
Abstract
Abstract
Vertical transmission of human immunodeficiency virus (HIV) remains a major global health problem. We assessed the association of mannose binding lectin (MBL) deficiency and vertical transmission of HIV. Novel diagnostics would be a major breakthrough in this regard. MBL is a liver-derived protein and a key component of the innate immune system. MBL levels may be classified as normal, intermediate, or deficient in the plasma and can use MBL2 haplotypes as a proxy. These haplotypes comprise polymorphisms in the MBL2 gene and promoter region and are known to result in varying levels of MBL deficiency. MBL deficiency can be defined as presence of A/O and O/O genotypes in the mothers and their children. MBL deficiency leads to defective opsonization activities of the innate immune system and increased susceptibility to several infections, including HIV-1. We determined the prevalence of MBL deficiency, using MBL2 haplotypes among 622 HIV-positive Zimbabwean mothers and their children aged 9–18 months old, in relation to the HIV-1 vertical transmission risk. The median age of the mothers was 30 (26–34, interquartile range [IQR]) years, and the babies' median age was 13 (11–15, IQR) months old at the time of enrollment. From the sample of 622 mothers who were HIV-1 infected, 574 babies were HIV negative and 48 were HIV-1-positive babies, giving a transmission rate of 7.7%. MBL2 normal structural allele A and variants B (codon 5 A>G), C (codon 57 A>G), and promoter region SNPs −550(H/L) and −221(X/Y) were detected. Prevalence of haplotype-predicted MBL deficiency was 34% among the mothers and 32% among the children. We found no association between maternal MBL2 deficiency and HIV-1 transmission to their children. We found no difference in the distribution of HIV-1 infected and uninfected children between the MBL2 genotypes of the mothers and those of the children. Taken together, the present study in a large sample of mother–infant pairs in Zimbabwe adds to the emerging literature and the hypothesis that MBL2 variation as predicted by haplotypes does not influence the vertical transmission risk for HIV. Research from other populations from the African continent is called for to test this hypothesis further.
Introduction
R
In children between the ages 6 and 24 months, MBL plays an important role in the control of infectious microorganisms as it is during this period in which the maternal protective IgG antibodies have waned and the child's immune system is still very immature (Dommett et al., 2006). The normal MBL2 allele is designated as A, and point mutations in the MBL2 gene result in presence of variant alleles, which encode three different structural variants designated B (codon 54, rs1800450 A>G), C (codon 57, rs1800451 A>G), and D (codon 52, rs5030737 T>C) (Madsen et al., 1995, 1998). These mutations affect the structural integrity of MBL protein, resulting in a 2-fold to 100-fold decrease in circulating MBL serum/plasma levels (Madsen et al., 1995, 1998). In addition to mutations in the MBL2 gene, point mutations in the MBL2 promoter region at positions H/L (−550, rs11003125 G>C), X/Y (−221, rs7096206 C>G), and in the 5′ untranslated region of exon 1 at position PQ (+4, rs7095891 T>C) have been well documented (Madsen et al., 1995, 1998). These variants lead to a 10% reduction of functional MBL in individuals that are heterozygous for defective alleles compared with the functional MBL found in individuals with two functional alleles (Summerfield et al., 1995). The SNPs at the promoter region are reported to be in strong linkage disequilibrium with SNPs at the exon 1 position, giving rise to different haplotypes classified into three groups as high or normal serum MBL producers (YA/YA, YA/XA), intermediate levels (XA/XA, YA/YO), and deficient levels (XA/YO, YO/YO) (Garred et al., 2003; Vengen et al., 2012).
