 2013). Generally plant physiology gives us a way to understand biotic stress and plant response. It has been pointed out that mere use of biochemistry and molecular biology is no longer useful for some diseases like Alternaria blight, which is the complex disease found in some crop plants such as Brassica.
In our laboratory, we have identified various components involved in pathogenesis of Alternaria blight of Brassica. Besides chlorotic and necrotic toxins, phyto hormones have been suggested to play important roles in the pathogenesis process (Pandey et al., 2001). The chlorotic toxin that has been identified as a cyclo depsipeptide affects many components of MAP kinase machinery, hypersensetive response, and systemic acquired response (Mishra et al. 2011; 2015). Despite the above studies, the potential target gene/protein that could act as key regulatory switch has not been identified. A systems biology approach could help in understanding the molecular mechanism of pathogenesis and solving such recalcitrant problems.
To understand the physiological and molecular mechanisms triggered in plants in response to various biotic stresses, the measurements from all levels of plant organization such as molecules, organelles, cells, tissues, as well as organs and whole plants, and population are necessary (Peterson and Higley, 2000). Advances in systems biology offer a prospect for greatly improved understanding with respect to biotic stresses that will help in providing disease resistance in crop plants.
Abiotic stress tolerance
Plants are exposed to a complex set of abiotic (salt, drought, heat, cold) stresses, and responses to these stresses are equally complex. Studies have revealed the plant genes that undergo altered expression on exposure to abiotic stress, and many of these such as AP2/ERF, MYC, DREB/CBF, MYB, HD-ZIP AREB/ABF, CBL-CIPK and NAC regulons have been proposed to show their role in abiotic stress tolerance (Gupta et al., 2012; 2013a; 2013b). The plant molecular responses to various abiotic stresses involve interactions and crosstalk with different regulatory pathway (Cramer et al., 2011).
A better understanding of regulatory mechanisms involved in crop plant responses during abiotic stresses will enable researchers to investigate and manipulate key regulatory components in order to enhance stress tolerance. Transcription factors (TFs) have played a vital role in crop improvement. Therefore, TFs are good candidates for systems-based genetic engineering to improve crop tolerance to various abiotic stresses because of their function as master regulators of clusters of genes. Various families of TFs were reported, such as CCAAT, homeodomain, NAC, bHLH, bZIP, AP2/ERF, and WRKY that have the potential to be tools for improving abiotic stresses tolerance (Gao et al., 2010; Pathak et al., 2013; Rabara et al., 2014; Shen et al., 2012; Ying et al., 2012; Zhang et al., 2009).
New advances in high-throughput phenotyping, both field based and greenhouse, have to facilitate improved phenomics of transgenics. Systems biology approaches define the underlying changes that result in better yields under abiotic stress conditions. These novel technologies help to show whether manipulating TFs can have effects on crop productivity under field conditions (Rabara et al., 2014).
Nutritional quality
Plants are the primary source of nutritional elements such as macro and micronutrients consumed by humans and animals. In comparison with meat, plants and their products are much less expensive to produce and it is good for better health. However, most plant products are nutritionally imbalanced because of their deficiency in some micro- and macronutrients, as well as some essential amino acids (Sun and Liu, 2004). Crop plants need at least 14 essential mineral nutrients to complete their life cycle. In natural soils, the availability of most essential mineral nutrients is extremely low and does not meet the demands of plants (Gong et al., 2014). Nitrogen (N) is a macronutrient for plant growth and is the essential constituent of many important primary and secondary organic compounds in plants, such as proteins, nucleic acids, and chlorophyll (Xu et al., 2012).
N deficiency can severely inhibit growth, subsequently reduce the yield (Liang et al., 2013). A proteomic analysis of maize roots and leaves in response to N deficiency has been performed using 2-DE and LC-ESIMS/MS (Prinsi et al., 2009). In leaves, TaWIN2, methionine synthase protein, oxygen-evolving enhancer protein 2, ATP synthase subunit alpha, and 23 kDa polypeptide of photosystem II were found to contribute to maize acclimation to N deficiency. The levels of many proteins altered in maize roots are enzymes involved in nitrate assimilation and are components of metabolic pathways involved in the balance of the energy and redox status of the cell.
By contrast, most altered proteins in abundance in maize leaves were involved in photosynthesis regulation. The nutritional level of the plant may affect two post-translational modifications of phosphoenolpyruvate carboxylase, phosphorylation and monoubiquitination in roots and leaves, respectively.
