
Abstract
Abstract
Culturomics is a new omics subspecialty to map the microbial diversity of human gut, coupled with a taxono-genomic strategy. We report here the description of a new bacterial species using microbial culturomics: strain S5T, ( = CSUR P1091 = DSM 28586) isolated from a stool specimen of a 25-year-old obese patient from Saudi Arabia. The strain S5T was a Gram-positive, strictly aerobic rod, which was motile by a polar flagellum, spore-forming, and exhibited catalase and oxidase activities. It grows optimally at 37°C, with a pH of 7.5 and 10% of NaCl. 16S rRNA gene-based identification revealed that strain S5T has 98.6% 16S rRNA sequence similarity with the reference O. oncorhynchi, phylogenetically the closest validated Oceanobacillus species. Here, we further describe the phenotypic characteristics of this organism and its complete genome sequence and annotation. The 5,388,285 bp long genome exhibits a G + C content of 37.24% and contains 5109 protein-coding genes and 198 RNA genes. Based on the characteristics reported here, we propose classifying this novel bacterium as representative of a new species belonging to the genus Oceanobacillus, Oceanobacillus jeddahense sp. nov. In a broader context, it is noteworthy that halophilic bacteria have long been overlooked in the human gut, and their role in human health and disease has not yet been investigated. This study thus further underscores the usefulness of the culturomics approach exploring the bacterial diversity of the gut.
Introduction
A
We have recently outlined a new method (taxono-genomics), including genomic data in a polyphasic approach in order to describe new bacterial species (Ramasamy et al., 2014). This strategy combines phenotypic characteristics with MALDI-TOF MS spectrum and genomic analysis (Lagier et al., 2012b; Mishra et al., 2012). Using this method, we isolated the strain S5T belonging to the genus Oceanobacillus. The first description of genus Oceanobacillus was made by Lu et al. (2001) and was then amended (Yumoto et al., 2005). These bacteria belong to the phylum Firmicutes within the Bacillaceae family. The Oceanobacillus genus currently includes 13 recognized species and two subspecies. These bacteria are Gram-positive, motile rods, obligate aerobic or facultative anaerobic, and obligate or facultative alkaliphilic.
Bacteria from the Oceanobacillus genus were essentially isolated from deep-sea sediment core (Lu et al., 2001; Kim et al., 2007; Yumoto et al., 2005), deteriorated mural paintings (Heyrman et al., 2003), salt fields (Lee et al., 2006), freshwater fish (Yumoto et al., 2005), algal mat (Romano et al., 2006), freshwater insects (Raats and Halpern, 2007), a Bacillus-dominated wastewater treatment system in Korea (Nam et al., 2008), fermented shrimp paste samples (Namwong et al., 2009), soy sauce production equipment (Tominaga et al., 2009), marine solar saltern (Lee et al., 2010), activated sludge in a bioreactor (Whon et al., 2010), traditional Korean fermented food (Hirota et al., 2013), fermented Polygonum indigo liquor sample (Yang et al., 2010), and, finally, a human stool sample (Roux et al., 2013).
Here, we used high salt culture conditions in order to cultivate halophilic bacteria from human faeces and a novel Oceanobacillus isolate, strain S5T ( = CSUR P1091 = DSM 28586), was isolated from a stool sample of a 25-year-old obese Saudi patient, as part of culturomics study of intestinal microflora, as a new representative of the genus Oceanobacillus.
Materials and Methods
Ethics and collection of the strain
The stool specimens were collected from a 25-year-old obese Saudi patient after defecation into sterile plastic containers, which were sampled and stored at −80°C until use. (Supplementary Table S1. Supplementary material is available online at www.liebertpub.com/omi). Informed and signed consent was obtained from the patient. The study and consent procedure were approved by the Ethics Committees of the King Abdulaziz University (King Fahd Medical Research Center, Saudi Arabia), under agreement number 014-CEGMR-2-ETH-P, and by the Institut Fédératif de Recherche 48 (Faculty of Medicine, Marseille, France), under agreement number 09-022. The salt concentration of the stool specimen was determined using a digital refractometer (Fisher Scientific, Illkirch, France) and the pH by using a pH-meter.