Several studies have investigated the role of MBL2 genetic variants in HIV transmission from mothers to their children (Amoroso et al., 1999; Arraes et al., 2006; Boniotto et al., 2000, 2003; Kuhn et al., 2006; Mangano et al., 2008; Mhandire et al., 2014; Zupin et al., 2016); as reviewed by Israels et al. (2012), the findings are conflicting (Israels et al., 2012). Some have reported MBL deficiency assessed by the presence of MBL2 genetic variants to be a risk factor for HIV vertical transmission (Arraes et al., 2006; Kuhn et al., 2006; Mangano et al., 2008), but others reported no association (Boniotto et al., 2000; Mhandire et al., 2014; Zupin et al., 2016). Zimbabwe is one of the countries in Southern Africa bearing a huge burden of mother to child HIV transmission (5–15%) (UNAIDS, 2013); it is therefore of public health importance to determine host genetic factors contributing to mother to child transmission (MTCT). The translation of knowledge and multiomics research are critical and urgently needed within the African context in view of the high diseases burden (Ahmed et al., 2014; Bosch et al., 2014; Dandara et al., 2014; ElRakaiby et al., 2014). This study aimed to determine the role of MBL2 genetic variants, promoter region variants, and MBL2 haplotypes in HIV-1 infection and vertical transmission in a subset of the blood samples collected during the national prevention of mother to child transmission (PMTCT) survey. The survey was carried out in 2012 by the Centre for Sexual Health and HIV/AIDS Research (CESHAAR) in Zimbabwe.
Materials and Methods
Study design and study population
This was a laboratory-based study and used dried blood spot samples that were collected as part of a national cross-sectional survey (McCoy et al., 2015a, 2015b) on PMTCT carried out by the CESHAAR Research Centre, Harare Zimbabwe. The primary study was entitled Evaluation of Zimbabwe's Accelerated PMTCT of HIV Program (Buzdugan et al., 2012; McCoy et al., 2015a, 2015b). This article reports the investigations on MBL2 genotyping on a subset of the dried blood samples collected during this national PMTCT survey (Fig. 1). Details of the PMTCT national survey, including blood sample collection, screening procedures, and the study population, have been described in detail elsewhere (Buzdugan et al., 2012). Briefly, the PMTCT survey was undertaken in September 2012 and targeted women aged 16 years and above who had children between 9 and 15 months old (McCoy et al., 2015a, 2015b). The survey examined the uptake of services and behaviors in the PMTCT in Zimbabwe and determined the factors associated with MTCT and maternal antiretroviral therapy (ART) or antiretroviral (ARV) prophylaxis. Dried blood spots were collected from 8800 babies and their biological mothers (McCoy et al., 2015a, 2015b).