A genome-wide analysis of miRNAs that respond to chronic, transient low N availability in maize has been performed (Xu et al., 2011). miR164, miR169, miR172, miR397–399, miR408, miR528, and miR827 in leaves, and miR160, miR167–169, miR319, miR395, miR399, miR408, and miR528 were detected in roots in response to N-limiting conditions. These miRNAs are mainly involved in the gene expression regulation, signal transduction, energy metabolism, oxidative species scavenging, and encoding the RNA slicer enzyme. Some of the miRNAs, such as miR169, miR398, miR408, miR528, miR827, and miR395, detected above, such as miR169 and miR395, were subsequently demonstrated to respond to N-deficient conditions in maize roots (Zhao et al., 2012).
Iron (Fe) is a micronutrient required for maize growth. However, owing to its low solubility in aerobic and neutral pH environments, Fe availability in soils is extremely low. Proteomic analysis revealed that many proteins change in abundance under low and high Fe conditions, in the plasma membranes of maize roots. These proteins mainly transport proteins, signaling proteins, membrane trafficking proteins, stress-related proteins, redox proteins, metabolism proteins, cell wall-related proteins, cytoskeleton, and protein folding proteins (Hopff et al., 2013). Changes in the transcriptome profiles of roots in response to Fe deficiency were also monitored (Li et al., 2014).
Genes involved in 2-deoxy-mugineic acid (DMA) synthesis, secretion, and uptake of Fe (III)-DMA were significantly induced. Many genes genetically related to protein kinases, plant hormones, and protein phosphatases responded to Fe deficiency stress. As a result, maize responded to Fe deficiency stress in different ways at the proteomic and transcriptomic levels. Systems biology and omics-based approaches can be useful in improvement of the nutritional quality of the crop plants.
Macronutrient improvement
Plants are a major source of food, but have poor content of some important macronutrients such as carbohydrates, lipids, and proteins. The main purpose of agriculture is to provide sufficient food for the people. Researchers have made much effort to improve the quality of protein content in crop plants (Sun et al., 2004). The de novo designed protein, CP3–5, containing 31% lysine and 20% methionine could be expressed and accumulated up to 2% of the total seed protein in the transgenic tobacco seeds lines, resulting in a significant addition in the total lysine content of the mature seeds (Keeler et al., 1997). Systems biology has the ability to identify the complex molecular basis of macronutrient accumulation in crop plant and improving its nutritional quality by balancing the essential amino acid, several minerals and carbohydrates content in plant seeds or edible portions of plants.
Micronutrient improvement
Some major crop plants are a poor source of many essential micronutrients such as iron, zinc, vitamins, and calcium; thus, it is a primary cause of micronutrient malnutrition. Iron, zinc, vitamins, and calcium deficiencies, which are common in developing countries, are responsible for reduced mental capacity, blindness, and decreased work productivity. There is a growing need of improving micronutrient quality in crop plants (Nirgude et al., 2014; Singh et al., 2013). Being complex QTLs involved in such nutritional traits, multi-pronged molecular-physio-genetics and systems biology approaches are being followed for study of mineral nutrition.
Systems Biology of Plant Architecture
Plant systems are complex, regulated by numerous levels of components. The root, stem, leaf, and fruit, as well as seed systems, are fundamental for plant growth, development, and productivity. For modeling of plant systems, two approaches have been used at the whole organism level. The first approach regards the growing plant from the outside, as an organism showing a phenotype interacting with and influenced by its surroundings; the second approach represents it from the inside, as a collection of organ construction by a plant according to internal rules (Lucas et al., 2011).
The first approach would be the same as pure ecophysiological models of plants (Tardieu, 2010). It focuses on reducing the plant to quantifiable or measurable environmentally-sensitive variables whose behavior can be modeled by sets of equations integrating environmental parameters. Ecophysiological models that are directly linked with advances in phenotyping techniques are able to measure phenotypic traits efficiently. Ecophysiological models have been used for a broad range of physiological processes, for example, from Arabidopsis leaf development (Granier et al., 2002) to the way Arabidopsis physiological rates varies with temperature (Parent et al., 2010).
The second approach would be the same as architectural models of plants, In which two main members are Lindenmayer systems (L-systems) and multiple tree graphs (MTG). L-systems are a mathematical formalism to model multicellular organisms (Lindenmayer, 1968). First, the L-systems were devised to give a formal explanation for the development of simple multicellular organisms and to demonstrate the neighborhood relationships among plant cells (Lindenmayer, 1975). They were then extended to explain higher plants and complex branching structures. L-systems support the central concept of rewriting. The concept can be perceived as analogous to the phenomenon of cell division where two daughter cells ‘replace’ a single mother cell or to the developmental process at a higher level where a bud is ‘replaced’ by a branch bearing its own leaves and buds (Lucas et al., 2011; Prusinkiewicz, 2004).