Isolation of the strain
Strain S5T was isolated in December 2013 using aerobic cultivation on a home-made culture medium consisting of a Columbia broth culture medium (Sigma-Aldrich, Saint-Quentin Fallavier, France) modified by adding (per liter): MgCl2 6H2O, 5 g; MgSO4 7H2O, 5 g; KCl, 2 g; CaCl2 2H2O, 1 g; NaBr, 0.5 g; NaHCO3, 0.5 g; glucose 2 g, and 10% of NaCl. The pH was adjusted to 7.5 with 10 M NaOH before autoclaving. Approximately 1 g of stool specimen was inoculated aerobically into 100 mL of our home-made liquid medium in flasks at 37°C, stirring at 150 rpm. Subcultures were conducted after 3, 10, 15, 20, and 30 days incubation. Serial dilutions of 10−1 to 10−10 were then performed in the home-made liquid culture medium and plated on agar medium. After 3 days' incubation at 37°C, all colonies that appeared were picked and streaked replant several times to obtain pure cultures.
Strain identification using MALDI-TOF and 16S rRNA sequencing
MALDI-TOF MS (matrix-assisted laser-desorption/ionization time-of-flight) protein analysis was carried out as previously described (Seng et al., 2009). The resulting twelve spectra of strain S5T were imported into the MALDI BioTyper software (version 2.0, Bruker) and analyzed using standard pattern matching (with default parameter settings) against the main spectra of over 6252 bacteria, including the spectra from the most closely related species: Oceanobacillus oncorhynchi CIP108867T, O. picturae CIP 108264T, O. profundus CIP 109535T, O. chironomi CIP 109536T, O. iheyensis CIP 107618T, and O. oncorhynchi subsp. incaldanensis CIP 109235T. The 16S rRNA PCR and sequencing were performed as previously described (Mourembou et al., 2015).
Growth conditions
The optimum growth temperature of the strain S5T was tested on our home-made solid medium by inoculating 105 CFU/mL of an exponentially growing culture incubated aerobically at 28°, 37°, 45°, and 55°C. The growth atmosphere was tested under aerobic conditions, in the presence of 5% CO2, and also in microaerophilic and anaerobic conditions created using GENbag microaer and GENbag anaer (BioMérieux, Marcy l'Etoile, France), respectively. The optimum NaCl concentration required for growth was tested at 0%, 0.5%, 1.5%, 5%, 7.5%, 10%, 15%, and 20% of NaCl. The optimum pH was determined by growth testing at pH 5, 6, 7, 7.5, 8. and 9.
Biochemical, sporulation, and motility assays
The commercially available Api ZYM, Api 20 NE, Api 50 CH strips (bioMérieux), supplemented by 10% NaCl (w/v), were used to characterize the biochemical properties of the strain according to the manufacturer's instructions. Strain sporulation was tested by thermic-shock at 60°C for 20 minutes. A fresh culture of the strain S5T was observed under DM1000 photonic microscope (Leica Microsystems, Nanterre, France) at 100X to assess the motility of the bacteria. The colony's surface was observed on our home-made culture medium after 24 h incubation under aerobic conditions at 37°C.
Antibiotic susceptibility testing
Susceptibility to antibiotics was tested using antibiotic disks (B. Braun Medical SAS, Boulogne, France) containing the following antibiotics: doxycycline, rifampicin, vancomycin, nitrofurantoin, amoxicillin, erythromycin, ampicillin, ceftriaxone, ciprofloxacin, gentamicin, penicillin, trimethoprim/sulfamethoxazole, imipenem, and metronidazole.
Microscopy and Gram test
Transmission electron microscopy of the strain S5T was performed after negative staining using a Morgani 268D (Philips, Verdun, France) at an operating voltage of 60kV. The Gram stain was performed using the colour Gram 2 kit (Biomerieux) and observed using a DM1000 photonic microscope (Leica Microsystems).
DNA extraction
DNA extraction of the strain S5T was conducted on ten Petri dishes of fresh culture that were harvested and resuspended in 4X 100 μL of TE buffer. 200 μL of this suspension was then diluted in 1 mL TE buffer for lysis treatment that included 30 min incubation with 2.5 μg/μL of lysozyme at 37°C, followed by overnight incubation with 20 μg/μL of proteinase-K at 37°C. Extracted DNA was then purified using three successive phenol-chloroform extractions and ethanol precipitation at −20°C overnight. After centrifugation, the DNA was resuspended in 160 μL TE buffer. The yield and concentration was measured using a Quant-it Picogreen kit (Invitrogen, Villebon sur Yvette, France) on the Genios-Tecan fluorometer.
