
Abstract
Abstract
Medicinal plants are part of the healthcare systems worldwide, especially in low- and middle-income countries. African lettuce (Launaea taraxacifolia) is cultivated extensively in Africa, from Senegal in the west to Ethiopia and Tanzania in the east, and in Southern Africa. Potential anticancer effects of L. taraxacifolia have been suggested, but little is known about putative molecular mechanisms or potential for herb–drug interactions through inhibition or induction of drug-metabolizing enzymes. We investigated the effects of crude aqueous extracts of L. taraxacifolia on growth kinetics and cell cycle progression of the WHC01 esophageal cancer cells. Antiproliferative and apoptotic effects were evaluated using the MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] assay and flow cytometry, while examining, in parallel, the genes regulating apoptosis and cell cycle in this cell culture model. In addition, we tested the inhibitory and enzyme kinetic effects of the aqueous L. taraxacifolia using recombinant human CYP450 isozyme model systems (CYP1A2, CYP2C9, and CYP2C19). L. taraxacifolia exhibited a significant growth inhibitory effect on the WHC01 cancer cells. Most cell cycle genes were downregulated. Cell cycle analysis showed a G0-G1 cell cycle arrest in WHC01 cells in the presence of L. taraxacifolia extract, accompanied by morphological changes. L. taraxacifolia extract treatment resulted in downregulation of expression levels of CYP1A2 (p < 0.0005) and CYP2C19 (p < 0.003) by 50–70%. L. taraxacifolia extract caused reversible and time-dependent inhibition of the recombinant CYP1A2, CYP2C9, and CYP2C19. This study provides new insights on possible anticancer effects of L. taraxacifolia, a widely used medicinal plant in parts of Africa and across the world especially by patients with cancer. Further mechanistic studies expanding on these observations would be timely and contribute to the field of global precision medicine that requires solid understanding of drug and herb molecular mechanisms of action and drug–herb interaction potentials, given the worldwide use of medicinal plants.
Introduction
C
Among cancers impacting global health, esophageal cancer is the eighth world-wide with an estimated 3.2% increase in new cases as on 2012 and the sixth most common cause of death with a projected mortality of 4.9%, presenting a serious health burden (Ferlay et al., 2015). There is a higher probability of 1 in 33 chances for a black South African male to develop esophageal cancer (Dandara et al., 2016). Major risk factors that are associated with cancers, including carcinoma of the esophagus, include tobacco smoking, alcohol consumption, nutritional deviancies, and environmental exposures (Danaei et al., 2005). Cancer control plans are needed to improve prevention and care, especially in developing countries, which are presenting with a growing burden.
The use of herbal medicines for the management of diseases, including cancer, in addition to allopathic medication, is a common phenomenon in Africa and across the world (Thomford et al., 2015). Data available show that patients hardly inform their physicians about the dual use of medicinal plant and prescribed drugs (Djuv et al., 2013). Launaea taraxacifolia of the family Asteraceae is a medicinal herbal plant grown and commercialized in different African countries such as Benin, Ghana, Ethiopia, Senegal, Sudan, and South Africa (Adejuwon et al., 2015; Ngozi et al., 2014). It is believed (through indigenous knowledge) to have lactogenic, aphrodisiac, antibiotic, antimalarial, and anticancer properties and blood pressure regulating and hemorrhoid treatment capacity (Dansi et al., 2008).
L. taraxacifolia contains vitamins, minerals, proteins, essential fats, and high amount of antioxidants (Adinortey et al., 2012). Medicinal and natural herbal products are thought to present with potentially high antioxidant activity and therefore often administered alongside chemotherapeutic agents to provide better protection against chemotherapeutic toxic side effects (Adejuwon et al., 2015; Nasr and Saleh, 2014). Since chemotherapy targets cancer cells and these cells express genes such as CYP450, involved in the metabolism of many drugs, the course of herbal medicine with chemotherapeutic agents have pharmacogenomic implications, especially efficacy and toxicity.
It is therefore important to characterize the effects of medicinal plants because of their unregulated use in healthcare systems across the world. L. taraxacifolia (African lettuce) is cultivated extensively in Africa from Senegal in the west to Ethiopia and Tanzania in the east and across to southern Africa. Potential anticancer effects of L. taraxacifolia have been suggested, but little is known on its putative molecular mechanisms. In addition, the effects of L. taraxacifolia enzymes that facilitate drug metabolism are not known. Drug-metabolizing enzymes, which include the cytochrome P450 family and drug transporters, play key roles in determining the pharmacokinetics and overall disposition of chemotherapeutic agents in the body.
In this study, we set out to investigate the effects of crude aqueous extracts of L. taraxacifolia on the growth kinetics and cell cycle progression of the WHC01 esophageal cancer cells using the MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] assay and flow cytometry. The second objective involved evaluating the inhibitory and enzyme kinetic effects of the aqueous L. taraxacifolia using a recombinant human CYP450 isozyme model system for CYP1A2, CYP2C9, and CYP2C19 as part of pharmacogenomic profiling of herbal medicinal plants.
Materials and Methods
Ethical approval
Ethical approval was obtained from the University of Cape Town Human Ethics committee (No. HREC REF: 826/2014).
Medicinal herbs
L. taraxacifolia, (UCC/BS/690) used in the study was obtained and authenticated by botanists from the University of Cape Coast (Cape Coast, Ghana). Voucher specimen is deposited at the University of Cape Coast herbarium. Plant material was dried at room temperature and grounded. The resulting powdery plant sample was then extracted using water to mimic the indigenous mode of extraction.