MTGs explain the architecture of trees, which include the topology and tri-dimensional geometry of the plant systems (Godin and Caraglio, 1998; Godin, 2000). They can be applied to model structural plant development through stochastic modeling (Guédon et al., 2001). For example, a stochastic model of Arabidopsis thaliana root development based on MTG was retrieved from a root architectural database (Lucas et al., 2008). Systems biology is used for modeling complex traits that help to predict multigenic traits involved in various biological and physiological processes such as root growth, stem growth, and other information with respect to agricultural productivity. The architecture of plant systems biology is described below.
Root biology
Genomics-based approaches in model plants have enhanced our understanding of root systems biology over the last decade. Currently, systems biology approaches have emerged as important in our understanding of root systems with respect to candidate genes/protein in regulatory pathways. The APSR1 gene, which controls the regulation of root hair elongation and modulates auxin distribution in the root tip, is also involved in regulating root response to Pi availability through PIN7 (González-Mendoza et al., 2013).
ANR1 was reported to play a central role in controlling lateral root development in response to nitrate signal in Arabidopsis (Zhang and Forde, 1998). MADS25 is one of five ANR1-like genes in Oryza sativa that belongs to the ANR1 clade. Recently the role of OsMADS25 in the plant's responses to external nitrate was investigated. It was found that the OsMADS25 protein is found in the nucleus and cytoplasm, and works as a positive regulator to control lateral and primary root development in rice (Yu et al., 2015).
Major advances have been made in employing ‘top down’ systems approaches to identify key molecular players (genes, RNA, protein, etc.), and have identified several gene regulatory pathways that control root growth and development (Bassel et al., 2012; Benfey et al., 2010; Ruffel et al., 2010). We are at the beginning of systems biology, and several challenging issues remain to be addressed in this emerging field (Hill et al., 2014).
Stem biology
The stem cell in plants has an ability to differentiate into different form of specified cell types. A stem cell represents asymmetric cell division by which one daughter cell keeps the characteristics of an undifferentiated mother cell, while the other acquires a precise cell fate, which is activated or triggered by intrinsic and extrinsic signals (Miyashima et al., 2013). Plants grow continuously in apical and lateral directions, making new cells and organs during their life cycle. Cell division, a fundamental process that encourages these growth and development activities, is concentrated in meristems, a specific domain actively dividing undifferentiated cells (Scheres, 2007). The plant vascular systems, made up of xylem and phloem tissues, have evolved to attach plant organs and transport a variety of molecules between them.
In post embryonic development, these conductive tissues are constitutively built from cells, which are derived from a lateral meristemetic cells, known as procambium and cambium. Progress has been made in identifying the molecular mechanisms leading to formation of vascular cambium, but presently regulation of vascular cambium is still poor in comparison with apical meristems. In Arabidopsis thaliana, apical meristems, the positions of stem cells and its niche, have been identified with molecular markers expressed in stem cells or niche cells. The position of stem cells and niche cells in cambium is currently unclear (Miyashima et al., 2013).
Literature studies show that various transcription factors regulate numerous aspects of vascular growth and development including cell differentiation and proliferation. However, very little is known about how these transcription factors construct the regulatory networks during vascular development. Recently, in an attempt to define gene regulatory network, comprehensive yeast one-hybrid as well as two-hybrid assays have been carried out against a numbers of transcription factors expressed in Arabidopsis stele in tissue-specific manners (Brady et al., 2011). Systems biology-based approaches will be very useful in the present as well as in the future for understanding the mechanism of stem cells and its key genes/proteins that play essential roles in regulating plant growth and development in different stages of plant life cycle.
Leaf biology
The leaf is the organ that characterizes plants as autotrophs. During their growth and development, leaves become photosynthetically competent and accumulate nutrients. Leaves then enter their senescence stage, followed by their death. Plant leaves are a unique organ that controls diverse physiological processes in crop plant systems, to understand processes that are associated with aging, senescence, and death (Woo et al., 2013).
In recent years, significant progress has been made in understanding the regulatory mechanisms of leaf development via molecular and genetic based approaches. In addition, increasing use of high throughput technologies is providing new biotechnological information at systems levels. In addition, systems biology provides a way to integrate the accumulating information into holistic computational models for getting a complete understanding of biological processes (Kalve et al., 2014).
Since leaf development is a complex and dynamic process, we must be able to model a single plant cell as it is progressing from the stem cell niche into different places in mature leaf to build a holistic model for leaf growth and developmental phenomenon. The results of integrated behavior and dynamics of all cells ultimately make the leaf an organ of reasonable size and morphology. This is presently a very desirous goal, but as a initial step researchers revealed the current state of knowledge with respect to regulatory networks that work in cells as they progress through their individual developmental pathway (Kalve et al., 2014). Since plant cell are exposed to various biotic and abiotic conditions, during their developmental stages, future development of computational model of different tissues interacting through an interface might provide useful information with respect to candidate genes/proteins.