Genome sequencing and assembly
Genomic DNA (gDNA) of the strain S5T was sequenced using MiSeq Technology (Illumina Inc, San Diego, CA, USA) with the mate pair strategy. The gDNA was barcoded in order to be mixed with 11 other projects with the Nextera Mate Pair sample prep kit (Illumina). The gDNA was quantified by a Qubit assay using the high sensitivity kit (Life Technologies, Carlsbad, CA).The mate pair library was prepared with 1 μg of genomic DNA using the Nextera Amate pair Illumina guide. The gDNA sample was simultaneously fragmented and tagged with a mate pair junction adapter. The profile of the fragmentation was validated on an Agilent 2100 BioAnalyzer (Agilent Technologies Inc., Santa Clara, CA) with a DNA 7500 labchip. The DNA fragments were sorted by size from 1 kb up to 10 kb. No size selection was performed and only 14 ng of tagmented fragments were circularized. The circularized DNA was mechanically sheared to small fragments with an optimal at 696 bp on the Covaris device S2 in microtubes (Covaris, Woburn, MA, USA).The library profile was visualized on a High Sensitivity Bioanalyzer LabChip (Agilent Technologies. Inc.).
The libraries were normalized at 2 nM and pooled. After a denaturation step and dilution at 10 pM, the pool of libraries was loaded onto the reagent cartridge and then onto the instrument along with the flow cell. The automated cluster generation and sequencing run were performed in a single 42-hour run in a 2 × 251-bp. Illumina reads were trimmed using Trimmomatic (Lohse et al., 2012), then assembled through Spades software (Bankevich et al., 2012; Nurk et al., 2013). Contigs obtained were combined together using SSpace (Boetzer et al., 2011) and Opera software v1.2 (Gao et al., 2011), with assistance from GapFiller V1.10 (Boetzer et al., 2012) to reduce the set. Some manual refinements using CLC Genomics v7 software (CLC bio, Aarhus, Denmark) and homemade tools improved the genome.
Genome annotation
Non-coding genes and miscellaneous features of the strain S5T were predicted using RNAmmer (Lagesen et al., 2007), ARAGORN (Laslett et al., 2004), Rfam (Griffiths-Jones et al., 2003), PFAM (Punta et al., 2012), and Infernal (Nawrocki et al., 2009). Coding DNA sequences (CDSs) were predicted using Prodigal (Hyatt et al., 2010) and functional annotation was achieved using BLAST+ (Camacho et al., 2009) and HMMER3 (Eddy 2009) against the UniProtKB database (The UniProt Consortium, 2011).
Genome comparison
To estimate the mean level of nucleotide sequence similarity at genome level between the strain S5T and the four closest species with an available genome, we used the Average Genomic Identity of Orthologous gene Sequences (AGIOS), an in lab pipeline. Briefly, this pipeline combines the Proteinortho software (with the following parameters: e-value 1e-5, 30% identity, 50% coverage, and algebraic connectivity of 50%) for detecting orthologous proteins between genomes compared pairwise, and retrieves the corresponding genes and determines the mean percentage of nucleotide sequence identity between orthologous ORFs using the Needleman-Wunsch global alignment algorithm (Ramasamy et al., 2014).
Results
Phenotypic and biochemical characterization
The colonies of the strain S5T were circular, grayish, shiny, and smooth with a diameter of 2–5 mm. Cells stained Gram-positive and were rod shaped (Fig. 1). They were motile by polar flagella, spore-forming, had a length of 2–4 μm and a diameter of 0.5 μm, and generally occurred singly or in pairs (Fig. 2). Strain S5T was mesophilic and grew at temperatures ranging from 15° to 45°C, with optimum growth at 37°C. The isolate required NaCl for growth and grew at salinity ranging from 0.5 to 20% of NaCl (optimum at 10%). The optimum pH for growth was 7.5 (range between pH 5 to 9). The strain S5T was aerobic and also grew in microaerophilia and in the presence of 5% CO2 but did not grow in an anaerobic atmosphere.