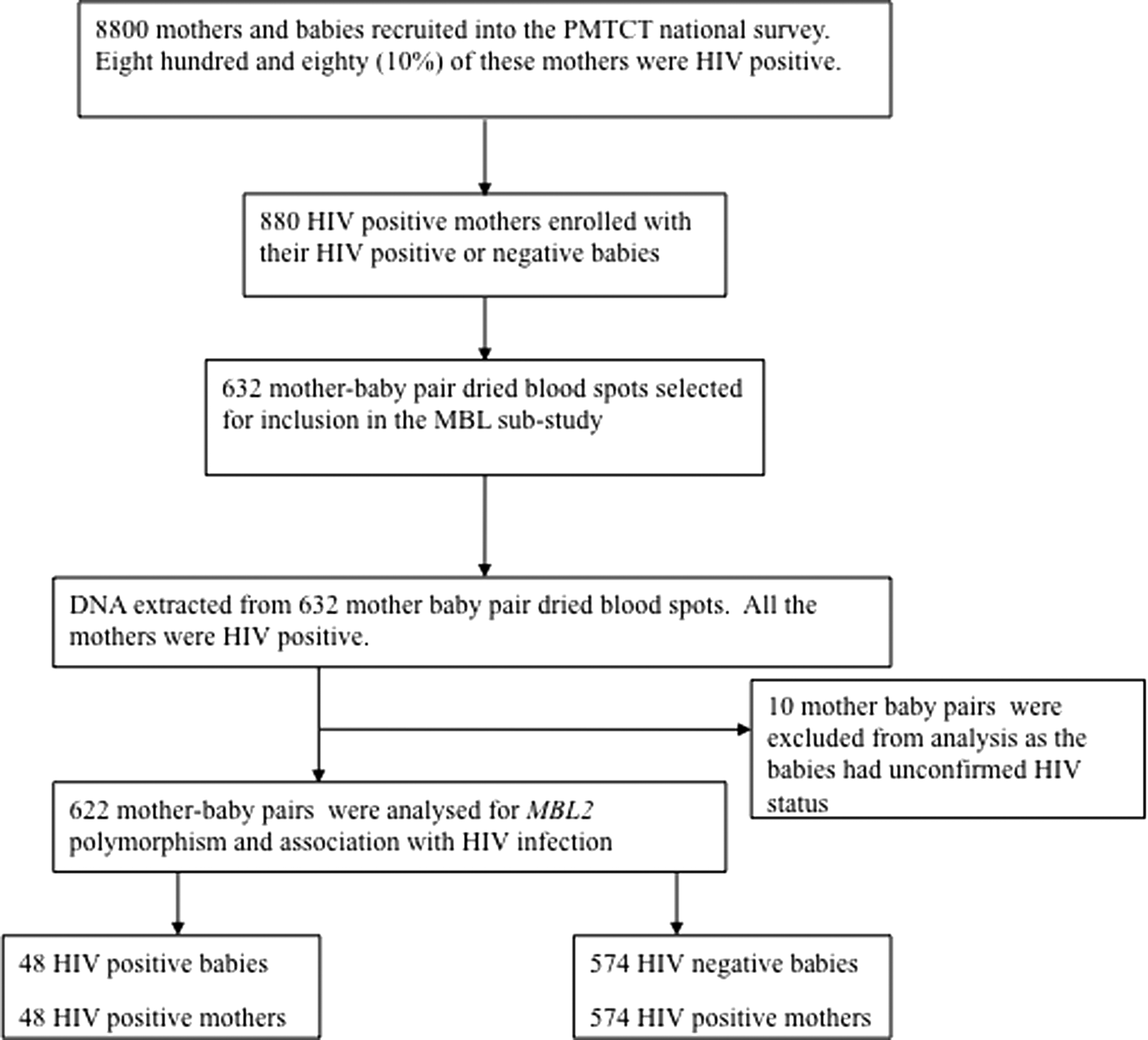
Participant Flow Chart showing the recruitment procedures for the PMTCT survey. Eight thousand eight hundred mother–baby pairs were enrolled in the national PMTCT survey, and 880 mothers were HIV positive. The 632 mother–baby pair dried blood spots selected for inclusion in the MBL substudy had adequate left over sample for DNA analysis. DNA was extracted from 632 mother–baby pair dried blood spots for MBL2 genotype analysis by the pyrosequencing method. Ten mother–baby pairs were excluded from analysis because of incomplete information. HIV, human immunodeficiency virus; MBL, mannose binding lectin; PMTCT, prevention of mother to child transmission.
Sample size calculation
The exposure of interest was MBL deficiency (using MBL2 haplotype as a proxy) among HIV-infected mothers, and the primary outcome was infant HIV status. Transmission rates range from 5% to 20% for mother–infant pairs on ART (UNAIDS, 2013). Assuming an alpha of 0.05 and beta of 0.90, 95% confidence interval, and 10% infant HIV status among the infants born from mothers with normal MBL2 genotype, we needed a sample size, among the exposed population, of 572 mother–infant pairs, 1144 individual blood spots in total. Fifty-eight (10%) more samples were selected to allow for dried blood spots that would not give adequate DNA for PCR, to give a final sample size of 632 mother baby pairs calculated using Epi Info 7 program (CDC, 2015). DNA was extracted from all 632 mother–baby pairs (Fig. 1). Complete data set on HIV status and MBL2 genotype was available on 622 mother–baby pairs used in the final analysis.
Ethics statement
The CESHAAR PMTCT study was approved by the National Research Ethics Committee of the Medical Research Council of Zimbabwe (MRCZ/A/1655), University of California and University of London Human Research Ethics Committees. The MBL2 substudy was also approved by the National Research Ethics Committee of the Medical Research Council of Zimbabwe (MRCZ/A/1770) and University of the Witwatersrand Human Research Ethics Committee (M130348). All the mothers who took part in the main PMTCT National Survey gave written informed consent for specimen collection, storage, and future laboratory studies.