Cell culture
WHC01 esophageal cancer line was derived from a biopsy of primary esophageal squamous cell carcinoma of South African origin (Veale and Thornley, 1989). The WHC01 cells were kindly provided by the International Centre for Genetic Engineering and Biotechnology (ICGEB), Cape Town Component, University of Cape Town. The WHC01 cells were maintained as an attached monolayer cultured in Dulbecco's modified Eagle medium (DMEM; Thermo Fischer Scientific, Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS; Thermo Fischer Scientific) and 100 U/mL penicillin, and 100 μg/mL streptomycin (Life Technologies, Grand Island, NY, USA) in a 10 cm dish. The cells were then incubated at 37°C in a humidified atmosphere of 5% CO2. Confluent monolayer cells were passaged by treating them with 0.05% Trypsin-EDTA solution.
Evaluation of cytotoxicity using MTT assay
Cytotoxicity of L. taraxacifolia plant extract was evaluated using the standard MTT (Sigma-Aldrich, Saint Louis, MO, USA) cellular viability assay. The concentration for 50% of maximal inhibition (IC50) value, which is the drug concentration at which 50% of cells are viable, was determined according to the manufacturer's protocol (Sigma-Aldrich, St. Louis, MO). Briefly, cells were seeded in 96-well plates at a concentration of 3 × 103 cells/well and incubated overnight to attach. Media containing different concentrations (0.2–4.0 mg/mL) of L. taraxacifolia extract was added to each well and incubated for 48 h. Then 10 μL of 5 mg/mL MTT was added and incubated with the cells for 4 h. This was followed by addition of 100 μL 10% sodium dodecyl sulfate (SDS) and 0.01 M HCl (Merck, Darmstadt, Germany) to solubilize the formazan crystals. Absorbance was read on a multiscan plate reader at 595 nm. IC50 was calculated from the sigmoidal dose–response variable slope curve fitting using GraphPad prism 5.0® software (GraphPad Software, Inc., San Diego, CA, USA).
Investigating the antiproliferative effects of L. taraxacifolia using cell proliferation/counting assay
The antiproliferative effects of aqueous extracts of L. taraxacifolia on the WHC01 cells were evaluated using cell counting employing the trypan blue exclusion method. Cells were seeded in six-well plates at a concentration of 5 × 103 cells/well and incubated overnight. After incubation, media containing serial dilutions (1:3) of each herbal extract ranging from 0.2 to 2.0 mg/mL were added (1000 μL/well) in triplicates. Each plate included cells cultured in plain medium with no herbal extract treatment (experimental control). Cells were cultured for 48 h after which media were aspirated and cells were washed twice with phosphate-buffered saline (PBS, 1 × ). Cells were trypsinized and suspended in 1 mL medium. Ten microliters trypan blue and 10 μL of suspended cells were mixed and counting performed in a Countess automated cell counter (Invitrogen, Carlsbad, CA, USA). Results of cell number and viability were analyzed using GraphPad prism 5.0 software (GraphPad Software, Inc.).
Cell treatment
WHC01 cells were maintained as an attached monolayer in DMEM supplemented with 10% FBS and 100 U/mL penicillin, and 100 μg/mL streptomycin for 7 h. The culture medium was changed to serum-free medium and cells maintained for 16 h. WHC01 cells were then washed with PBS and fresh media added. Fresh media contained aqueous extract of L. taraxacifolia and incubation was continued for a further 48 h. Untreated cells served as control with acetaminophen (ACET) and 5-flourouracil (5-FU) serving as a positive control. After incubation, cells were washed twice with PBS and used in various analyses as indicated below.
RNA extraction and Real-Time quantitative Polymerase Chain Reaction (RT-qPCR)
Total RNA was isolated using Trizol reagent (Life Technologies, Grand Island, NY) according to the protocol by Chomczynski and Sacchi (2006). Purity of the isolated RNA was determined using Nanodrop spectrophotometer by measuring the optical density of the sample at 260 and 280 nm. Complementary DNA (cDNA) was generated from 2 μg of total RNA using the Impromp II reverse transcriptase (Promega, Madison, WI, USA) according to the manufacturer's protocol. Real-time quantitative polymerase chain reactions were performed and monitored using the Light cycler 480 II (Roche, Indianapolis, IN, USA) for cell cycle genes and Bio-Rad CFX-96 thermal cycler (Bio-Rad, San Francisco, CA, USA) for CYP450 genes.
The cDNA samples in a total volume of 20 μL in triplicates samples were analyzed using primers listed in Table 1 under the following conditions: initial denaturation at 94°C for 5 min followed by 35 cycles of 94°C for 20 sec, 55°C for 20 sec, and 72°C for 20 sec for the cell cycle genes. For CYP450, the following conditions were used: initial denaturation at 95°C for 1 min, followed by 40 cycles of 95°C for 10 sec, temperature gradient range of 53–62°C for 20 sec with melt curve analysis for 10 sec for CYP450 genes. The cycle threshold (Ct) values for each sample were obtained and the relative gene expression levels were computed using untreated control cells with the data normalized to a reference gene GAPDH (glyceraldehyde 3-phosphate dehydrogenase). Changes in expression were illustrated as fold increase or decrease where significance was set at p < 0.05.
BCL-2, B-cell lymphoma 2; BCL-XL, B-cell lymphoma-extra large; CYP1A2, cytochrome P450 1A2; CYP2C19, cytochrome P450 2C19; CYP2C9, cytochrome P450 2C9; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; MMP-9, matrix metallopeptidase 9; PCNA, proliferating cell nuclear antigen.
