
Abstract
Abstract
Deciphering the role of cell-to-cell communication in acquisition of cancer traits such as metastasis is one of the key challenges of integrative biology and clinical oncology. In this context, extracellular vesicles (EVs) are important vectors in cell-to-cell communication and serve as conduits in the transfer of cellular constituents required for cell function and for the establishment of cellular phenotypes. In the case of malignancy, they have been shown to support the acquisition of common traits defined as constituting the hallmarks of cancer. Cellular biophysics has contributed to our understanding of some of these central traits with changes in tissue biomechanics reflective of cell state. Indeed, much is known about stiffness of the tissue scaffold in the context of cell invasion and migration. This article advances this knowledge frontier by showing for the first time that EVs are mediators of tissue biomechanical properties and, importantly, demonstrates a link between the acquisition of cancer multidrug resistance and increased tissue stiffness of the malignant mass. The methodology used in the study employed optical coherence elastography and atomic force microscopy on breast cancer cell monolayers and tumor spheroids. Specifically, we show here that the acquired changes in tissue stiffness can be attributed to the intracellular transfer of a protein complex comprising ezrin, radixin, moesin, CD44, and P-glycoprotein. This has important implications in facilitating mechano-transduced signaling cascades that regulate the acquisition of cancer traits, such as invasion and metastasis. Finally, this study also introduces novel targets and strategies for diagnostic and therapeutic innovation in oncology, with a view to prevention of metastatic spread and personalized medicine in cancer treatment.
Introduction
C
Extracellular vesicles (EVs) are small membrane vesicles derived from the ubiquitous cellular phenomenon of membrane budding (Bebawy et al., 2009; Gong et al., 2014; Hugel et al., 2005; Jaiswal et al., 2013; Lu et al., 2013, 2016). They carry surface antigens as well as cytoplasmic and nuclear constituents from their originating cell and mediate intercellular cross talk through the transfer of these constituents from donor cell to recipient cell (Gong et al., 2012; Hugel et al., 2005).
Previously, we have shown that a subtype of EVs derived from multidrug-resistant breast cancer cells, called microparticles (MPs), transfer P-gp to drug-sensitive cells and confer MDR in vitro and in vivo (Bebawy et al., 2009; Jaiswal et al., 2013) in less than 4 and 24 h, respectively. We also showed that the vesicular transfer of MDR is facilitated by the selective packaging of P-gp in MPs along with a protein complex comprising ezrin, radixin, moesin, and CD44 (Pokharel et al., 2014). Ezrin–radixin–moesin (ERM) proteins are a group of closely related proteins comprising three domains, including an N-terminal, central α-helical, and C-terminal tail domain (Tsukita et al., 1994; Turunen et al., 1994). ERM function as cytoskeletal linkers by binding directly to the C-terminal domain of actin filaments and the cytoplasmic NH2-terminal domain of integral membrane proteins, such as CD44, intercellular adhesion molecule-1 (ICAM-1), ICAM-2, and CD43 (Barreiro et al., 2002; Tsukita et al., 1994; Turunen et al., 1994; Yonemura et al., 1998). P-gp has been shown to bind to the N-terminal domain of ezrin as well as coregulating and coimmunoprecipitating with CD44 (Brambilla et al., 2012; Tsukita et al., 1994). In MDR cells, binding of CD44 with hyaluronic acid activates the expression of P-gp (Tsukita et al., 1994).
Similarly, the actin cytoskeleton interacts with P-gp for the endosomal trafficking of P-gp to the plasma membrane, demonstrating an essential role in its regulation and function (Fu and Roufogalis, 2007). We demonstrated that intercellular membrane insertion of P-gp in recipient cells was dependent on the presence of ezrin and moesin, whereas P-gp functionality is dependent on the presence of all ERM proteins in recipient cells (Pokharel et al., 2016). The acquisition of MDR was also shown by us to be associated with a conferred capacity for enhanced cell invasion and cell migration (Gong et al., 2014; Jaiswal et al., 2012).
Stiffness (which can be parameterized by the Young's modulus, E) is a biomechanical property of tissue that has been identified as a key factor in cellular function, including adherence, motility, and invasion (Cross et al., 2008; Schrader et al., 2011). Since cancer cells are known to be on average softer (lower Young's modulus) than noncancerous cells, stiffness can help in distinguishing nonmalignant from malignant tissue (Cross et al., 2008; Schrader et al., 2011). The reason for such biomechanical differences is yet to be fully understood; however, it is known that as healthy cells transform into cancerous states, their cytoskeletal structure changes from an organized to an irregular network, which is expected to alter their biomechanical properties (Iyer et al., 2009; Vignjevic and Montagnac, 2008).