Laboratory procedures
Collection of dried blood spots
Venous blood from the heel of the child and a finger of the mother was collected on filter papers (PerkinElmer 226, Health Sciences). Blood was collected on five designated spots on the filter papers. The filter papers were air-dried overnight and then packed the next day in zip lock bags with desiccant granules to prevent humid formation and keep the spots dry. The zip lock bags were kept at room temperature until analysis of the spots.
HIV testing
All the mothers were initially tested for HIV antibodies using EIA HIV test kits (OyToilette 3, FIN-01720; AniLabsystems Ltd). All the mothers that tested HIV positive on the initial screening tests were confirmed using Enzygnost Anti-HIV ½ Plus ELISA (Dade Behring), and indeterminate results were resolved by Western Blot. All the children as they were below 2 years of age were tested using HIV polymerase chain reaction test kits (Roche Amplicor HIV-1 DNA Test, version1.5) (McCoy et al., 2015b).
DNA extraction
Genomic DNA was extracted from dried blood spots using SIGMA Extract and Amp kit. Briefly, two 3 mm punches were made for each dried blood sample and incubated in 20 μL Lysis solution (SIGMA) at 75°C for 30 min. Subsequently, 180 μL of Neutralizing solution (SIGMA) was added and mixed by pipetting in the plate wells and centrifuged for 1 min at 1000 rpm. The extracted DNA quantified using the Qubit kit according to the manufacturer's instructions and stored at 4°C.
Pyrosequencing method
Amplification of MBL2 genes by Initial general PCR
DNA was amplified by PCR using the following reaction mixture (per sample): 41.125 μL Milli-Q water, 0.625 μL dNTPs at 10 mM each (from Pharmacia Biotech), 0.25 μL of 5U/μL HotStar DNA Taq Polymerase Enzyme (QIAGEN), 1 μL of 20 pmol of MBL2 Exon 1, H/L and Y/X labeled forward and reverse PCR primer (DNA Technology), 5 μL of 10 × HotStar PCR buffer, and a 2 μL sample DNA extract. A positive control (a Danish DNA sample of known MBL2 genotype) and a negative control (no DNA added) were included in each PCR run. The oligonucleotide primer sequences used for MBL2 genotyping are as shown (Supplementary Table S1). The following thermocycling program was used: an initial denaturation step at 95°C for 15 min, followed by 38 cycles of 95°C for 30 sec, 53°C for 30 sec, and 72°C for 30 sec; the reaction was completed by a final elongation/extension step at 72°C for 10 min. The PCR was expected to produce one specific amplicon of ∼240 bp; this was evaluated by agarose gel electrophoresis. This PCR product was used for the MBL2 pyrosequencing reaction.
Visualization of PCR products by agarose gel electrophoresis
The PCR products were separated by a 2% agarose gel (Invitrogen) electrophoresis run in an electrophoresis chamber (Sub-cell Biorad) at 150V, 400 mA for 32 min, including the positive and negative control samples. The gels were then carefully transferred to the UV Transilluminator (Syngene Synoptics Ltd) where a photograph of the gel was taken. Samples that showed the required MBL2 band were used for pyrosequencing (Fakruddin et al., 2012; Nyren 2007; Ronaghi 2001; Roos et al., 2006; Vengen et al., 2012)
Pyrosequencing reaction
In preparation for the pyrosequencing reaction, 25 μL of biotinylated PCR product was mixed with 3 μL of streptavidin-coated magnetic beads (Streptavidin Sepharose High performance; GE Healthcare Life Sciences), 37 μL of binding buffer (Qiagen), and 15 μL Milli-Q water. This mixture was agitated for 10–20 min at 1000 rpm on a plate shaker to allow the biotinylated PCR product to bind to the streptavidin sepharose beads. Subsequently, immobilized DNA was transferred, using a PSQ 9 magnetic sample prep tool (Pyrosequencing; Biotage), through a series of washing steps and finally released into a primer plate (Pyromark Q96; Qiagen) containing 39 μL annealing buffer (Qiagen), 1 μL of 16 pmol/μL of each specific pyrosequencing primer that is, Exon1, HL, and YX (DNA Technology) (Supplementary Table S1). This plate was incubated for 2 min at 80°C followed by cooling in the pyrosequencing machine (Pyromark Q96; Qiagen) for 10 min. Enzymes (DNA polymerase, ATP sulfurylase, luciferase, and apyrase), luciferine, and adenosine-5-phoshosulphate (APS, Pyromark Q96 cartridge reagents kit; Qiagen) were added to the cartridge wells according to instructions from the installed pyro program.