Fruit biology
Fruits that are characteristic features of Angiospermic plants are found in a variety of forms and types and act an important source of supplementary diet, providing vitamins, minerals, fiber, and antioxidants for animals (Osorio et al., 2013). Fruit maturation and development is an extremely complex process to understand. Systems biology-based approaches allow comprehensive phenotyping at the systems level, which facilitate an overview of metabolite and gene regulatory network (Lee et al., 2012; Osorio et al., 2012; Pan et al., 2013) and a better analysis of the systems regulation underlining fruit development (Osorio et al., 2013).
One of the major objectives of systems biology is to use these networks to predict its dynamics when perturbed by several environmental conditions. Nowadays, much of this research has been completed in tomato due to availability of genetic and genomic resources for this fruit. Targeted metabolic assay, coupled with microarray profiling in the Nr tomato ethylene receptor mutant has led to identification of multiple points of ethylene control throughout fruit development (Alba et al., 2005).
It was observed that 869 genes (mainly ACS, ACO, ETR, CTR, EBF, and EIN) were differentially expressed during normal fruit maturity, and that the mutation of Nr (i.e., ethylene) affected 37% of these genes. In addition, Nr also controlled fruit morphology, seed number, ascorbate accumulation, ethylene evolution, and carotenoid biosynthesis, consistent with the well-known role of ethylene in ripening (Gupta et al., 2006).
Systems biology-based approaches in correlation or co-expression-based studies have been successfully utilized in the metabolic engineering to fill gaps in biosynthetic pathways (Gapper et al., 2014). One such case was identified in the tomato plant, where the genetic and genomic diversity of Solanum pennellii introgression lines was utilized to identify regulators of carotenoid biosynthesis (Chung et al., 2010).
Systems biology has become staggering in the last few years. We now have the blueprints of how plant germinate and undergo complex developmental events through the fast expanding collection of plant genomic sequences. However, understanding these blueprints remains a challenge. Systems biology-based tools have already provided novel insights into underlying biology of fruit development, and a greater prospect in future for understanding the fruit development (Gapper et al., 2014).
Seed biology
Seeds are the main source of nutrients for humans and other animals. Information on seed biology offers tools for better agricultural practices and for managing genetic and genomic resources (Mendoza et al., 2008). The development of genomics has progressed in a number of plant species, resulting in gold-standard genome for some important dicot species such as Arabidopsis, Brassica, and Soybean, as well as monocot species such as rice, maize, and shorghum (Ikeda et al., 2012; Paterson et al., 2010).
Substantial progress has been made in identifying more than 80% of the genes in the large genomes of Triticeae crops such as barley (Mayer et al., 2011) and wheat (Philippe et al., 2012). These resources allow us to investigate genome dynamics by using phylogenomics and comparative genomics analysis tools to facilitate the identification of key genes such as MONOPTEROS, ABI3, FUS3, LEC2, AFL B3 etc. for seed development across a wide range of species that differ substantially in seed size and storage product composition (Goel et al., 2012; Sreenivasulu et al., 2013).
Integration of the different omics data generated through high-throughput technology allows more robust identification of molecular targets for future biotechnological applications in crop plants to manipulate seed size and improve grain quality. Different systems biology approaches have been implemented for identification of regulatory genes and address the missing links between molecules like genes, metabolites, hormone, and understanding physiology of crop plant through the use of bottom-up such as kinetic models, metabolic mass flow, and reaction fluxes, and top-down as gene regulatory and protein–protein interaction networks, metabolic networks and flux analysis strategies (Fukushima et al., 2009; Saito et al., 2010; Yoshida et al., 2010).
Systems biology approaches are just beginning to be broadly used in seed biology studies. They must be applied to more species to broaden the data set for comparative analysis and must be refined and partly adapted to the cellular and subcellular levels. In addition, integration of various high throughput omics data is needed to reconstruct complex networks that determine the phenotypic characteristics in individual seed compartments and identify key regulatory genes (Sreenivasulu et al., 2013).
Systems Biology of Plant Physiological Processes
The physiological mechanism in crop plants such as photosynthesis, respiration, transpiration, nutrition, photoperiodisms, seed germination, and response to various stresses are governed by complex traits involved in various processes. Plants are significantly suitable for systems biology experimentations, because many have relatively short generation period, are easy to handle, and produce sufficient offspring for genetic and genomic analysis, Thus plant systems modeling and experimental validation can be executed very quickly with the excellent systems biology tools available today.