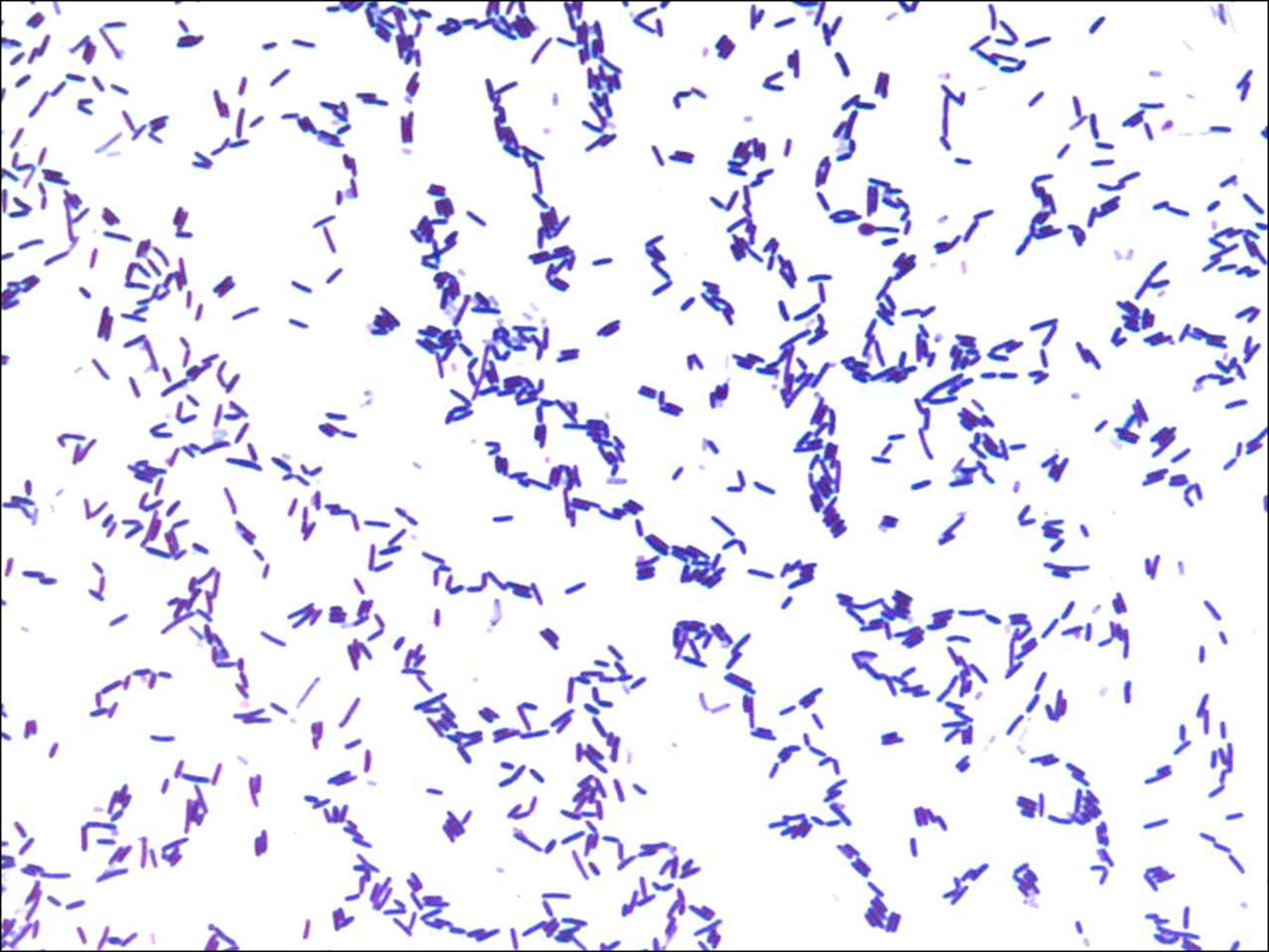
Gram staining of Oceanobacillus jeddahense strain S5T.

Transmission electron microscopy of Oceanobacillus jeddahense strain S5T. The scale bar represents 500 nm.
The strain S5T exhibited catalase and oxidase activity. Using an Api ZYM strip and Api 20 NE, a negative reaction was observed only for alkaline phosphatase, β-galactosidase, N-acetyl-β-glucosaminidase, and urease. Positive reactions were observed for nitrate reduction. Using API 50CH strip and Api 20E at 37°C, negative reactions were obtained for D-lactose, L-arabinose, D-galactose, and D-ribose. Positive reactions were obtained for D-glucose, D-fructose, D-mannose, D-mannitol, D-maltose, and D-sucrose. Phenotypic characteristics were compared to those of the most closely related species (Table 1). Antimicrobial susceptibility testing demonstrate that the strain S5T was susceptible to doxycycline, rifampicin, vancomycin, nitrofurantoin, amoxicillin, erythromycin, ampicillin, ceftriaxone, ciprofloxacin, gentamicin, penicillin, and imipenem, but resistant to trimethoprim/sulfamethoxazole and metronidazole.