MBL2 genotypes and haplotype functional groups
Participants were classified into the different MBL2 Exon 1 genotypes, normal MBL2 allele designated as A and point mutations in the MBL2 gene and the variants designated as B (codon 54, rs1800450 A>G), C (codon 57, rs1800451 A>G), and D (codon 52, rs5030737 T>C) and the different combinations of variants (Madsen et al., 1995, 1998). Participants were classified into six MBL2 haplotype functional groups, which were combined into three groups for statistical analysis: haplotypes that give normal plasma MBL levels (YA/YA, YA/XA), intermediate levels (XA/XA, YA/YO), and deficient levels (XA/YO, YO/YO) (Garred et al., 2003; Vengen et al., 2012). We neither had plasma nor serum samples to determine the plasma MBL concentration for our study population, but used MBL2 genotypes as a proxy for plasma MBL levels (Amoroso et al., 1999; Arraes et al., 2006; Boniotto et al., 2000, 2003; Crovella et al., 2005; Dzwonek et al., 2006; Kuhn et al., 2006; Mangano et al., 2008; Singh et al., 2008) and to classify individuals into normal plasma MBL levels, intermediate levels, and deficient levels (Amoroso et al., 1999; Arraes et al., 2006; Boniotto et al., 2000, 2003; Crovella et al., 2005; Dzwonek et al., 2006; Kuhn et al. 2006; Mangano et al. 2008; Singh et al., 2008).
Statistical analyses
All statistical analyses were done using Stata 11 statistical package (STATA Corp, Timberlake Consultants). For MBL2 genotype analysis, normal homozygous MBL2 is denoted as A/A. The heterozygous variant MBL2 genotypes were grouped together as A/O heterozygous MBL2 as they all give low plasma MBL levels, and all homozygous and compound homozygotes were grouped together as O/O. Conformation to the Hardy–Weinberg equilibrium (HWE) was determined using the Chi Square test in the SHEsis online program (Li et al., 2009; Shi and He, 2005). The frequencies of the MBL2 alleles were obtained by direct gene counting. To determine prevalence of MBL deficiency, MBL2 genotypes were used as proxy for MBL levels, MBL2 A/A genotype was considered as normal, and those with A/O and O/O categorized as deficient MBL. Participants were classified into six MBL2 haplotype functional groups, which were combined into three groups for statistical analysis: haplotypes that give normal plasma MBL levels (YA/YA, YA/XA), intermediate levels (XA/XA, YA/YO), and deficient levels (XA/YO, YO/YO), and promoter region haplotypes (Garred et al., 2003; Vengen et al., 2012). Differences in proportions of MBL deficiency were determined using Chi Square or Fisher's exact tests. The differences in distribution frequency between HIV-1 infected and uninfected children, of the MBL2 genotypes, promoter region variants, and promoter haplotypes were determined by the Chi Square test or Fisher's exact tests. The effect of maternal MBL deficiency on HIV status of the children was determined in a logistic regression model after controlling for use of ART. The level of statistical significance used was p < 0.05.