We were interested in assessing the biomechanical properties of tissue with acquired MDR and enhanced invasive/metastatic capacity through the intercellular transfer of MDR-derived MPs. For this purpose, we developed a tumor spheroid model, which mimics the in vivo state of MDR intercellular transfer and provides for an organized tumor mass. We used the emerging technique of optical coherence elastography (OCE) to measure the biomechanical properties of the spheroids (Kennedy et al., 2014). OCE is a technology that uses the in vitro and in vivo imaging modality optical coherence tomography (OCT) to capture images of tissue elasticity (elastogram) undergoing deformation caused by external loading. OCE has several inherent advantages for measuring or imaging tissue biomechanics, including high mechanical sensitivity on the micron scale and cellular level over tissue volumes without use of exogenous agents (Kennedy et al., 2015a).
This study describes, for the first time, the role of EVs in regulating tissue biomechanical properties and demonstrates a link between acquisition of MDR and increased tissue stiffness of the malignant mass. We observed MDR breast cancer spheroids to have greater stiffness than drug-sensitive spheroids. We also detected an increase in stiffness in the drug-sensitive spheroids, following a 24-h coculture with MPs isolated from MDR cells. Additionally, we examined, using an atomic force microscope (AFM), the role of a protein complex, which included P-gp, CD44, and ERM, in regulating the biomechanical properties of recipient cells. We observed a significant decrease in the stiffness of drug-resistant breast cancer cells upon silencing P-gp and CD44 and a significant increase in stiffness following silencing of the entire ERM protein complex. Similar to the spheroid model, the transfer of MPs resulted in an increase in the stiffness of drug-sensitive breast cancer cells when grown as monolayers.
Materials and Methods
Cell culture
The drug-sensitive human breast adenocarcinoma cell line, MCF-7, the drug-resistant cell line, MCF-7/Dx (referred to as Dx for simplicity), and the drug-resistant human acute lymphoblastic leukemia cell line, VLB100, were used in these studies. These cell lines have been validated as appropriate models for the study of P-gp-mediated MDR in vitro and in vivo (Bebawy et al., 2009; Jaiswal et al., 2013). All cells were cultured in RPMI 1640 medium (Sigma-Aldrich), supplemented with 10% (v/v) heat-inactivated fetal bovine serum (Life Technologies), and maintained in the absence of antibiotics at 37°C and 5% CO2.
Harvesting MPs
We have previously described the isolation of MPs and characterization of their morphology, size, and resistance protein expression (Bebawy et al., 2009; Gong et al., 2013; Lu et al., 2013). The protocols for in vitro gene silencing and immunohistochemistry have been also previously described by us in Pokharel et al. (2016) and Jaiswal et al. (2013), respectively.
Protocol for 3D spheroid cultures
Individual wells in 96-well plates were coated with 50 μL of heat-sterilized 1.5% agarose (in RPMI). Plates were exposed to ultraviolet light for 45 min; 4 × 103 cells in 200 μL of complete media were added to each well. Cells were incubated at 37°C with 5% CO2 for 4–6 days to allow for cell aggregation and spheroid formation. MP transfer experiments were performed on day 6, following which the spheroids were fixed and embedded in paraffin as described previously (Jaiswal et al., 2013).
MP transfer experiment
Coculture experiments on drug-sensitive MCF-7 cells and their spheroids were performed over 24 h using 180 μg of MPs isolated from drug-resistant breast cancer cells (Dx-MPs) or from drug-resistant leukemic cells (VLB100-MPs) at 37°C and 5% CO2. Unbound MPs were removed by washing the spheroids three times with PBS at ambient temperature. The cells and spheroids were fixed in 4% formaldehyde for 30 min before analysis to allow for preservation of cellular height and structural integrity (Chang et al., 1993; Francis et al., 2010; Sinniah et al., 2002).