In the model plant Arabidopsis thaliana, much information is available with respect to physiological mechanisms. Knowledge based on Arabidopsis will be very useful for identification of complex physiological traits linked with agricultural productivity in closely related and, ultimately, other plant species (Keurentjes et al., 2011). The challenge for systems biology is to integrate genetic signaling as well as metabolic networks to construct a complete in silico cell for physiological studies in crop plants. This approach has great potential in agricultural research to study the functioning of plant systems at the molecular level to phenotypic level (Pascual et al., 2013; Raikhel et al., 2003). Therefore, in the case of disease, it is interpreted that it results due to systems fault rather than fault in a single component.
Adaptation of external stimuli
Adaptation is a specialized feature of any organism, including plants. Adaptation allows survival and growth in a particular habitat. Plants are challenged at every stage in their life cycle by external stimuli (abiotic and biotic). Plants have the ability to receive specific external and internal signals in response to biotic and abiotic stresses by a cellular receptor, and to initiate specific signals through the signal transduction pathway that increase the level of growth hormones and activate the defense systems with respect to phytoalexin production for crop plant protection, which finally enhances the adaptation.
Nutritional alteration
Plant products play an essential role in our diet, because they contain macro and micro nutrients, which are necessary for growth and development of people and animals. Several factors may have an effect on the nutritional quality of crop plant fruits and products in planting location, climate factor, and harvest season, all of which are responsible for the nutritional alternation of crop products (Livinali et al., 2014). Systems biology will be helpful for the development of environmental friendly crop plants with less nutritional alternation.
Integrated In silico and Experimentation-Based Analysis in Systems Biology
Systems biology is able to provide a clear picture for complex agricultural traits through interconnected networks of biochemical reaction derived from both wet lab experimentation and computational techniques. Systems biology is a science of the twenty-first century that renovates the reductionist approach to a global view and adds a new dimension to agriculture, as well as biological research. In the context of crop protection and agricultural productivity, systems biology involves integrating networks of biochemical reactions through experimental and computational approaches to provide a comprehensive understanding of complex agricultural traits linked with agricultural productivity. It provides better understanding of the systems dynamics in different physiological and environmental conditions, therefore computational approaches are required for curation, pathway modeling, analysis, and visualization of agricultural traits.
In silico systems biology approaches are best used in modeling and analysis of high throughput data generated from next generation sequencing technology for identification of novel traits in crop plants, which can be used through genetic engineering for development of transgenic crop plants with diseases resistance, better productivity, and efficient environmental adaptability.
Design of software
Designing and development of software for analysis of biological systems is a challenging task because teams of researchers from interdisciplinary fields such as biological sciences, physics, chemistry, mathematics, statistics, and computer science with strong backgrounds are required to develop better tool with efficient algorithms. Advances in the field of bioinformatics and increasing data sets generated through high-throughput sequencing technology will provide better improved tools for future understanding of plant responses to various environmental studies
Here we describe the some important systems biology tools with their application (Table 1), which will help to address current and future research problems in agriculture (Ghosh et al., 2011; Jen et al., 2006).
Development of databases
Documentation of available information from literature, genomics, and proteomics data obtained from high-throughput technology with respect to gene, protein, and metabolites with their interacting partner, as well as signal transduction networks created through computational tool is an interesting task for a bioinformatician. The main objective is to collect a huge amount of biological data and information to develop a database in organized form for the benefits of the scientific community (Chelliah et al., 2013; Franceschini et al., 2013; Le et al., 2006; Mi et al., 2013; Ogata et al., 1999; Snel et al., 2000; Thomas et al., 2003). These database resources are very useful for visualization and analysis of complex traits as they can be used for further remodeling of networks and pathway engineering to obtain the dynamics of whole systems (Table 2).
Application of Systems Biology in Agriculture
Plant physiology is an assembly of different biological phenomenon spanning intracellular molecular communications to the whole systems phenotypic response (Kanani et al., 2010). Systems biology attempts to break these multi-scale networks and bridge the link between genotype and phenotype. The organization and dynamics of these pathways/networks are responsible for managing the phenotypic state of a cell (Baghalian et al., 2014; Fraser et al., 2013; Shao et al., 2007). Diverse cells and various tissues coordinate together to produce an organ level response, which further controls the physiological states of plant systems. Such an approach would prove very useful in understanding and managing the problems of agriculture.
The pathway modeling and analysis for targeting complex diseases are expected to find remedies for these diseases. This approach has some advantages in the form of identification of rational agrochemical (Fungicide, Herbicide, and Insecticide) targets, effective agrochemical design with least side effects in humans and environments, effective management strategies, diagnosis of actual source of disease, treatment of disease sources rather than symptoms, early and reliable diagnosis of disease using predictive models. Such an approach has tremendous potential in agricultural research for improving agricultural productivity (Yin et al., 2008; 2010). Some salient applications of systems biology in agriculture is given below.