Results
Demographic characteristics
The national PMTCT survey collected 8800 dried blood spots from mother baby pairs for HIV testing; among these, 880 mothers (10%) were HIV positive (Fig 1). Dried blood spots with an adequate sample were selected from 632 HIV-positive mothers and their children for inclusion in this substudy, that is, 1264 dried blood spots were selected for MBL analysis (Fig 1). The mothers' median age (interquartile range) was 30 (26–34) years old and the children were 12 (11–15) months old at the time of enrollment. Some of the HIV-positive mothers reported that they and their children were receiving ART (McCoy et al., 2015a, 2015b).
Frequency of MBL2 alleles and haplotypes of mothers and children
DNA was extracted from all 632 mother–child pair dried blood spots. Ten mother–baby pairs were excluded from analysis because the children had unconfirmed HIV status by the time of data analysis. We report here MBL2 analysis of 622 mother–baby pairs with complete information on MBL2 genotyping and HIV status. All 622 mothers were HIV-1 infected; there were 574 HIV-negative babies and 48 HIV-1-positive babies. The MBL2 exon 1 alleles A/A, A/C, C/C,
This analysis was done using direct gene counting, n = 622 mother–baby pairs, HIV-positive children n = 48, HIV-negative children n = 574.
HIV, human immunodeficiency virus; MBL, mannose binding lectin.
Relationship between mothers' and infants' MBL2 genotype, haplotypes, and HIV status
We determined the relationship between MBL2 genotype in the mothers and the HIV status of their children. We found no statistically significant difference in the distribution of the HIV infected and uninfected children between the MBL2 exon 1 genotypes of the mothers (p = 0.546) (Table 2). We also found no association between HIV infection and all the promoter genotypes detected and the three MBL2 functional classes (Table 2). We also found no statistically significant difference in the distribution of HIV infected and uninfected children in relation to the children's MBL2 genotypes, all promoter genotypes detected and the three MBL2 functional classes (Table 3). Participants were classified into six MBL2 haplotype functional groups, which were combined into three groups for statistical analysis: haplotypes that give normal plasma MBL levels (YA/YA, YA/XA), intermediate levels (XA/XA, YA/YO), and deficient levels (XA/YO, YO/YO) (Garred et al., 2003; Vengen et al., 2012).We found no statistically significant difference in distribution of HIV infected and uninfected children between these MBL2 functional groups of the mothers (Tables 2) and of the children (Table 3).
This analysis was done using the Chi Square and Fisher's exact tests, n = 622 mother–baby pairs, HIV-positive children n = 48, HIV-negative children n = 574.
This analysis was done using the Chi Square and Fisher's exact tests, n = 622 mother–baby pairs, HIV-positive children n = 48, HIV-negative children n = 574.
The effect of MBL deficiency on HIV status of children after controlling for ART
All the mothers were HIV positive, 72.3% (450 out of 622 mothers) were on ART, Nevirapine, AZT, and Nevirapine and AZT. Of the 622 perinatally exposed children, 71.5% (445 out of 622) of the children were on ART during the time of the survey using such drugs as Nevirapine only, Nevirapine and AZT syrup, and AZT only. The effect of MBL deficiency of the mothers on HIV status of the children was determined in a logistic regression model after controlling for use of ART by the mothers and was found not statistically significant (p = 0.624), OR 1.12 (95% CI), and the test of the overall model was also not statistically different (p = 0.421). The effect of MBL deficiency of the babies on their HIV status after controlling for use of ART by the children in a logistic model was not significant (p = 0.423), OR 1.18 (95% CI), and the test of the overall model was also not statistically different (p = 0.564).
The effect of MBL deficiency on HIV status of infants after controlling for type of ART used
Out of 622 mothers included in our analysis, 456 mothers were reported to be on maternal ARVs; additional information about what drugs they used is known for 306 mothers: 116 were on NVP only, 39 were on AZT only, 126 were on NVP and AZT, 7 were on HAART, and for 18 others the ARVs were not specified. The effect of MBL deficiency of the mothers on HIV status of the children was additionally determined in a logistic regression model after controlling for type of ART used by the mothers and was found not statistically significant (p = 0.633), OR 0.83, and the test of the overall model was also not statistically different (p = 0.877).