Atomic force microscopy
A JPK Nanowizard SENSE AFM (JPK Instruments) was used for both imaging and mechanical measurements. All measurements were performed in liquid using contact mode with sharpened silicon nitride cantilevers (L:225 μm, W:46 μm, T:1.0 μm, SPM Probe Model:SHOCON; AppNano) with spring constant of 0.03 N/m and tip radius of <10 nm. Initially, the cell image was obtained and the cantilever was precisely positioned on the top/center of cells to avoid the cell edge (marked spots in Fig. 1) over a 3 × 3 μm area. Force–displacement curves were recorded at 1 Hz to determine the Young's modulus (E), a quantitative indicator of cell stiffness (Ophir et al., 1991). The data files were analyzed using JPK data processing (version spm-5.0.63) and the measured stiffness was expressed as averaged E value.
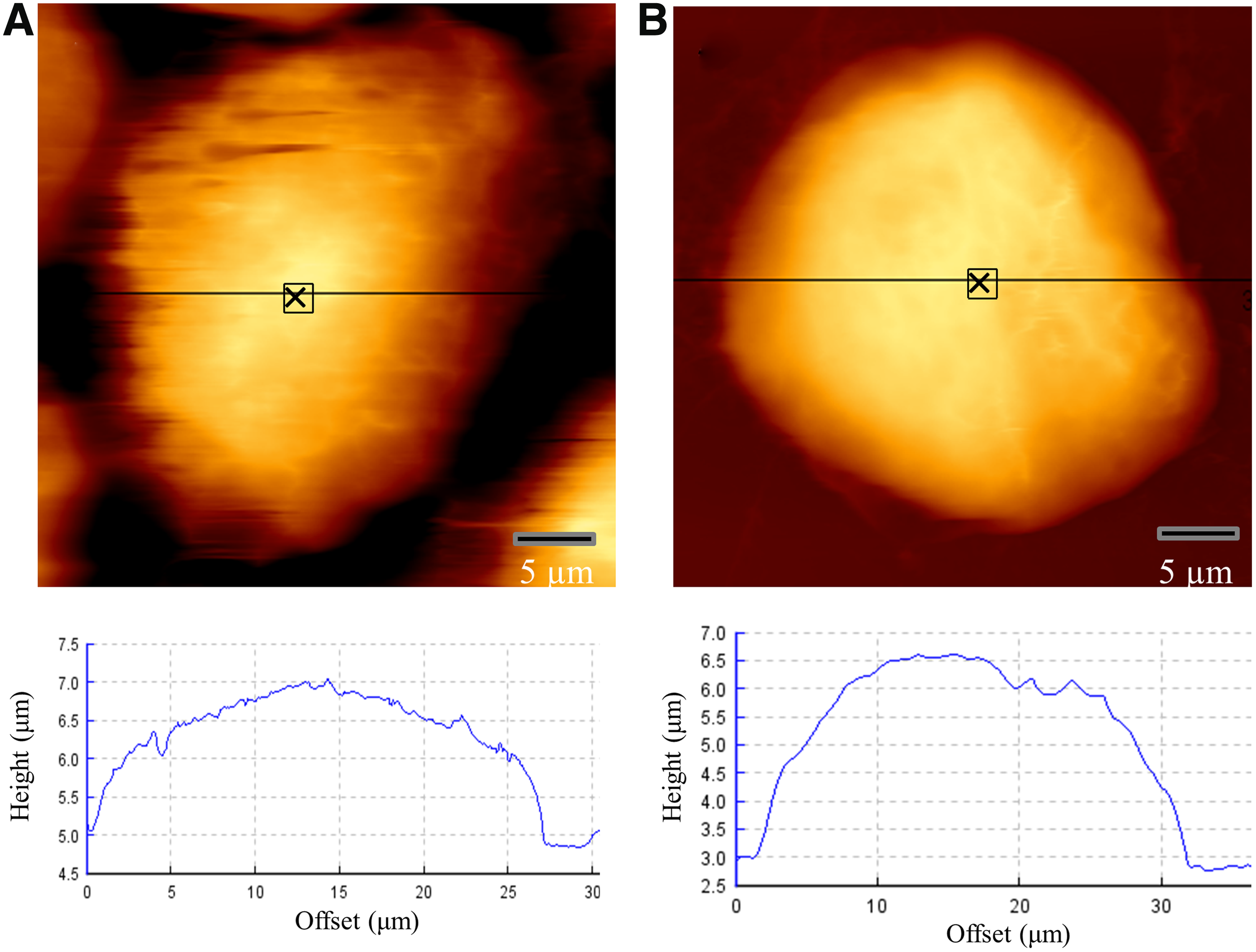
Representative AFM topographical height measurement image of
Optical coherence elastography
OCE is an emerging e method in tissue biomechanics. OCE utilizes the imaging modality, OCT (Kennedy et al., 2014). With the inherent advantages of high-resolution real-time imaging and millimeter-scale penetration depth in tissue, OCE techniques are suited to measuring and imaging biomechanical properties on the micro- to millimeter scale at the tissue and cellular levels (Curatolo et al., 2016; Kennedy et al., 2015a; Mulligan et al., 2016; Wijesinghe et al., 2015). The version of OCE employed here has been previously described in detail (Kennedy et al., 2009, 2015b). Briefly, tissue stiffness was extracted by step-wise application and release of compression to the sample and measuring the resultant deformation using a fiber-based spectral-domain OCT system. The OCT system employs a superluminescent diode source operating at a central wavelength of 835 nm and with a bandwidth of 50 nm, illuminating the sample with 6.5 mW of power. The resolution of OCT structural images was 11 μm laterally and 7.8 μm (in air) axially (depth). The stiffness spatial resolution of the OCE system was 20 μm and 50 μm in the transverse and axial (depth) directions, respectively. The OCE images presented here were sectioned 20 μm below the top surface of the spheroid. OCE images were overlaid on grayscale OCT structural images, primarily to mask out regions external to the spheroid mass.
Statistical analyses