Western blot analysis
Control cells and those treated with extract of L. taraxacifolia were washed with ice-cold PBS, lysed in RIPA buffer (10 mM Tris-HCl pH 7.6, 10 mM NaCl, 3 mM MgCl2, and 1% [v/v] Nonidet P-40), and protease inhibitors added. The protein concentration was determined using the Pierce™ BCA protein assay kits (Thermo Fischer Scientific) according to the manufacturer's protocol using bovine serum albumin as standard. Total protein lysate (50 μg) was separated on 10% polyacrylamide/SDS gels under reducing conditions. The proteins were then transferred from the gel to a nitrocellulose membrane and blocked in 5% fat-free milk in Tris-buffered saline containing tween-20 (TBST). The membrane was incubated overnight at 4°C with primary antibodies (Santa Cruz Biotechnology, Santa Cruz, CA, USA) as indicated in Table 2. The membranes were washed with TBST and reincubated with specific horseradish peroxidase-conjugated secondary antibodies also from Santa Cruz Biotechnology as shown in Table 2 for 1 h. After two washes, detection was done using Lumiglo substrate (KPL, Gaithersburg, MD, USA).
PTEN, phosphatase and tension homolog.
Flow cytometry/cell cycle analysis
Cells (1 × 106) were seeded in 10 cm dishes and treated with aqueous extracts of L. taraxacifolia and 5-FU. Untreated cells served as controls. The cells were washed twice with cold PBS. Cells were detached and processed for flow cytometry analysis. The cells were resuspended in PBS and fixed in ice-cold 70% ethanol and kept at 4°C for 60 min. The cells were washed with PBS and stained with 200 μL of propidium iodide stock solution (50 μg/mL propidium iodide, 3.8 mM sodium triphosphate in PBS) supplemented with 50 μL RNase A (10 μg/mL) for 3 h at 4°C and then analyzed with a FACScan cell sorter (Becton Dickinson, Franklin Lakes, NJ, USA). Ten thousand cells were collected and the cell cycle profiles were calculated using the Cellquest software (Becton Dickinson, San Jose, CA).
Reversible time-dependent inhibition and kinetics determination
Determination of reversible and time-dependent inhibition (TDI) of extract of L. taraxacifolia was assessed using Vivid® CYP450 Screening Kits (Life Technologies, Grand Island, NY) according to the manufacturer's protocol. The kit is designed to assess the metabolic activity and inhibition of recombinant human CYP450 isozymes involved in drug metabolism by the use of 7-benzyl-oxymethyloxy-3-cyanocoumarin (BOMCC) and 7-ethoxy-methloxy-3-cyanocoumarin (EOMCC) as probe substrates. The protocol for reversible and time-dependent inhibitory profile was as outlined by Thomford et al. (2016). The kinetic profile of the L. taraxacifolia extracts on CYP1A2, CYP2C9, and CYP2C19 enzyme activity was performed according to a published protocol (Krippendorff et al., 2009). The conditions and concentration of the enzymes are indicated in Table 3.
BOMCC, 7-benzyl-oxymethyloxy-3-cyanocoumarin; Em, emission wavelength; EOMCC, 7-ethoxy-methloxy-3-cyanocoumarin; Ex, excitation wavelength.
Briefly, L. taraxacifolia extract with a starting concentration of 100 μg/mL was serially diluted six consecutive times in a black Costar (Corning, Cambridge, MA, USA) 96-well plate and preincubated with a master mix A (regeneration system, NADPH [nicotinamide adenine dinucleotide phosphate], reaction buffer) for 10 min. A master mix B (reaction buffer, substrate, enzyme) was added to each well and the activity of the enzyme was monitored by measuring the formation of fluorescent metabolites at intervals of 5 min incubating at 37°C after the start the reaction. Standard inhibitors were recommended by the manufacturer.
Results
L. taraxacifolia extract affects cell morphology and inhibits WHC01 cell proliferation
Aqueous L. taraxacifolia extract demonstrated a significant growth inhibitory effect on WHC01 cells. MTT viability assay showed that the IC50 of aqueous L. taraxacifolia on WHC01 was 2.00 ± 0.04 mg/mL. This IC50 is acceptable to use in this model because the concentration is less than the amounts that are normally consumed by people, which is a minimum of 200 mg for commercialized L. taraxacifolia products. The aqueous L. taraxacifolia affected the morphological features of WHC01 cells showing a distinct change in morphology for the 2.0 mg/mL concentration, which represented the IC50 obtained in the preliminary assessment (Fig. 1A–C). Blebbing was observed in the morphology of the cells after treatment with L. taraxacifolia. There was a decrease in cell proliferation after cells were counted with over a 50% reduction in cell population (Fig. 2A, B).