Breaking yield barriers
Maximizing yield is the aim of scientists working in field of crop improvement and agricultural productivity. The population of world is continuously growing. Huge amounts of agriculture products are required to save the lives of people. Exploiting the potential of systems biology for disease management and crop productivity point of view, has currently received much attention. Recent advances in the field of systems biology can decode the complexity of multiple traits concerned with genotypic to phenotypic responses such as photosynthesis, carbon and nitrogen metabolism, water use efficiency, plant architecture and other physiological mechanisms. Researchers working in areas of agriculture biotechnology with respect to crop improvement and agriculture productivity can understand the power of computational tools and utilize systems biology resources in their research that can be very useful for development of high yielding crop plants for society.
Adaptation to climate change
Agriculture is one of the most important vulnerable sectors to changes in climates, due to its reliance on adequate environmental conditions for achieving high agricultural productivity (Huntingford et al., 2005). Crops plants are affected by shortages or excesses of water or excessively high or low temperatures during growing periods (Porter and Semenov, 2005).
Recently published studies have represented the potential of adaptation strategies design, the two major goals must to be pursued in future studies: (i) a better knowledge of driving processes under future changes in climate; and (ii) a coupling between genetic and crop growth models—perhaps at the expenditure of the number of genes/traits analyzed. Significantly, the latter may imply additional complexity in crop systems modeling studies. Therefore, systems biology approaches will be useful for modularity in crop models as well as individual component testing against observational data, which would be one of the critical components in any attempts to simulate crop-breeding strategies under future climate scenarios (Ramirez-Villegas et al., 2015).
Induction of defense and disease management
Plant–pathogen interaction is a well-known mechanism that involves the activation of various signaling pathways during manifestation of defense response against pathogen. This type of response facilitates the host plant to avoid further infections. Disease resistance and management have always been the main objective of any crop improvement program (Gururani et al., 2012). Efficient use of systems biology tools for disease resistance and management could not only assist us for better understanding of the plant defense signaling, but could also disclose new insight on the molecular interactions networks linked with plant protection and agricultural productivity. The different strategies of systems biology that can be applied for identification of defense-related traits are highlighted in Figure 4.
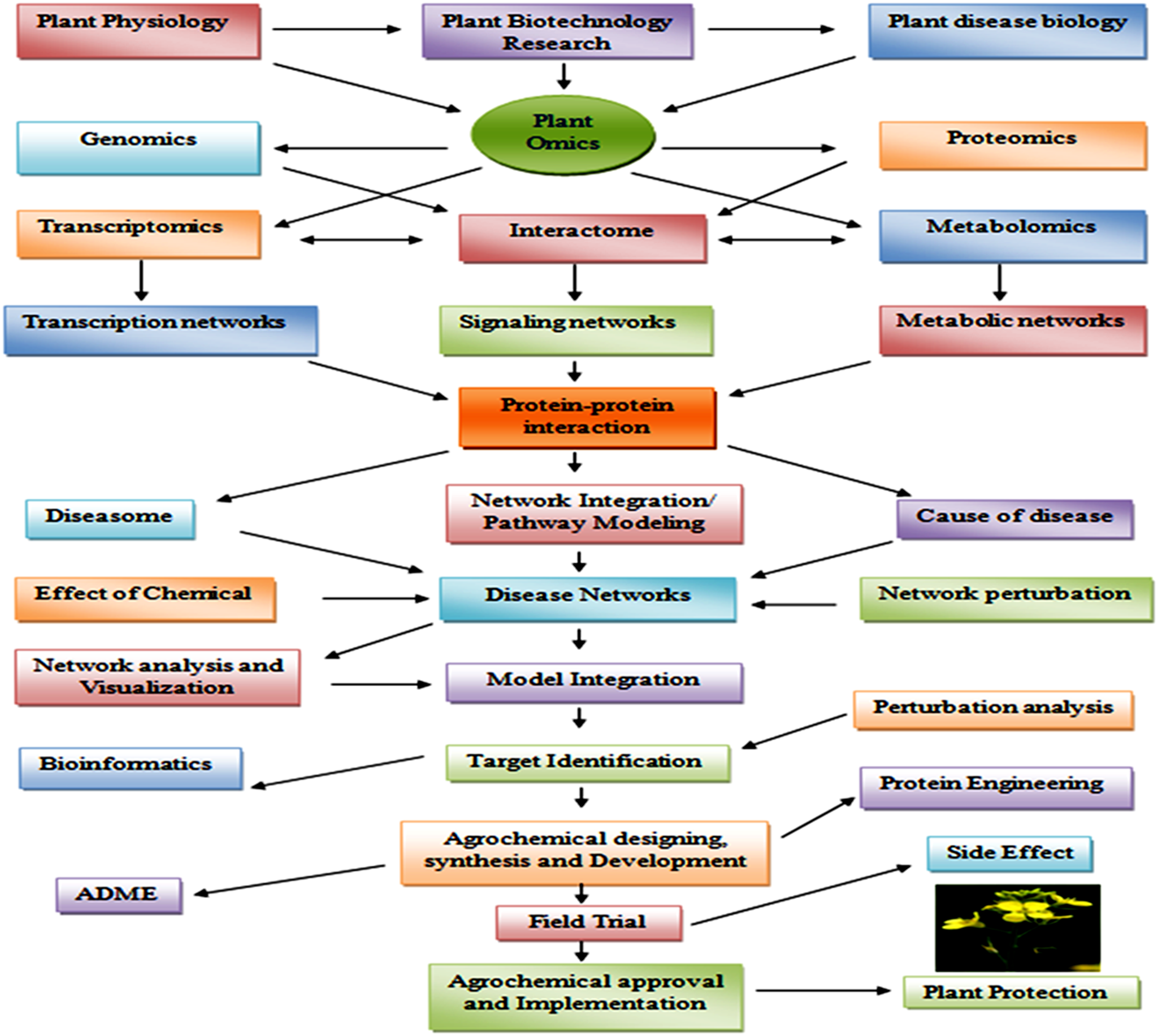
Summary of the Systems Biology approaches in crop plant protection.
