
Abstract
Abstract
Glioblastoma multiforme (GBM) is one of the most lethal brain tumors with an inadequately understood pathophysiology. Biomarkers that guide accurate diagnosis and treatment decisions would greatly support precision medicine for GBM. Previous studies of GBM have focused on signaling pathways such as epidermal growth factor receptor (EGFR), platelet-derived growth factor receptors (PDGFRs), notch, wnt, and others, identified with single omics technology platforms (genomics, transcriptomics, or proteomics), but not with their integrated use. In this context, we report here a multi-omics pathway view, expanded through integration of the expression data at transcriptomic and proteomic levels, followed by selection of a functionally related group of proteins such as kinases deregulated in GBM. By using this strategy, we observed a highly significant enrichment of the gonadotropin-releasing hormone (GnRH) signaling pathway that was not deciphered with single omics datasets. The curation of the GnRH pathway with extensive literature analysis brought about a comprehensive annotation of the pathway, which included several additional pathway members that were not previously annotated. A targeted search resulted in identification of additional nonkinase members of the pathway in the GBM multi-omics datasets. We found evidence of GnRH receptor expression in GBM and other cancers. We offer here an updated generic pathway map of GnRH signaling, show its enrichment in the context of GBM, and discuss its plausible cross-connectivity with EGFR, wnt, calcium, and focal adhesion kinase signaling pathways that were earlier shown to be the top deregulated pathways in GBM. In conclusion, this study demonstrates the promise of multi-omics research and analyses to better understand complex cancers and suggests continued efforts and research in this direction in the field of integrative biology.
Introduction
G
Unbiased genomic, transcriptomic, and proteomic analyses applied to clinical specimens have provided extensive molecular information of GBM etiology (Deighton et al., 2010; Dong et al., 2010; Kalinina et al., 2011; Cancer Genome Atlas Research Network, 2008). These studies have identified significant pathways and processes to be deregulated in GBM and high-grade gliomas. Some of the major deregulated pathways include receptor tyrosine kinases (RTKs) such as epidermal growth factor receptor (EGFR) and platelet-derived growth factor receptors (PDGFRs) regulating tumor growth, pathways such as P53 and RB modulating DNA repair and cell survival, and those involved in tumor differentiation such as notch, wnt, and calcium signaling among others (Brennan et al., 2013; Cerami et al., 2010; Dong et al., 2010; Polisetty et al., 2012; Cancer Genome Atlas Research Network, 2008).
One of the key observations is the altered expression of many calcium regulatory molecules, including integrins, annexins, S100 proteins, CAM kinases, and voltage-, ion-, or nucleotide-gated calcium channel proteins that are cross-linked to RTK and G-protein-coupled receptor (GPCR) pathways, suggesting deregulated calcium signaling to be an integral part of GBM tumorigenesis (Leclerc et al., 2016; Liu et al., 2011; Polisetty et al., 2012). However, given the limitations of detection and biases involved in proteomic and transcriptomic analyses and the differences in the regulation of steady-state levels between the two, not all the altered entities can be identified uniformly at both levels. A combined approach to view perturbations at both transcriptomic and proteomic levels can thus provide better coverage of the pathway entities used to map pathways.
Identification of concordant molecular entities increases the confidence of the molecular changes observed for biomarker applications (Gupta et al., 2015). Combining the information from such high-throughput multi-omic studies for mapping signaling pathways and networks also provides an enhanced biological context of the tumor (Higdon et al., 2015; Jayaram et al., 2014; Karagoz et al., 2015). The molecular information can also be integrated with clinical data to characterize disease phenotypes and generate clinically more efficient molecular targets for novel diagnostics and therapeutics (Higdon et al., 2015). We note that the pathway information in the present databases has scope for updates and curation by incorporating new annotations, which would bring in additional entities from experimental datasets. Targeting specific protein families for pathway mapping may add further advantage as we can mine the datasets for specific interactors/regulators of these proteins mapping to the pathways.
Protein kinases (PKs) are a family of proteins that are major components of signal transduction pathways acting as membrane receptors (RTKs) or as intracellular signaling mediators (nonreceptor PKs) and several PKs have been implicated in gliomagenesis (Nakada et al., 2013; Pandey et al., 2016). Several studies have also shown altered expression of PKs in GBM, and targeted therapies directed toward RTKs using kinase inhibitors are in clinical trials (Berezowska and Schlegel, 2011; Pillay et al., 2009). There is renewed optimism in the use of kinase inhibitors to treat GBM (Mellinghoff et al., 2012). New therapeutic strategies have emerged that use multitargeted kinase inhibitors to simultaneously disrupt multiple kinases (Sathornsumetee and Reardon, 2009). Hence, we have used the kinase group of proteins, which are differentially expressed to map to pathways to explore newer pathway information, and identified gonadotropin-releasing hormone (GnRH) signaling pathway as the top pathway. We discuss its implications in the context of GBM and cross-connectivity with other pathways involved in GBM.