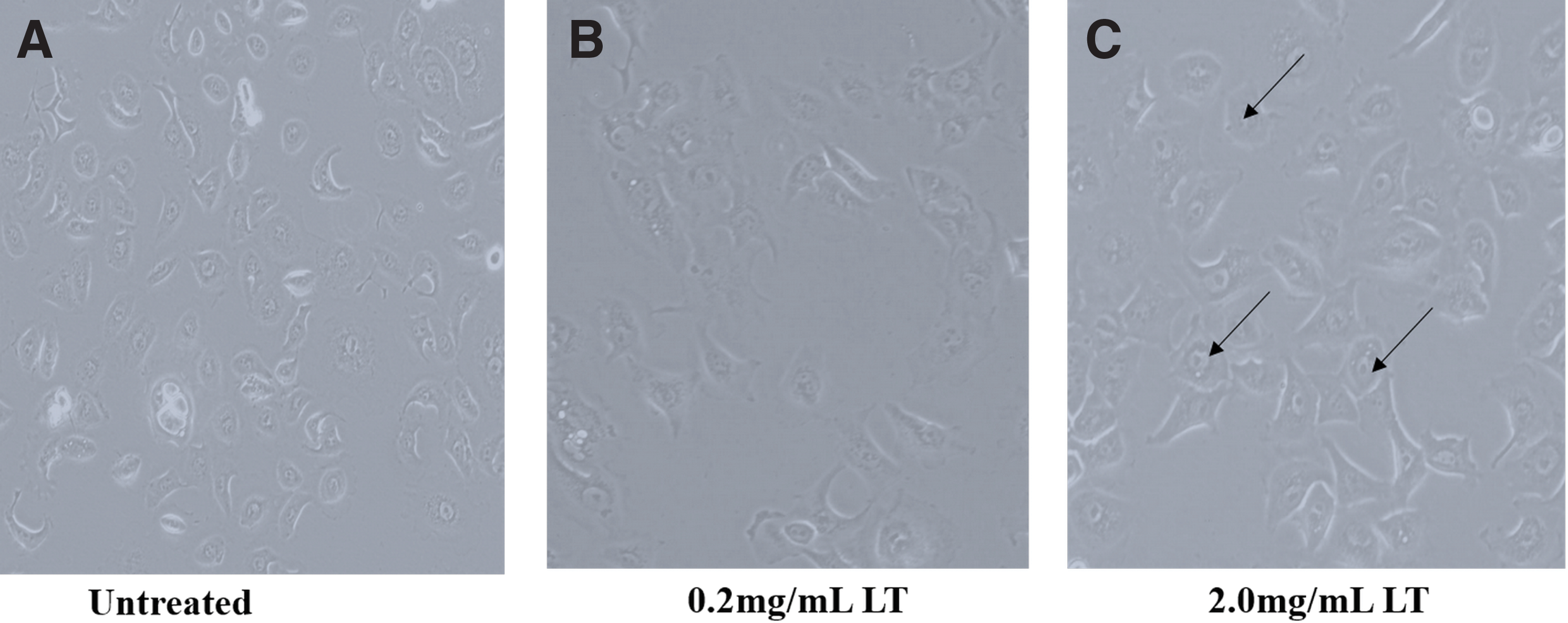
Morphological changes elicited by Launaea taraxacifolia treatment in WHC01 cells. L. taraxacifolia caused a change in the shape of cells compared to untreated controls as shown by the arrows. Blebbing is observed, which indicates signs of apoptosis.
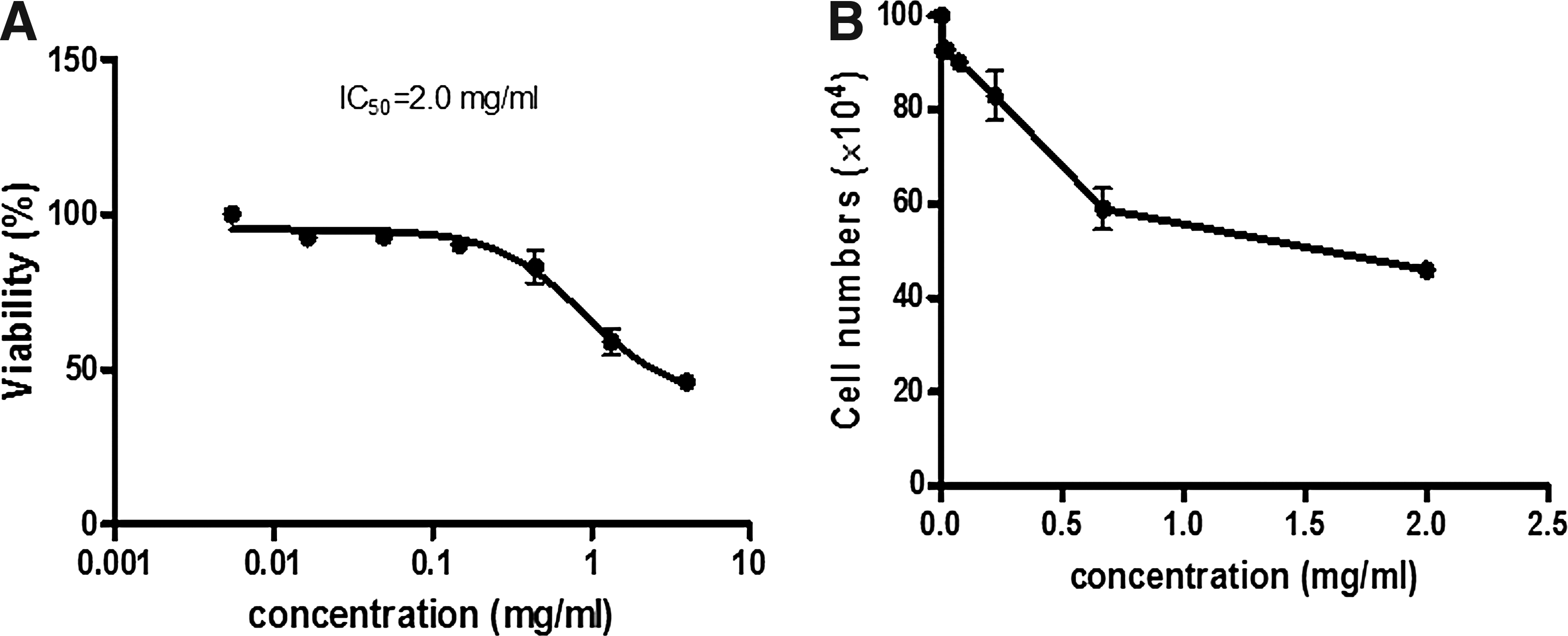
Antiproliferative effects of L. taraxacifolia on WHC01 cells.
Aqueous L. taraxacifolia extract downregulates cell cycle-associated gene expression
Aqueous L. taraxacifolia downregulated the expression of genes involved in cell cycle regulation. Downregulation of expression was observed on cyclin B1 (fold decrease of 60%, p < 0.0046) and proliferating cell nuclear antigen (PCNA; fold decrease of 75%, p < 0.0007) in the presence of the L. taraxacifolia extract (Fig. 3A, B) compared to untreated controls. Expression of senescence markers p21 and p53 showed an increase (2.5-fold, p < 0.0084, and fivefold, p < 0.0023, respectively) (Fig. 3C, D). Several genes known to be expressed by cancer cells were downregulated in the presence of L. taraxacifolia, for example, Bcl-2 (60% decrease, p < 0.006), Bcl-xL (50% decrease, p < 0.025), cathepsin D (30% decrease, p < 0.1085), and matrix metallopeptidase 9 (MMP-9; 80% decrease, p < 0.0001; Fig. 3E−H). These results were substantiated by the Western blot analysis (Fig. 4).