Network biology for identification of disease associated genes or proteins
The pathogenesis of most diseases involves interaction of various proteins or genes. The network biology approach is being used extensively to identify the candidate genes/proteins responsible for various crop diseases. Topological analysis of biological network shows that the majority of the disease genes are nonessential and do not have a high degree of protein–protein interaction network and lie at boundary or periphery of the network, while essential genes encoding most of the hubs lie centrally to the network (Goh et al., 2007; Qi et al., 2006). These observations reveal the existence of disease-specific functional components.
The network measures and topological analysis will significantly help in deriving productive information from the complex network such as degree, clustering centrality, shortest path connectivity, and hubs in the network. This information further assists in designing experimentally or by computer simulation. In a outstanding work in humans, Lage et al. (2007) have applied such analysis to construct the interactions network of various genes and proteins. Based on network analysis, they found a total of 669 linkages, out of which 298 correctly ranked the disease causing proteins as top candidates. Network construction and analysis has great potential to identify large number of genes and proteins responsible for diseases and facilitate identification of important traits for agricultural productivity.
Pathway modeling
Pathway modeling and analysis are useful tools for understanding the inter-relation between different biological components (genes, proteins, metabolites) for identification of complex agricultural traits in crop plants. Theoretical analysis of biological pathway has a long history, and has been successfully applied to the analysis of metabolic pathway and physiological processes (Heinrich, 1985; Leclercq et al., 1983). Computational modeling of biological systems is becoming increasingly useful in many areas of biological sciences, including in the study of signaling pathways for identification of a growing number interactions within and between signaling pathways in the cell.
Computational models should represent the biological systems as accurately as possible and be able to mimic the behavior of the systems over a wide variety of conditions, including various abiotic and biotic stresses in crop plant systems. Collecting information from published literature and databases is one of the most important aspects in the development of computational models. However, often this is not sufficient. As these types of models become more complex, it will require experimental data for accurate validation and analysis; such data should include cellular concentration of the components, as well as the kinetic reactions for interactions between components. Systems biology graphical notations (SBGN) are available for modeling of biological systems by using different types of available computational tools (Kitano et al., 2005). SBGN facilitates the researcher with respect to symbols for receptor, gene, protein for the purpose of interlinking of cell components that provides better analysis and visualization of complex agricultural traits.
Networks for disease resistance
The molecular interaction network comprising of biomolecules associated with different agriculturally important traits/genes, which influence the network function from genotype to phenotype, is known as traits network. The inherent complexity in the molecular interaction networks results in various components yielding an emergent property essential for normal functioning. Recognition of the components required for specific functional properties and their perturbation results in alteration in the phenotypic response which results in complex diseases, are essential to identify desired traits (Altaf-Ul-Amin et al., 2014; De Vleesschauwer et al., 2014; Doncheva et al., 2012; Mackay et al., 2009; Xu et al., 2014).
Network-based approaches indicate that multiple component nodes may be involved in a disease state that further affect the functional multigeneic modules found in nature. For example, multiple target molecules of both plant and pathogen appear to be involved in important crop plant diseases such as Alternaria blight, which is caused by Alternaria brassicae and A. brassicicola and results in 10%–70% of yield losses in different parts of Northern India according to climatic conditions (Kumar et al., 2001; Pandey et al., 2001; Priyanka et al., 2013). There is a need of network-based approach for crop systems modeling and analysis, which provides way for development of novel disease management strategy.