Oceanobacillus oncorhynchi (Yumoto et al., 2005), Oceanobacillus iheyensis (Lu et al., 2001), Oceanobacillus sojae (Tominaga et al., 2009), Oceanobacillus massiliensis (Roux et al., 2013), Oceanobacillus chironomi (Raats et al., 2007), Oceanobacillus picturae (Lee et al., 2006), Oceanobacillus indicireducens (Hirota et al., 2013).
na = data not available; v = variable reaction; w = weak.
MALDI-TOF and phylogenic analysis
MALDI-TOF results showed that the spectra we obtained for strain S5T did not match with one of our database (Bruker constantly increment with our data), suggesting that our isolate was a new species. We added the spectrum from strain S5T to our database (Fig. 3). In contrast, PCR-based identification of the16S rRNA of the strain S5T (HG931338) demonstrated 98.6% 16S rRNA gene sequence homology with reference O. oncorhynchi (DQ089679), the phylogenetically closest validated Oceanobacillus species (Fig. 4). This value is under the threshold that allows for identification of new species, establish by Stackebrandt and Ebers (2006). Consequently, our strain is considered as a new species, named Oceanobacillus jeddahense. Finally, the gel view demonstrates the spectral differences with other members of the genus Oceanobacillus (Fig. 5).

Reference mass spectrum from Oceanobacillus jeddahense strain S5T. Spectra from 10 individual colonies were compared and a reference spectrum was generated.

Phylogenetic tree highlighting the position of Oceanobacillus jeddahense strain S5T relative to other type strains within the genus Oceanobacillus. GenBank accession numbers are indicated in parentheses. Sequences were aligned using CLUSTALW, and phylogenetic inferences obtained using the maximum-likelihood method within the MEGA software. Halobacillus litoralis was used as outgroup. The scale bar represents 0.005% nucleotide sequence divergence.
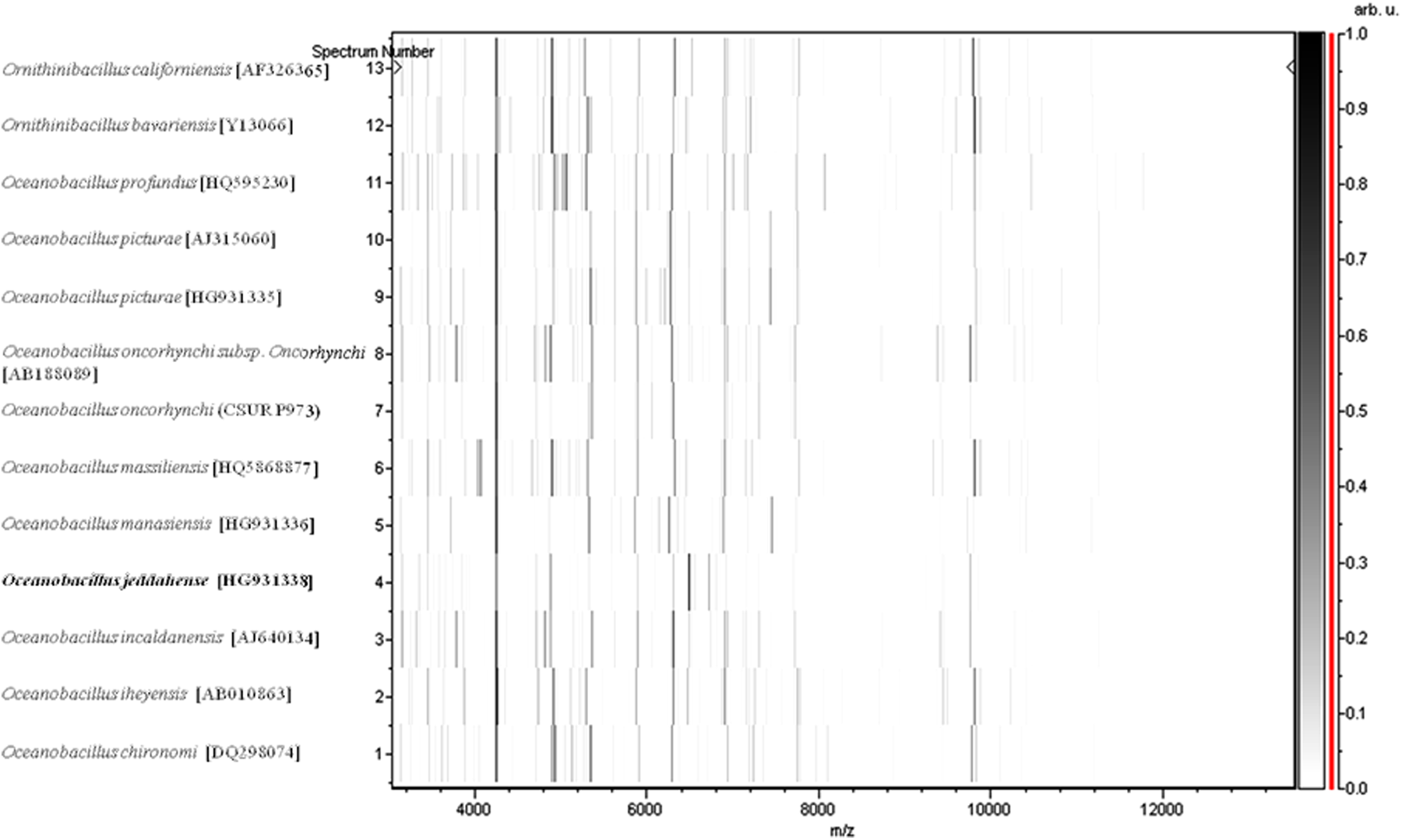