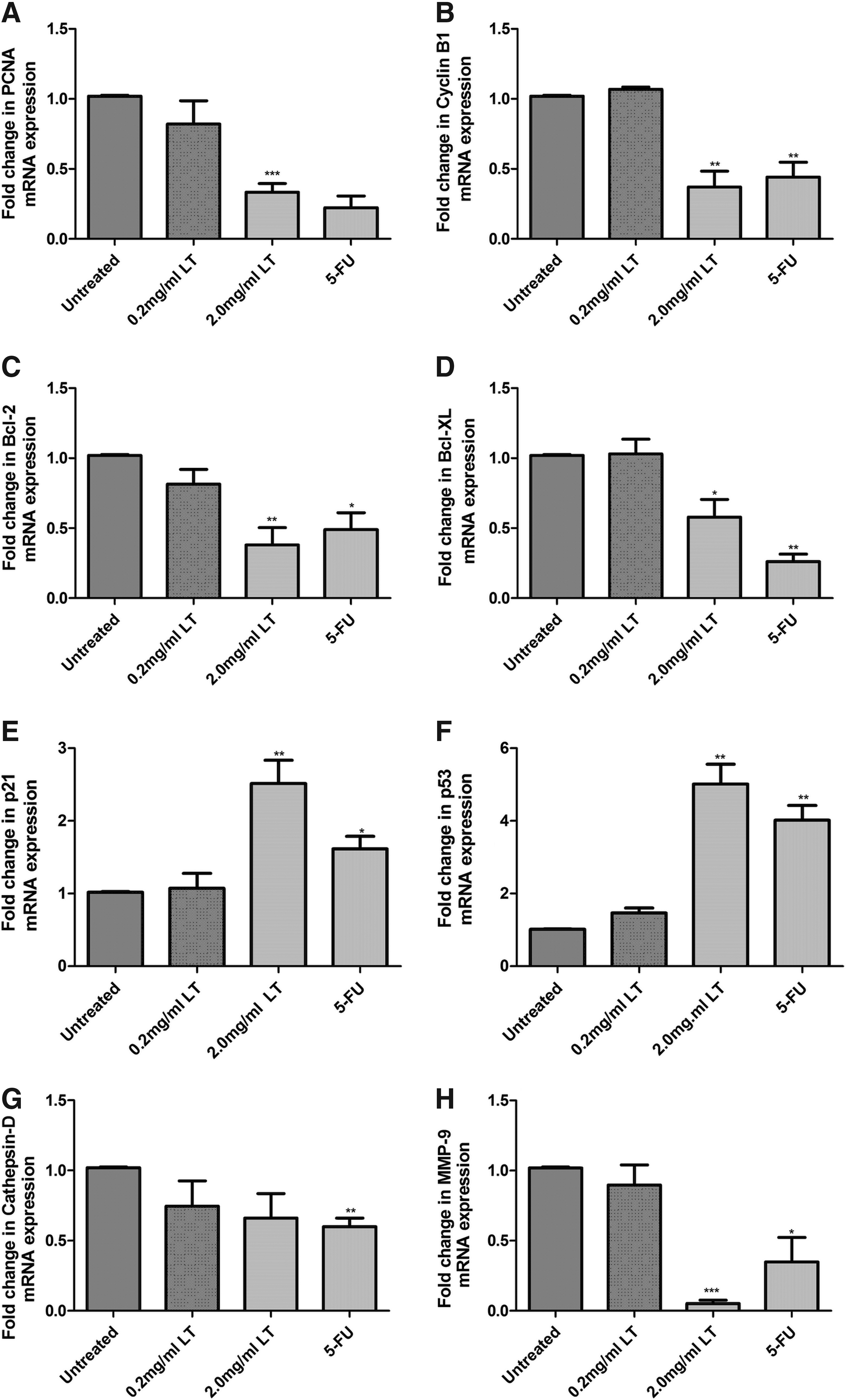
mRNA expression of genes involved in cell cycle regulation.