Research studies based on topological analysis of disease network have suggested that disease genes lie on periphery of the network, which may be effectively targeted without much side effects on society and environment (Hase et al., 2009; Zhu et al., 2009). There can be multiple targets at different network levels that can be identified by network dynamics analysis and visualization (Moller 2001). So, identification of the target and designing of the potential agrochemical/peptides that can modulate the network response are important.
The task of network-based systems targeting has become very difficult as compared to targeting single genes/protein or trait, as this needs an in-depth knowledge of the regulatory dynamics of the network and availability of the accurate system parameters. The dynamics simulations of network response can be carried out by performing perturbation analyses using systems biology tool (Table 1), these approaches are continuously evolving with immense potential to discover agriculturally important traits and their behavior in desired conditions similar in health and diseases (Somvanshi et al., 2014).
Biofortification and nutraceutical development
The main objective of biofortification and nutraceutical development is to enrich the plant foods for essential micronutrients and proteins as plants grow naturally. It has been realized that biofortification of staple food crops would solve the malnutrition problem associated with rural poor. In addition, they play essential roles in development of functional food (Nutraceuticals) for nutrition, health, and well-being by the application of systems biology. Recently published studies demonstrated that much work is required for development of new varieties with enhanced nutritional qualities by using interdisciplinary approaches (Hefferon, 2015), to achieve the goal for providing the crops with additional health benefits on a global scale.
In our lab, we have successfully sequenced the transcriptome of finger millet (Eleusine coracana), Through systems biology approach, 82 unique calcium sensor genes were categorized into eight calcium sensor gene family viz., CaM and CaMLs, CBLs, CIPKs, PEPRKs, CRKs, CDPKs, CaMKs, and CCaMK. 12 genes were found from out of 82 that are diverse from the rice orthologs (Kumar et al., 2014; Singh et al., 2014). A total of 19 Ca2 + transporter genes were also identified using transcriptome data of developing spike of finger millet. The variation in amino acid sequence between finger millet and rice Ca2 + transporter were observed to be 33%–90%. High level of co-relation among the expression of EcCAX3 gene and the amount of calcium accumulated in spikes were also recorded (Singh et al., 2015). It will help in opening new vistas for biofortification of agriculturally important crops.
Systems Biology in Agriculture: Future Directions
The sequencing of big and complex genomes of crop plants, facilitated by new sequencing technologies, data integrations, modeling, visualization, and simulation approaches has provided new opportunities in filling the gap between genotype to phenotype for improving the crop productivity (Bevan et al., 2013; Gjuvsland et al., 2013). The functional interactions between various biomolecules such as nucleic acid, transcription factors, enzymes, proteins, and metabolites form the basis of the components for a molecular interaction pathway. The data and information with respect to such molecular interaction and their regulation from molecular to organism level is called as interactome. Interactome analysis enables insight into the link between genotype and phenotype in terms of biomolecular interaction pathways that can lead to detection of different type of diseases. Hence it is necessary to investigate various interacting partners and analyze the pathway for proper diagnosis of disease along with its mechanisms (Cantu et al., 2013; Sahu et al., 2014; Seo et al., 2011).
Current researchers applied such systems biology approaches to identify and visualize various candidate genes/traits of agriculturally importance, as well as to provide new biological insight in root, stem, leaf, fruit, and seed levels for agricultural productivity. Such systems biology approaches will be very useful, when we use diverse genomic and transcriptomic information to increase the accuracy with which phenotype can be predicted from genotype, thus accelerating crop improvement and helping to address disease management issues, crop productivity, and food security in future (Bevan et al., 2013; Higgins et al., 2010; Matsuoka et al., 2014).
Conclusion
Systems biology helps us to understand crops at systems level, which provide novel molecular insights for augmenting agricultural productivity in the future. The progress in systems biology is still ongoing, and available database resources are updated timely with new information. The software development program with improved algorithms is also running in various systems biology institutions and R and D divisions of software industries in the world, with the aim of better understanding of crop root, stem, leaf, fruit, and seed systems or as a whole crop systems using both Bottom up and Top-down approaches. The directional development in systems biology will help in understanding the temporal and spatial molecular perturbations during different stages/conditions of plant development. The molecular sketch thus obtained will enable us to engineer the crops for fulfilling the future demands of food and nutritional security.
Footnotes
Acknowledgments
We gratefully acknowledge the financial support by Biotechnology Information System Network (BTISNet), Department of Biotechnology (DBT), Govt. of India. Bioinformatics Centre (Sub DIC) at G. B. Pant University of Agriculture and Technology, Pantnagar, India is also duly acknowledged for providing research facilities.
Author Disclosure Statement
The authors declare that they have no competing financial interests.