Out of 622 children included in this analysis, 449 children were reported to be on infant ARVs: 262 were on NVP & AZT syrup, 132 were on NVP only, 13 were on AZT only, and 42 children had unspecified ARVs. The effect of MBL deficiency of the babies on their HIV status was additionally determined in a logistic regression model after controlling for type of ART used by the children was not significant (p = 0.567), OR 1.18, and the test of the overall model was also not statistically different (p = 0.402).
Discussion
The main aim of our study was to determine prevalence and role of polymorphic MBL2 genotypes in vertical transmission among HIV-1-positive mothers and their HIV-1 infected or uninfected children enrolled in a national PMTCT survey. The prevalence of MBL deficiency as determined by presence of variant MBL2 genotypes A/O and O/O was high, at 34% among the mothers and 32% among the children, similar to other reports in children (Kuhn et al., 2006). We found no association between MBL deficiency of the mothers and of the children, HIV infection in the children and vertical transmission. We used MBL2 genotyping as a proxy to determine MBL deficiency in children, similar to reports by others (Amoroso et al., 1999; Arraes et al., 2006; Boniotto et al., 2000, 2003; Crovella et al., 2005; Dzwonek et al., 2006; Kuhn et al., 2006; Mangano et al., 2008; Singh et al., 2008; Zupin et al., 2016).
We detected currently known MBL2 Exon 1 coding and promoter region variants (Madsen et al., 1994, 1995, 1998) in our study population, in accordance with reports from other African populations (Garred et al., 1997; Lipscombe et al., 1992; Madsen et al., 1998; Madsen et al., 1995). We detected MBL2 variants B (codon 5 A>G) and C (codon 57, rs1800451 A>G) in our study population. The available evidence on association between HIV infection and MBL deficiency in children is conflicting (Eisen et al., 2008; Israels et al., 2012). Our results showed no association between mothers' MBL2 Exon 1 variants and HIV vertical transmission, in accordance with another published study on Zimbabwean children (Mhandire et al., 2014) and among Zambian mothers and their children (Zupin et al., 2016). In contrast to our findings, other studies reported an association between HIV vertical transmission and MBL deficiency, showing a statistically significant higher frequency of MBL2 Exon 1 variants among HIV-infected children (Amoroso et al., 1999; Arraes et al., 2006; Boniotto et al., 2000, 2003; Crovella et al., 2005; Dzwonek et al., 2006; Kuhn et al., 2006; Mangano et al., 2008; Singh et al., 2008).
Promoter region variants have been reported to significantly affect plasma/serum MBL concentration (Madsen et al., 1995). The H/L promoter alleles are associated with high (H/H), medium (H/L), and low (L/L) plasma/serum MBL expression; the MBL2 promoter region genotypes in combination with Exon 1 region genotypes result in MBL2 haplotypes with varying levels of serum/plasma MBL concentrations (Boldt and Petzl-Erler, 2002; Boldt et al., 2006, 2006, 2010; Madsen et al., 1995, 1998; Steffensen et al., 2000). In our study, the L/L allele had the highest frequency among both mothers and children, and H/H allele had the lowest frequency among both mothers and their infants, in accord with reports on African populations (Madsen et al., 1995). The H allele is found predominantly in white populations who also have very low L and X alleles (Madsen et al., 1995). Our results showed that the MBL2 variant haplotypes were not associated with HIV infection in the mothers, the children, or with MTCT. Mangano et al. (2008) also reported no difference in frequency of X/Y and H/L alleles between HIV infected and uninfected children (Mangano et al., 2008); others have also reported no association with MBL2 haplotypes (Mhandire et al., 2014; Zupin et al., 2016).