Gel view comparing Oceanobacillus jeddahense strain S5T to other species within the genus Oceanobacillus and Ornithinibacillus. The gel view displays the raw spectra of loaded spectrum files arranged in a pseudo-gel like look. The x-axis records the m/z value. The left y-axis displays the running spectrum number originating from subsequent spectra loading. Peak intensity is expressed by a gray scale scheme. The color bar and the right y-axis indicate the relation between the colour a peak is displayed with and the peak intensity in arbitrary units. Displayed species are indicated on the left.
Genome information and properties
The O. jeddahense strain S5T genome was sequenced as part of culturomics study aiming to isolate all bacterial species colonizing the human gut (Lu et al., 2001) and because of its phylogenetic affiliation to the Oceanobacillus genus. Strain S5T represents the fourth genome sequenced in the Oceanobacillus genus and the first genome of O. jeddahense sp. nov. The O. jeddahense strain S5T genome contains 5,388,285 bp with a G + C content of 37.24% (Fig. 6) and consists of 11 scaffolds with 12 contigs.
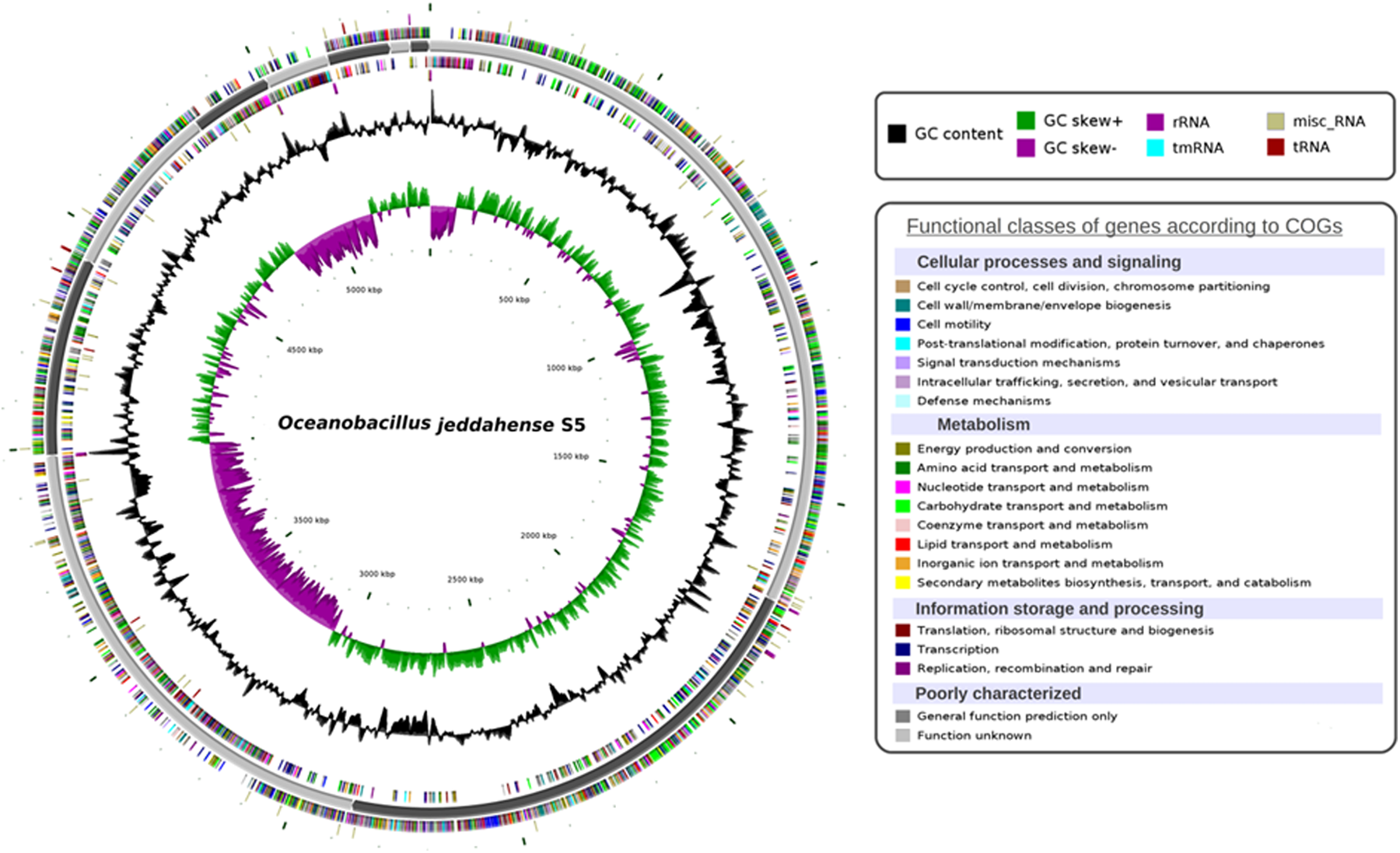
Circular representation of the Oceanobacillus jeddahense strain S5T chromosome. Circles from the center to the outside: GC skew (green/purple), GC content (black), RNA (dark red, purple, blue, beige) on forward strand, genes on forward strand colored by COGs categories, scaffolds in alternative grays, genes on reverse strand colored by COGs, RNA (dark red, purple, blue, beige) on reverse strand.
The draft genome (Genbank accession number CCDM010000001.1) was shown to encode at least 198 predicted RNA including 17 rRNA, 65 tRNA, 1 tmRNA, and 115 miscellaneous RNA (Table 2). 5109 protein-coding genes were also identified, representing a coding capacity of 4,382,541 bp (coding percentage: 81.3%). Of these genes, 860 (16.83%) were found to be putative proteins and 1109 (21.71%) were assigned as hypothetical proteins. Moreover, 4303 genes matched against at least one sequence in the Clusters of Orthologous Groups (COGs) database (Tatusov et al., 2000) with BLASTP default parameters. Table 3 presents the distribution of genes in COGs functional categories.