The measurements of Young's modulus are expressed as mean ± SD of two to three independent experiments. The average was obtained from measurements within the full volume of the spheroids, excluding a 50-μm-thick rim around the edge to minimize edge effects. The data were analyzed using one-way analysis of variance (ANOVA) and p-values of <0.05 were accepted as being statistically significant. All statistical analyses were performed using GraphPad Prism version 5.0 for Windows Software (GraphPad Software).
Results
MP transfer using in vitro tumor spheroids mimics MP transfer in mouse xenograft models in vivo
Previously, we showed that MPs isolated from MDR breast cancer cells (Dx-MPs) transfer P-gp in vitro and in vivo (Bebawy et al., 2009; Gong et al., 2014; Jaiswal et al., 2013; Lu et al., 2013). MP transfer of P-gp in vivo resulted in the transfer and localization of P-gp within the core of the solid tumor mass within 24 h of MP exposure (Jaiswal et al., 2013). The protein was stably expressed in the absence of drug or subsequent MP exposure for at least 2 weeks following a single subcutaneous injection (Jaiswal et al., 2013). In vivo xenograft models are costly, labor intensive, require specialist facilities/infrastructure, and are associated with ethical issues and barriers. In contrast, conventional 2D in vitro cell models are cost-effective experimental models, less labor intensive, and are of high capacity (Osato et al., 2003). However, 2D cell culture models lack the characteristics of human tumor tissues, including the morphology, metabolic requirements, and tumor microenvironment. Therefore, we sought to establish a tumor spheroid model to mimic the in vivo state with respect to MP-mediated transfer of P-gp and remove the need for animal experimentation required to define the MDR pathway.
We observed that MCF-7 spheroids were highly susceptible to disassociation upon handling (Fig. 2A). In comparison, the spheroids established from Dx cells were compact with a distinctly spherical morphology (Fig. 2B). We cocultured MCF-7 spheroids with Dx-MPs for 24 h and performed immunohistochemistry to look for the presence of P-gp. As expected, we detected no P-gp expression in the MCF-7 spheroid control (Fig. 3A, B); however, consistent with our previous findings in vivo, we observed the efficient transfer of P-gp to the MCF-7 tumor spheroid core 24 h after MP exposure (Fig. 3C, D).