Studies done on African populations which reported results similar to ours include (Dzwonek et al., 2006; Kuhn et al., 2006; Mhandire et al., 2014; Zupin et al., 2016). We report here the frequency of A/O MBL2 variants among the children to be 35%, comparable to other reports of 24% (Mhandire et al., 2014) and 32.4% (Kuhn et al., 2006) on African children. Elsewhere frequencies of A/O MBL2 variants are reported as 41% among Hispanic children and 35% among white children (Singh et al., 2008). We found that a very low frequency of C/C homozygous MBL2 variant was 2% among HIV infected children, but others have reported frequencies of 20% among black children, 7% among Hispanics, and 5% among white children (Singh et al., 2008). We used a sample size of 622 mother and baby pairs with a 90% power to detect presence of any association between MBL deficiency, HIV infection in the children, and MTCT.
Most of the studies reporting effect of MBL deficiency on pediatric HIV infection were done during the pre-ART era, but the children in our study were on ART or were exposed to ART through treatment given to their mothers. We found MBL deficiency was not associated with HIV infection in a logistic regression model after controlling for ART in the children and their mothers. The only available study that has determined association between HIV paediatric infections and MBL deficiency in the context of ART (Dzwonek et al., 2006) found that despite the complications of performing this analysis in the era of HAART, MBL2 variants were associated with HIV infection and disease progression in children. The only study available in literature that showed association between MBL2 haplotypes XA/XA and HIV infection and faster disease progression was done in Argentina (Mangano et al., 2008).
In our study, distribution of the MBL2allele
We carried out similar research in an adult population in Zimbabwe and found that MBL deficiency neither played a role in HIV infection (Zinyama-Gutsire et al., 2015a,b) nor in disease progression and survival (Zinyama-Gutsire et al., 2015a).
A limitation in our study was that we did not have serum/plasma samples to determine MBL concentration. However, MBL2 exon 1 region and promoter region polymorphisms can be used as proxy to classify participants into normal plasma/serum MBL levels, intermediate, and deficient (Boldt and Petzl-Erler, 2002; Boldt et al., 2006, 2006, 2010; Madsen et al., 1994, 1995, 1998; Steffensen et al., 2000; Zupin et al., 2016).
Conclusion
In conclusion, our results showed a high prevalence of MBL2 genetic variant AC among the mothers and their children, in accordance with other studies of MBL2 genetic variation in African populations (Madsen et al., 1995; Mhandire et al., 2014; Zupin et al., 2016). This MBL2 variant form is associated with MBL deficiency; however, the high prevalence of MBL deficiency among the mothers and their children in our study was not associated with HIV-1 vertical transmission. It is important to explore the genetic makeup of the Zimbabwean population in view of the high prevalence of several viral, parasitic, and bacterial infections considering the conflicting evidence associating MBL deficiency and protection against some infections and increased susceptibility in others. It is our hope that more OMICS research in African countries will provide more knowledge on the hypothesis role of the MBL protein in HIV infection and vertical transmission, a current major health problem in African populations.
Footnotes
Acknowledgments
Serum State Institut (SSI) for providing accommodation, many thanks to SSI technical staff, namely Dennis Schmidt and Karina Liebmann Madsen, for expert technical assistance on MBL2 genotyping assays in Denmark, and the Director and the Executive Committee of the Medical Research Council of Zimbabwe (MRCZ) for permission and time to do this research project. Finally, special gratitude to all the women and children who participated in the national PMTCT survey in Zimbabwe in September 2012.
Authors' Contributions
All authors have met the ICMJE criteria for authorship and have read the article and approved the final version.
Financial Support
Funding was provided by UNFPA Zimbabwe and Soul City for the CESHAAR PMTCT National Survey. The Letten Research Foundation, University of Oslo, Norway/Zimbabwe Collaborative PMTCT BHAMC Program, special mention to the late Professor Letten F. Saugstad. Laboratory space and reagents for MBL2 genotyping were provided by Professor Michael Christiansen at Serum State Institut Copenhagen, Denmark.
An abstract of the results of this study was accepted and presented as a poster in Zimbabwe, Harare, at the Letten Foundation Research Symposium, 14–15 October 2015, funded by the Letten Research Foundation, University of Oslo Norway/Zimbabwe Collaborative Program. Another abstract on this study was accepted for presentation as a poster at the AIDS 2016: 21st International AIDS Conference, 18–22 July 2016, Durban, South Africa.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.