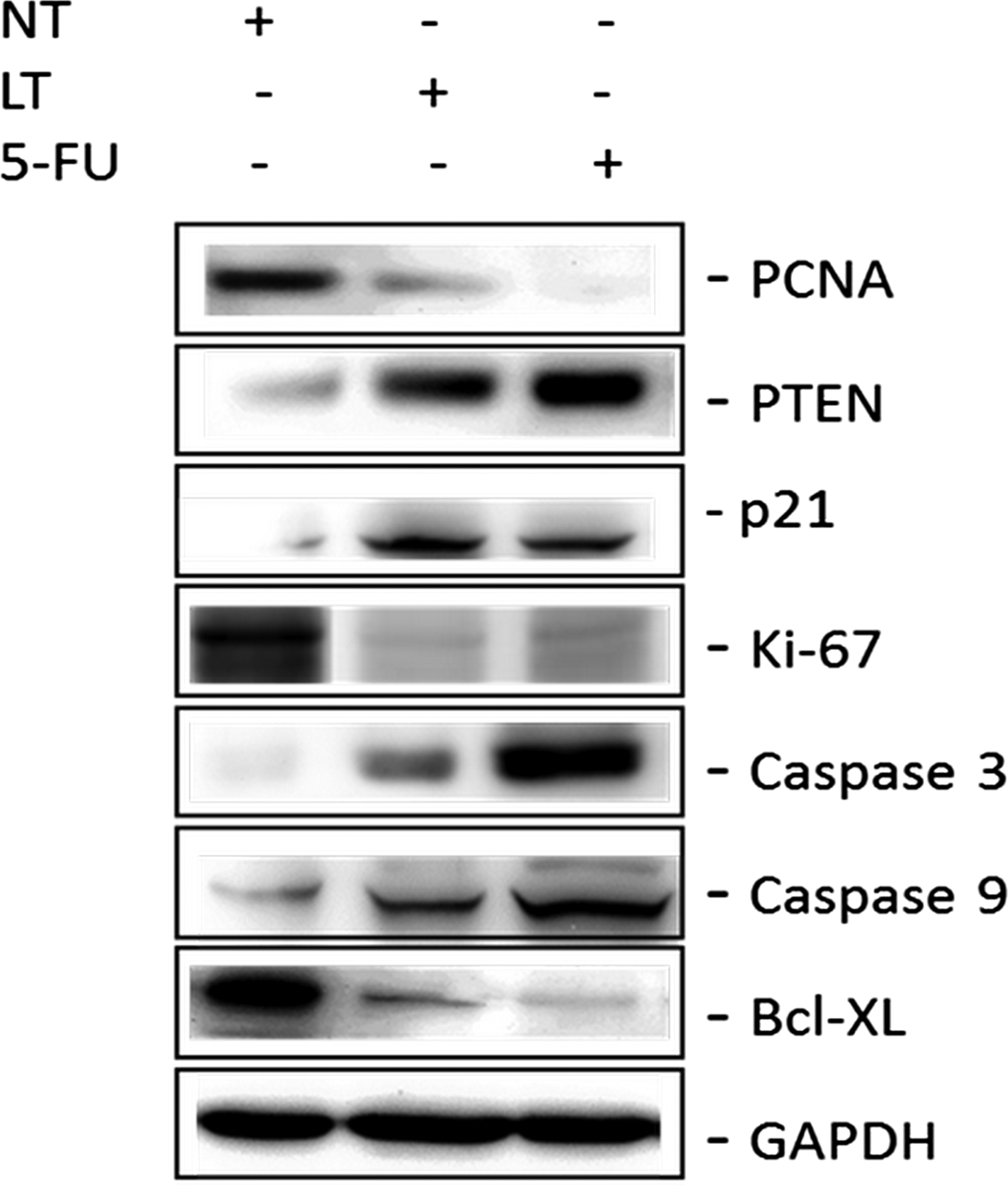
Protein expression of genes involved in cell cycle. Protein blots showing the protein level of PCNA, PTEN, p21, Ki-67, caspase 3, caspase 9, and Bcl-XL detected by respective antibodies. Samples were normalized by their GAPDH expression levels and densitometry analysis performed. 5-FU, 5-flourouracil; LT, Launaea taraxacifolia; NT, no treatment; PCNA, proliferating cell nuclear antigen; PTEN, phosphatase and tension homolog.
L. taraxacifolia promotes G0/G1 cell cycle arrest
Cell cycle analysis measured by flow cytometry showed that L. taraxacifolia extract caused cell cycle arrest at the G0/G1 phase (Fig. 5). There was an increase in cells in the G1 phase from about 51–81% (Table 4). Cell population in the S phase decreased from 41% to 12% in the presence of 2 mg/mL of aqueous L. taraxacifolia. A much lower dose of L. taraxacifolia did not show any effect on cell cycle progression. The observed significant increase in the G0/G1 population after treatment with L. taraxacifolia at the highest concentration suggests that L. taraxacifolia affects the cell cycle at that phase significantly leading to cell cycle arrest.

Cell cycle analysis of L. taraxacifolia-treated cells.
p < 0.05.
G0, resting phase of cell cycle; G1, gap phase I of cell cycle; G2, gap phase II of the cell cycle; LT, L. taraxacifolia; S, synthesis phase.
L. taraxacifolia downregulates CYP450 expression and causes reversible and TDI of these enzymes
Aqueous L. taraxacifolia generally downregulated messenger RNA (mRNA) expression of CYP1A2 (60% decrease, p < 0.005) and CYP2C19 (40% decrease, p < 0.03), but not CYP2C9 in WHC01 cells (Fig. 6) compared to the untreated control. Aqueous L. taraxacifolia showed a reversible inhibition profile on the recombinant human CYP enzymes (Table 5), with a strong reversible inhibitory effect on CYP1A2 (IC50 = 2.33 μg/mL) isozyme and moderate reversible inhibitory effects on CYP2C9 (IC50 = 15.19 μg/mL) and CYP2C19 (IC50 = 13.78 μg/mL) isozymes. The inhibitory kinetics of inactivation on CYP1A2 by L. taraxacifolia were Ki = 2.09 μg/mL and Kinact = 0.016 min−1. Inactivation kinetics for CYP2C9 were Ki = 1.64 μg/mL and Kinact = 0.022 min−1 and for CYP2C19 were Ki = 3.14 μg/mL and Kinact = 0.044 min−1. L. taraxacifolia displayed a TDI effect on CYP1A2 (TDI potency of 1.91), but not on CYP2C9 and CYP2C19.