OCE image of drug-sensitive and MDR tumor spheroids:
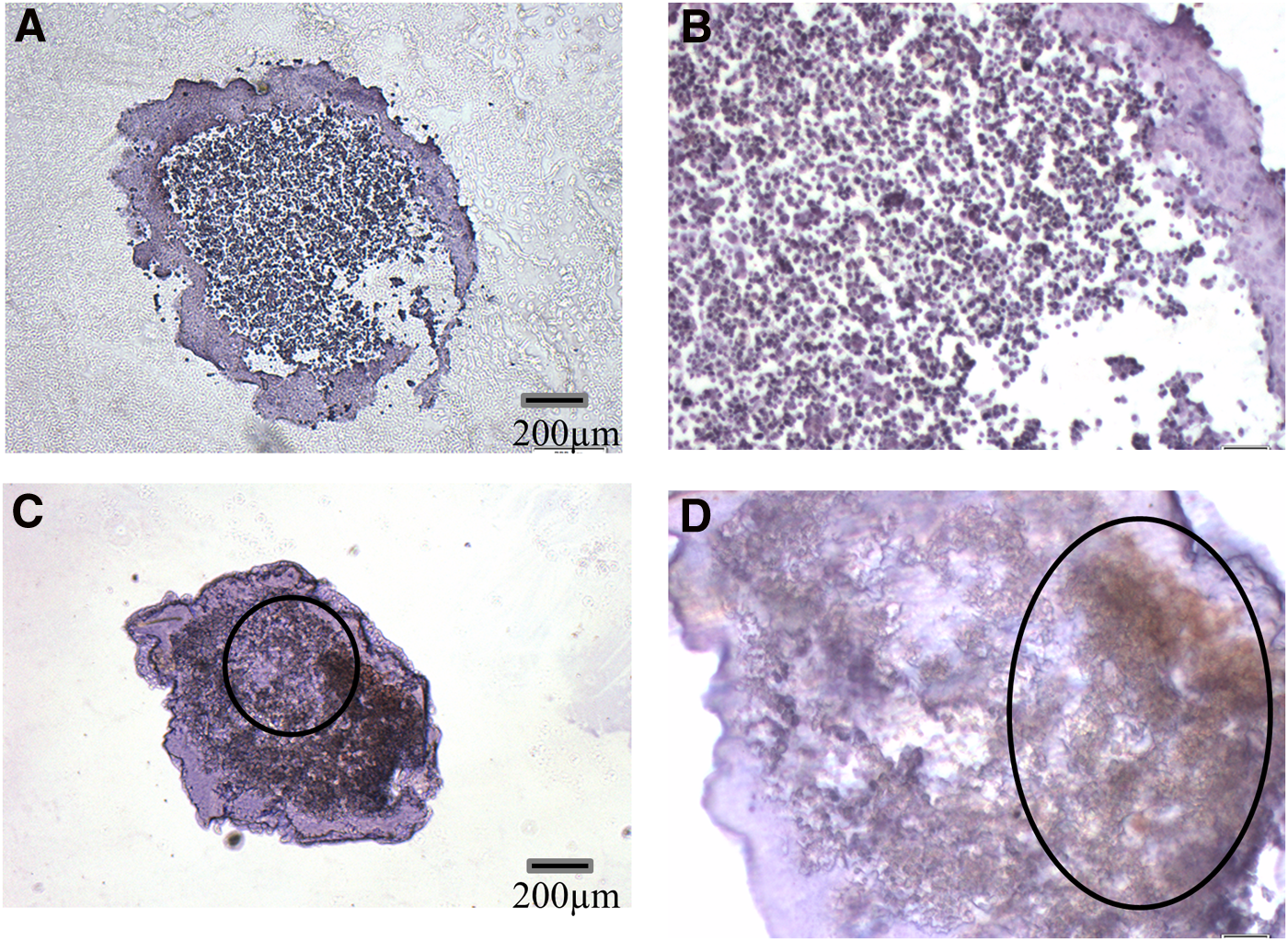
P-gp localized in tumor spheroid core following MP transfer: MCF-7 tumor spheroids were cocultured for 24 h with Dx-MPs. The spheroid was resected and fixed and sections subjected to immunohistochemical staining with antibodies specific to P-gp. Sections were examined at
Biomechanical properties of drug-resistant and drug-sensitive spheroids
We measured the stiffness of Dx and MCF-7 spheroids using OCE (Fig. 4A). We observed the Dx spheroids to have a 2.2 ± 0.15-fold greater Young's modulus relative to MCF-7 spheroids (p < 0.01) (Fig. 4A). Additionally, we used AFM to compare the stiffness of cells grown as a monolayer and observed similar results (Fig. 4B). Dx cells display a significant 2.3 ± 0.08-fold greater stiffness (higher Young's modulus) relative to MCF-7 cells (p < 0.05) (Fig. 4B).