The total is based on either the size of the genome in base pairs or the total number of protein coding genes in the annotated genome.
Genome comparison
Brief comparisons were made with Oceanobacillus iheyensis strain HTE831 (BA000028), which is currently the closest available annotated sequenced genome. The draft genome sequence of O. jeddahense strain S5T is bigger in size than the O. iheyensis (respectively, 5.39 Mb vs. 3.63 Mb). O. jeddahense strain S5T exhibit more genes (5217 genes compared to 3593 genes) and also has a similar ratio of genes per Mb (967.9 genes/Mb compared to 989.8 genes/Mb).The G + C content is close to Bacteroides intestinalis (37.24% and 35.7%, respectively). The gene number difference (in percentage) related to each COG categories between O. jeddahense and O. iheyensis is presented in Table 3. The maximum difference relates to the COG “Transcription” which does not exceed 2.32%. In terms of the other categories, the two species are highly similar.
In addition, a whole genome comparison was performed between O. jeddahense, O. massiliensis, O. kimchii (uid 199920), O. iheyensis (uid 57867), and Halobacillus halophilus (uid 162033). At the time of analysis, only three whole genome sequences of Oceanobacillus were available at NCBI. Among the species of Oceanobacillus spp., the draft genome of O. jeddahense strain S5T (5.4 Mb) is the largest, followed by O. kimchii (3.8 Mb), O. iheyensis (3.6 Mb), and O. massiliensis (3.5 Mb), respectively. H.halophilus has a genome size of 4.1 Mb.
The AGIOs values in Oceanobacillus genus for the four studied genomes range from 67.70% to 89.62%. The AGIOs values for O. jeddahense strain S5T and other species of Oceanobacillus range from 67.70% (O. massiliensis) to 69.85% (O. iheyensis and O. kimchii), thus confirming its new species status (Table 4). The orthologous genes shared between the O. jeddahense strain S5T and other compared species are roughly half compared to the total proteins in the strain S5T, suggesting a high number of hypothetical proteins in this genome.