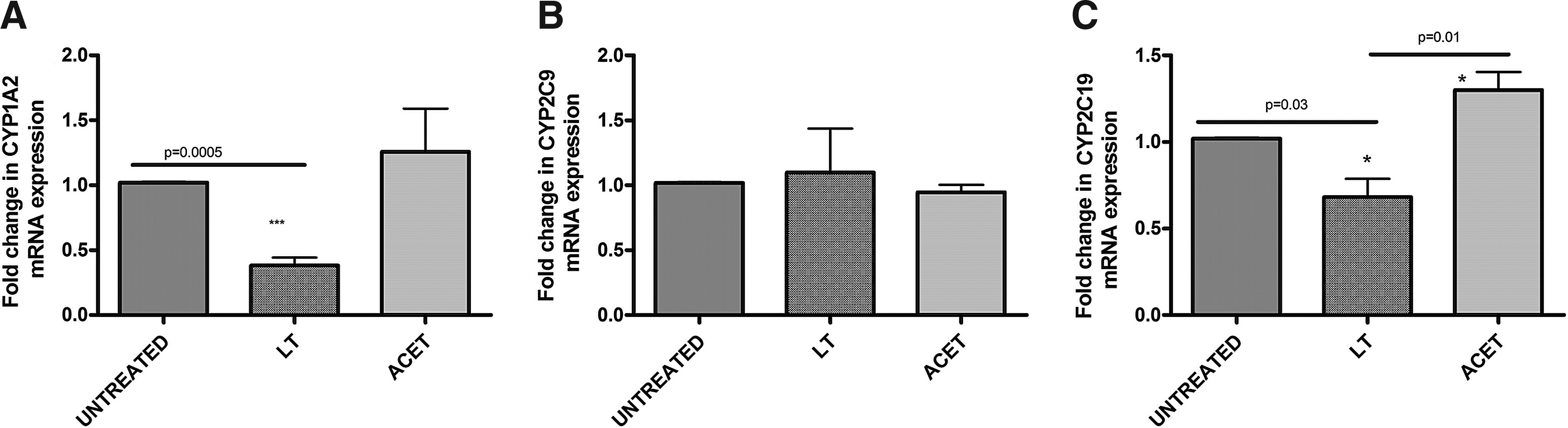
mRNA expression of CYP1A2, CYP2C9, and CYP2C19 genes. Data were expressed as the mean fold mRNA expression relative to the mRNA expression level of untreated controls. GAPDH transcript levels were used to normalize the target transcript abundance. *p < 0.05; ***p < 0.005.
CYP450, cytochrome P450; IC50, inhibitor concentration that reduces 50% enzyme activity/transporter activity; Ki, inhibitor concentration required for half-maximal inactivation; Kinact, maximal rate of inactivation; NADPH, nicotinamide adenine dinucleotide phosphate; TDI, time-dependent inhibition.
Discussion
Precision medicine is an ultimate goal for improved patient management. However, the focus has largely been on therapeutic drugs and not much on other environmental factors including herbal medicinal plants; yet, they play a significant role in different health systems. This study offers the following key observations that have an important relevance in the way L. taraxacifolia may in the future play a role in anticancer therapeutics as well as herb–drug interactions when L. taraxacifolia is used in combination therapy with drugs. It was also observed that L. taraxacifolia extracts affect the proliferation of WHC01 cancer cells, significantly downregulating cancer genes such as bcl-2, bcl-xL, cathepsin D, and MMP-9 and upregulating senescence-associated genes p21 and p53. This has relevance for cancer therapy since the objective is to prevent cancer cells from dividing and eventually causing their death. L taraxacifolia presents a potential source of cancer therapy as a significant number of chemotherapeutic drugs in the market such as vinblastine, doxorubicin, and paclitaxel were harnessed from herbal remedies (Wang et al., 2012).
It was also observed that L. taraxacifolia inhibited CYP450 enzymes and significantly downregulated CYP1A2, CYP2C9, and CYP2C19, which are important pharmacogenes with relevance to chemotherapeutic drug metabolism. This has relevance for herb–drug interactions in chemotherapy since cancer patients partly depend on herbal remedies like L. taraxacifolia for their high antioxidant content to help mitigate some of the side effects associated with cancer drugs (Yarney et al., 2013). Several herbal medicines are used in addition to conventional therapy to improve the efficacy and reduce drug-associated side effects and complications. Use of specific herbal medicines as adjuvants to conventional therapy needs to be evaluated to reduce potential risks of herb–drug interactions. There is evidence of beneficial effects of herbal medicine in managing cancer and alleviating side effects of chemotherapy in patients, and this is attributed to phytochemicals and their high antioxidant activities (Yin et al., 2013). However, there are potential risks that are associated with the combined use of herbal medicines with chemotherapeutic agents due to herb–drug interactions (Thomford et al., 2015; Werneke et al., 2004).
It has been previously reported that crude extracts of various herbal medicines such as Zingiber officinale (Ginger) and Rhazya stricta independently suppress proliferation and induce apoptosis in human breast cancer cell lines, MCF and MB-MDA-231 (Elkady et al., 2012; Nabih et al., 2012). L. taraxacifolia contains high amounts of antioxidants and other phytochemicals (Adinortey et al., 2012), a profile which is believed to give it its medicinal properties. L. taraxacifolia is administered in addition to other conventional medicines, including chemotherapeutic agents, to provide better protection against toxic side effects.
In this study, the effects of L. taraxacifolia as a herbal medicine were assessed in an esophageal cancer cell line WHC01. L. taraxacifolia downregulates the mRNA expression of cell cycle genes PCNA and cyclin B1 that are involved in cell proliferation by over 60%. P21 and p53, genes that are involved in the cell senescence process, were significantly upregulated by over 2.5-fold. Genes that are known to be expressed by cancer cells during progression of the disease, such as Bcl-2, Bcl-xL, cathepsin D, and MMP-9, were all downregulated by over 60%. This explains the reduction in proliferation of the cells.
Flow cytometric analysis indicated an arrest at the G0/G1 phase of cell cycle as a possible mechanism. Also, the protein expression levels of genes involved in cell cycle regulation such as PCNA, Ki-67, and Bcl-xL were downregulated, while phosphatase and tension homolog (PTEN), p21, caspase 3, and caspase 9 were upregulated. These findings provide evidence about the potential beneficial medicinal effects of L. taraxacifolia. When used for the management of cancers, such as esophageal cancer, there is a possibility of reducing the proliferation of cancerous cells. Historically, herbal remedies have been used for therapeutic purposes and with the expansion of pharmaceutical technologies, many lead compounds have been discovered from them. Herbal remedies still remain an important route to new pharmaceuticals. L. taraxacifolia, thus, presents a target for discovery of some novel compounds, which may lead to effective and more tolerable drugs for the management and/or treatment of cancer.