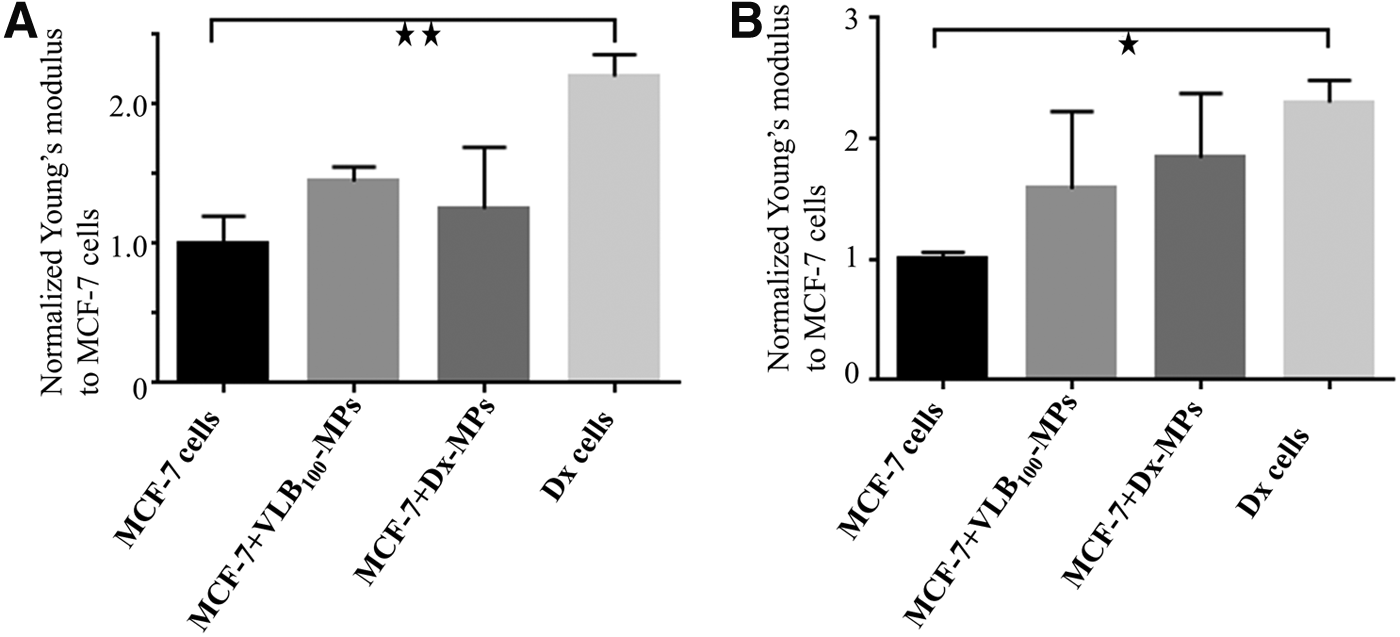
Comparative stiffness of cell monolayers in the presence and absence of MDR MP exposure:
MPs derived from MDR cancer cells increase the stiffness of drug-sensitive spheroids and monolayers
Previously, we have shown that MPs derived from MDR cancer cells transfer their protein cargo and confer a drug-resistant phenotype in less than 4 h (Bebawy et al., 2009; Jaiswal et al., 2013). We also showed that acquisition of MDR resulted in an enhanced capacity for cell invasion and migration (Gong et al., 2014; Jaiswal et al., 2012). In this study, we sought to examine whether the acquisition of these traits was also associated with a change in the biomechanical properties of the drug-sensitive recipient cells. Dx-MPs and MPs derived from MDR leukemic cells (VLB100-MPs) were used for this experiment. Both MPs represent EVs derived from MDR cancers of solid and hematological origin and allow for the study of the effect of transferred characteristics irrespective of cell origin. MPs were cocultured for 24 h with drug-sensitive MCF-7 cell monolayers and spheroids and stiffness analyzed using AFM and OCE, respectively.