Discussion
This work describes the isolation of a moderately halophilic bacterium associated with the human microbiota using culture-based detection methods. The culturomics approach was applied to render a dynamic description of new bacteria species and genera from the human gut, including samples from various geographical origins. In this way, several new halophilic bacteria species were recently isolated in our laboratory, including two new species recently described (Khelaifia et al., 2015; Seck et al., 2015). Here, we report the phenotypic characterization and genomic description of a new halophilic bacterium isolated from a stool specimen collected from 25-year-old obese patient from Saudi Arabia. This strain was deposited in two different collections of microorganisms, Deutsche Sammlung von Mikroorganismen und Zellkulturen (DSMZ) and the Collection de Souches de l'Unité des Rickettsies (CSUR). After this study, O. jeddahense was isolated from three different stool specimens and one duodenum simple (unpublished data), confirming it is not uncommon in the human microbiota.
This work supports the usefulness of the culturomics approach exploring the bacterial diversity of the gut and its new approach proposed to describe the new species of microorganism based on a phenotypic characterization and genomic description. Indeed, as this work demonstrates, halophilic bacteria have long been overlooked in the human gut, and their role in the human health and disease has not yet been investigated. Culturomics is so far the only approach that has looked at halophilic organisms colonizing the human gut. It should be noted that this work does not intend to describe the medical or biotechnological importance of this new bacterium; it aims to extend the repertoire of microorganisms that colonize colonise the human gut in order to better understand the composition of its microflora.
Conclusion
Based on phenotypic, phylogenetic, and genomic analyses, we formally propose the creation of Oceanobacillus jeddahense sp. nov., represented here by the type strain S5T. The strain has been isolated from a stool specimen of a Saudi male individual suffering from morbid obesity. Several other bacterial species as yet undescribed were also isolated from different stool specimens as part of the culturomics study using different culture conditions, suggesting that the human intestinal microbiota remains partially unknown and its diversity has yet to be fully explored.
Description of Oceanobacillus jeddahense sp. nov.
Oceanobacillus jeddahense strain S5T (jed.dah.en'.se L.gen. neutr. n.; from Jeddah, Saudi Arabia, where the specimen was obtained from an obese Saudi patient). Growth of O. jeddahense strain S5T occurred between 15° to 45°C (optimal growth at 37°C), in aerobic conditions with a pH ranging from pH 5 to 9 (optimum pH 7.5). It required NaCl for growth, ranging from 0.5% to 20% (optimum at 10%). Colonies of O. jeddahense strain S5T were circular, grayish, shiny, and smooth, with a diameter of between 2 and 5 mm. Cells are Gram-positive and rod-shaped, measuring 2 to 4 μm in length and 0.5 μm in diameter. They were motile because of polar flagella, and spore-forming.
The O. jeddahense strain S5T exhibited catalase and oxidase activities. Using an Api ZYM strip and Api 20 NE, negative reactions were observed for alkaline phosphatase, β-galactosidase, N-acetyl-β-glucosaminidase, and urease, and positive reactions were observed for nitrate reduction. Using API 50CH strip and Api 20E, negative reactions were obtained for D-lactose, L-arabinose, D-galactose and D-ribose and positive reactions were obtained for D-glucose, D-fructose, D-mannose, D-mannitol, D-maltose and D-sucrose. O. jeddahense strain S5T was susceptible to doxycycline, rifampicin, vancomycin, nitrofurantoin, amoxicillin, erythromycin, ampicillin, ceftriaxone, ciprofloxacin, gentamicin, penicillin, and imipenem, but resistant to trimethoprim/sulfamethoxazole and metronidazole. The G+C% content of the genome is 37.24%. The 16S rRNA and genome sequences are deposited in GenBank under accession numbers HG931338 and CCDM010000001.1 respectively. The habitat of the microorganism is the human digestive tract. The type strain of O. jeddahense S5T ( = CSUR P1091 = DSM 28586) was isolated from a stool specimen of a Saudi male suffering from morbid obesity.
Footnotes
Acknowledgments
This project was funded by the Deanship of Scientific Research (DSR), King Abdulaziz University, Jeddah, under grant no. (3-140-1434-HiCi). The authors, therefore, acknowledge DSR technical and financial support with thanks. This study was also funded by the “Fondation Méditerranée Infection”.
Author Disclosure Statement
The authors declare that there is no financial conflict of interest.
Abbreviations
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.