It has been previously demonstrated that activities of CYP450 enzymes enhance chemosensitivity in the human tumor cell lines such as MCF7 (derivative of breast cancer carcinoma) and BxPc3 and T3M4 pancreatic cancer cells lines (El-Ghamari, 2014). The study looked at the effects of L. taraxacifolia on mRNA expression of CYP1A2, CYP2C9, and CYP2C19 in WHC01 cells and its reversible and time-dependent inhibitory profiles using recombinant human CYP450s as recommended by FDA (Zhang et al., 2009) and EMA (European Medical Agency, 2012).
With respect to available commercial herbal medicines, it is important that such studies are conducted for labeling purposes to caution patients taking them for possible herb–drug interactions. We report differential expression of CYP1A2, CYP2C9, and CYP2C19 in the WHC01 esophageal cancer line used in this study. CYP1A2, CYP2C9, and CYP2C19 are known to metabolize drugs, including cancer drugs such as 5-FU (CYP1A2), cyclophosphamide (CYP1A2, CYP2C9, CYP2C19), and thalidomide (CYP2C19).
Based on the expression levels observed in this study, the possibility of herb–drug interaction at the site of the tumor is likely to be through either the activation or deactivation of chemotherapeutic agents when coadministered together.
It is important to understand that where a chemotherapeutic drug requires prior activation from prodrug status to active metabolite, in the presence of inhibitors for such enzymes, this could be compromised; however, in the scenario that the administered compound is the active component, inhibition of drug metabolizing enzymes at cancer site enhances drug concentration and possibly anticancer effects. L. taraxacifolia significantly decreased the expression levels of CYP1A2 and CYP2C19, which implies that at the site of action, dose with a CYP1A2 and CYP2C19 chemotherapeutic substrate will decrease the activity of the enzyme, thus increasing the toxicity of the chemotherapeutic agent at the site. However, L. taraxacifolia did not affect the expression levels of CYP2C9. The reversible and time-dependent inhibitory profiles of L. taraxacifolia showed a strong reversible inhibition of CYP1A2 isozyme with a potential time-dependent inhibitory potency.
The overall effect of new chemical entities causing enzymatic TDI even if there is withdrawal of the compound results in permanent destruction of the enzyme which will require de novo synthesis of new enzyme (Thomford et al., 2016). Thus, should enough L. taraxacifolia be absorbed, the continuous use of L. taraxacifolia by patients allows for its possible accumulation in the cells leading to inhibition of CYP1A2 enzyme and possible accumulation of CYP1A2-dependent drugs, including chemotherapeutic agents, due to reduced enzyme activity. The in vitro IC50 of 2 mg/mL concentration observed in the study is a lesser representation of what happens in vivo as patients who consume commercial L. taraxacifolia take a minimum dosage of 200 mg (Thomford et al., 2016) and for individuals who take the raw parts like leaves, they likely consume excessively more.
Therefore, the extent of the observed effects in the study depends on the bioavailable fraction that will be absorbed, which is also dependent on the phytoconstituents and the processing of the extract. It is important to realize that phytoconstituents for most herbal plants are dependent on the geographical area from which each plant is harvested; thus, it will be important to carry out studies comparing profiles of constituent components of the same herbal plant grown in different regions. It was observed that there was a moderate reversible inhibitory potency for CYP2C9 and CYP2C19 with no TDI potency observed for CYP2C9 and CYP2C19, implying that the inactivation of CYP2C9 and CYP2C19 by L. taraxacifolia extracts at the enzyme level is likely to be transient since the inhibition can be overturned to allow for enzyme function to be restored if there is withdrawal of L. taraxacifolia use.
This study is important for cancer therapy since some patients are continuously using herbal medicines in addition to conventional medicines (Adejuwon et al., 2015; Nasr and Saleh, 2014; Thomford et al., 2015) in the hope of being cured. The challenge is that most chemotherapeutic drugs present with side effects; thus their co-use with herbal medicines such as L. taraxacifolia may result in potentially increased toxicity for some substrates. One of the major problems is that patients seldom disclose to their physicians' herbal medicines they are currently taking, in addition to their prescribed drugs. Thus, for populations in health transition such as Africa that presents with a huge disease burden, it is imperative such studies be conducted to provide information to be incorporated into the health delivery system. There are still questions remaining, for example, the effects of herbal plants on a patient's microbiome. Microbiome profiles are increasingly being identified as possible biomarkers of disease susceptibility or treatment response.
Conclusions
L. taraxacifolia causes the arrest of cell cycle at the G0/G1 phase by affecting differential expression of genes involved in cell cycle regulation, presenting its potential beneficial effects. Generally, changes in gene expression levels that accompany the administration of herbal medicines play a vital role in treatment options available to patients in a population in health transition. Chemotherapy is one of the effective methods of treating cancers such as esophageal cancer. However, with the pharmacokinetic and pharmacodynamic interaction involved in herb–drug metabolism, it is important to advice patients when coadministering herbal medicines such L. taraxacifolia and cancer drugs.
Footnotes
Acknowledgments
Research reported in this publication was supported by the National Research Foundation (NRF) under an Indigenous Knowledge Systems Research Grant and partly, the pharmacogenetic component, by the South African Medical Research Council (SAMRC) under a Self-Initiated Research Grant. The views expressed are personal opinions of the authors and do not necessarily reflect those of their affiliated institutions or the funders. N.E.T. is funded by an NRF grant-holder linked bursary.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.