We observed an increase in stiffness of the MCF-7 monolayers and spheroids by 1.58 ± 0.64 and 1.45 ± 0.13-fold, respectively, after coculturing with VLB100-MPs (Fig. 4A, B). Similarly, the stiffness of MCF-7 cells increased by a factor of 1.84 ± 0.53 and 1.25 ± 0.44-fold in monolayers and spheroids, respectively, after coculturing with Dx-MPs (Fig. 4). The differences in stiffness of drug-sensitive and MDR cells are not associated with morphological differences at the individual cell level. There was no significant difference in height (Dx cell: 7.88 ± 1.89 μm, MCF-7 cell: 8.13 ± 1.86 μm) or area (Dx cell: 29.23 ± 3.34 μm, MCF-7 cell: 29.84 ± 3.82 μm) between MCF-7 and Dx cells as visualized using AFM (Fig. 1). We did observe that Dx cells grow in loose disaggregated clusters as a monolayer, but they organize into a strong compact mass as spheroids (Fig. 5). On the contrary, MCF-7 cells grow in dense multicellular colonies as a monolayer, but form weak spheroids (Fig. 5).

MCF-7 and DX cells and clusters as visualized using AFM:
Effect of silencing of P-gp, ERM, and CD44 on the stiffness of MDR breast cancer cells
We have previously shown that the acquisition of functional MDR involves a protein complex, including P-gp, CD44, and ERM proteins (Pokharel et al., 2014). We have shown that radixin and CD44 play a significant role in P-gp functionality, whereas the vesicular transfer of P-gp requires the presence of all members of the ERM family in recipient cells (Pokharel et al., 2016). We sought to examine the role of these proteins in regulating tissue stiffness. We silenced P-gp, CD44, and ERM individually in Dx cells and examined the changes in stiffness as measured using AFM. We observed a significant 1.87 ± 0.13 (p < 0.001) and 1.62 ± 0.03 (p < 0.001)-fold decrease in the stiffness of Dx monolayers after silencing P-gp and CD44, respectively. Silencing of the ERM protein complex resulted in a 1.15 ± 0.11-fold increase in cell stiffness. Upon silencing P-gp and CD44, or ERM and CD44, we observe a 1.32 ± 0.05 (p < 0.01) and 1.21 ± 0.10-fold increase in stiffness of Dx cell monolayers, respectively (Fig. 6).
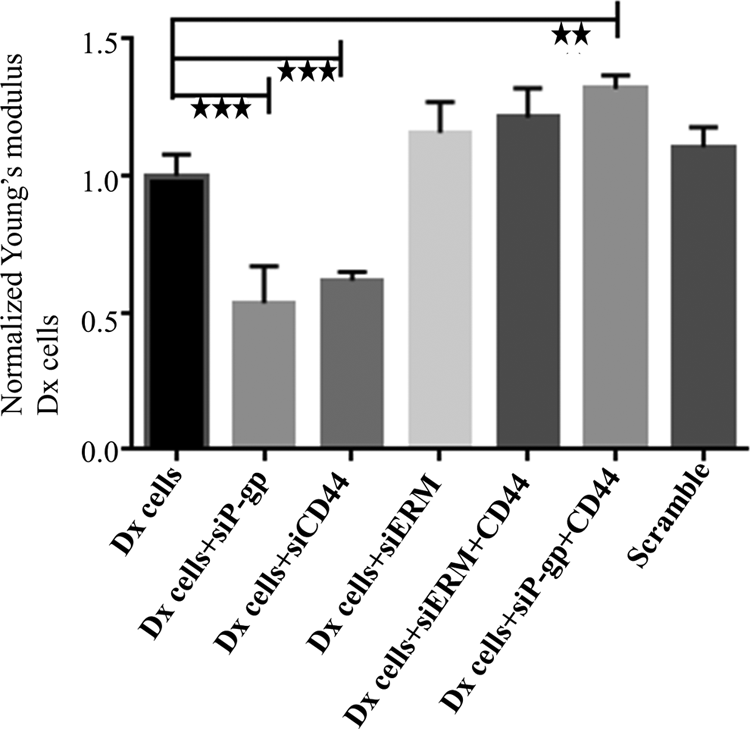
Effect of silencing of P-gp, ERM, and CD44 proteins on stiffness in Dx cells: Dx cells were silenced using siRNAs targeting P-gp (siP-gp), CD44 (siCD44), ezrin–radixin–moesin (siERM), and the combination of siP-gp+siCD44 and siERM+siCD44 over 3 days. Stiffness was measured using AFM. Data were normalized to control Dx cells. Data represent mean ± SD (n = 3). **p < 0.01, ***p < 0.001.
Discussion
Cancer cell migration and invasion are complex events that are influenced by biomechanical properties of the tissue (Cross et al., 2008; Swaminathan et al., 2011). Several studies have reported on the biomechanical disparity between normal and cancer cells (Cross et al., 2008). However, little is known about the distinct biomechanical properties of malignant tissue in the context of its drug-responsive state. Likewise, the role of EVs in regulating tissue biomechanical properties has previously been unexamined.
In this study, we assessed the biomechanical properties (stiffness) of drug-sensitive MCF-7 spheroids and their MDR counterparts; Dx spheroids as well as MCF-7 spheroids that had acquired the presence of the MDR protein, P-gp, following intercellular transfer by MPs. Using OCE, we show that Dx spheroids are stiffer than MCF-7 spheroids (Fig. 4). A similar result was reported in cisplatin-resistant human ovarian cancer cells, where the resistant cells had significantly greater cell stiffness relative to drug-sensitive cells (Sharma et al., 2012). Interestingly ovarian cancer cells with a greater stiffness had lower migration and invasion properties (Xu et al., 2012). However, our previous studies combined with the current studies show that drug-resistant breast cancer cells were more invasive, migratory, and stiffer than drug-sensitive cells (Gong et al., 2014). The basis for these observed differences is unknown and may relate to the tissue examined and the nature of drug exposure. This current study demonstrates that tissue stiffness has the potential to be used as a clinical prognostic tool as surrogate marker of MDR status.
An interesting finding arising from this work is the discovery that MPs confer biomechanical alterations in recipient cell populations. We observed an increase in stiffness of MCF-7 spheroids after coculture with Dx-MPs and VLB100-MPs (Fig. 4). Previously, we have shown that MPs are powerful vectors in the intercellular transfer of functional resistance proteins and identified the selective packaging of ERM and CD44 among other proteins in Dx-MPs (Pokharel et al., 2014). We showed that these protein constituents are required for the tissue-selective transfer of P-gp and in the insertion and functionality of P-gp in recipient cells by MPs (Jaiswal et al., 2013; Pokharel et al., 2014, 2016).
In this study, we used siRNA to silence P-gp, CD44, and ERM in Dx cells and assessed the effect of this on the stiffness of cell monolayers using AFM. We observed a significant decrease in the stiffness of Dx cells after silencing either P-gp or CD44, supporting their involvement in regulating tissue biomechanical properties (Fig. 6). Interestingly, the silencing of ERM together with CD44 as well as P-gp together with CD44 resulted in an increase in cell stiffness (Fig. 6). ERM, CD44, and actin are found to be colocalized with P-gp in MDR cells and directly interact to affect its membrane localization and drug efflux function (Bourguignon et al., 2008; Luciani et al., 2002; Zhang et al., 2013). P-gp and CD44 are membrane-localized proteins, whereas ERM proteins cross-link these membrane proteins with the actin cytoskeleton (Jensen et al., 2009; Tsukita et al., 1994; Yonemura et al., 1998).
The basis for the increase in stiffness observed under these is currently unknown and requires investigation in the role of these proteins at the level of the membrane ultrastructure and cytoskeleton. Indeed, disruption of actin cytoskeleton and Rho GTPase has been shown to impact on tissue stiffness (Sharma et al., 2012).
Conclusions and Ramifications
We describe, for the first time to the best of our knowledge, a role for MPs in regulating the biomechanical properties of malignant tissue. We show that MPs shed from MDR cells increase the stiffness of recipient cells. We confirm the role of P-gp, and CD44 in particular, in affecting the Young's modulus of MDR cancer cells, suggesting that their transfer by MPs plays a role in the observed increase in the stiffness of recipient cells. These observations have broader clinical and cell biology ramifications and introduce, for the future, novel targets and strategies for diagnostic and therapeutic innovation in oncology, with a view to prevention of metastatic spread and personalized medicine in cancer treatment.
Footnotes
Acknowledgments
The authors thank Dr. Ritu Jaiswal for reading the manuscript and Dr. Stuart Hodgetts for providing his laboratory for experiments at the University of Western Australia.
Authors' Contributions
D.P. performed the experiments, analyzed the data, and prepared the manuscript. P.W. assisted with OCE, V.O. assisted with immunostaining, and J.F.L., D.D.S., B.F.K., and V.P.W. provided the technical support, conceptual advice, and revised the manuscript. M.B. designed and supervised the research project, provided conceptual advice, and revised the manuscript. All authors approved the final version of the manuscript for publication.
Author Disclosure Statement
No competing financial interests exist.