Methodology
Integrated analysis of GBM proteomic and transcriptomic datasets
We carried out a detailed comparison of protein datasets with published transcriptomic data. The GBM transcriptomic data were obtained from The Cancer Genome Atlas (TCGA) that is made freely available through the Anduril website (http://csbi.ltdk.helsinki.fi/anduril/tcga-gbm), while the mass spectrometry-based proteomic data used were generated earlier from our laboratory (Polisetty et al., 2012). Supplementary Tables S1A and S1B give the list of proteins and transcripts identified in GBM clinical tissues along with their respective fold change values. All entities with expression levels greater than 1.4-fold were considered for analysis. Ingenuity pathway analysis (IPA) and GeneSpring Pathway Architect (version 12.6) software were used for the present pathway analysis and multi-omics analysis.
Pathway analysis of differentially regulated kinases in GBM
Furthermore, to examine the validity of observations made at proteomic and transcriptomic levels, we selected functionally related family of proteins such as kinases in GBM. We identified 102 differentially regulated kinases in GBM proteomic (Polisetty et al., 2012) and transcriptomic datasets (Dong et al., 2010) to identify the top canonical pathways through IPA. The list of the identified kinases in GBM omics datasets used for IPA is provided in Supplementary Table S2. The GnRH pathway was revealed to be the top pathway with a high significance score in this analysis. To show full enrichment, additional nonkinase entities from GBM omics data that are also part of the GnRH signaling pathway were identified. This enabled better visualization of the interplay and potential role of GnRH signaling entities in GBM.
Building a comprehensive map of GnRH signaling
Although the GnRH signaling pathway was revealed to be the top deregulated pathway in our analysis, a detailed well-annotated map of the GnRH signaling pathway was lacking in the current pathway resources. The pathway provided by IPA resource was also sparsely annotated. Similarly, the GnRH signaling pathway shown in KEGG resource (www.genome.jp/kegg-bin/show_pathway?hsa04912) had some incomplete signaling reactions downstream of PKC, possibly due to lack of information at the time the pathway was created. Hence, before we mapped the omics data in GBM on this pathway, it became essential to construct a well-annotated generic pathway map of GnRH signaling. We did so by incorporating information from literature pertaining to GnRH.
A thorough literature search was carried out using GnRH and signaling as keywords in PubMed and Google Scholar. The keyword searches revealed more than 2000 research articles in PubMed database. Information and references listed in the review articles on GnRH signaling were also taken and manually studied for annotations. All articles were first screened for specific information pertaining to GnRH signaling such as protein–protein interactions (PPIs), post-translational modifications (PTMs), activation/inhibition events, translocations, and gene regulatory events. Only ligand-stimulated, receptor-mediated signaling events and experimentally proven reactions were considered for curation. The source species for all reactions was normal or neoplastic human cells. However, where information was not available in human systems, experiments done in other mammals such as mouse, rat, and monkey were also considered for annotations (such information contributed only small component of curation). To increase the confidence of particular interactions, reactions supported by multiple experiments or supported by different research articles were given more weight for annotation of the pathway. For PTM events, information on specific site and residue altered was included, where available.
Screening, thus, resulted in 250 research articles that were used for curation and building an enhanced view of the pathway. Following curation, several entities were newly added to the existing GnRH signaling pathway annotation in KEGG and IPA pathway resources. A freely available pathway drawing software called PathVisio (version 3.1.3) was used for drawing the curated pathway map. All pathway reactions were further reviewed by a pathway authority, an expert in the field (R.R., a coauthor in this article). The fully curated generic map of GnRH signaling is provided in Supplementary Figure S1. We mapped the differentially expressed protein and transcript entities from the GBM datasets to this curated pathway map of GnRH signaling.
Results
We have integrated differentially expressed transcriptomic and proteomic data of GBM and carried out pathway analysis using the kinase group of proteins and their interactors. The overall workflow of the analysis is shown in Figure 1. To assess the overlapping transcripts and proteins identified for GBM, we compared all entities with greater than 1.4-fold change from differentially expressed proteomic data [n = 1086; Polisetty et al. (2012)] and the gene expression-based transcriptomic data [n = 1628; Ovaska et al. (2010) and Cancer Genome Atlas Research Network (2008)] from GBM. A total of 711 entities were found to be common between the two omics datasets, of which 670 (94%) were concordant with respect to the trend in expression, as shown in the Venn diagram in Figure 2.
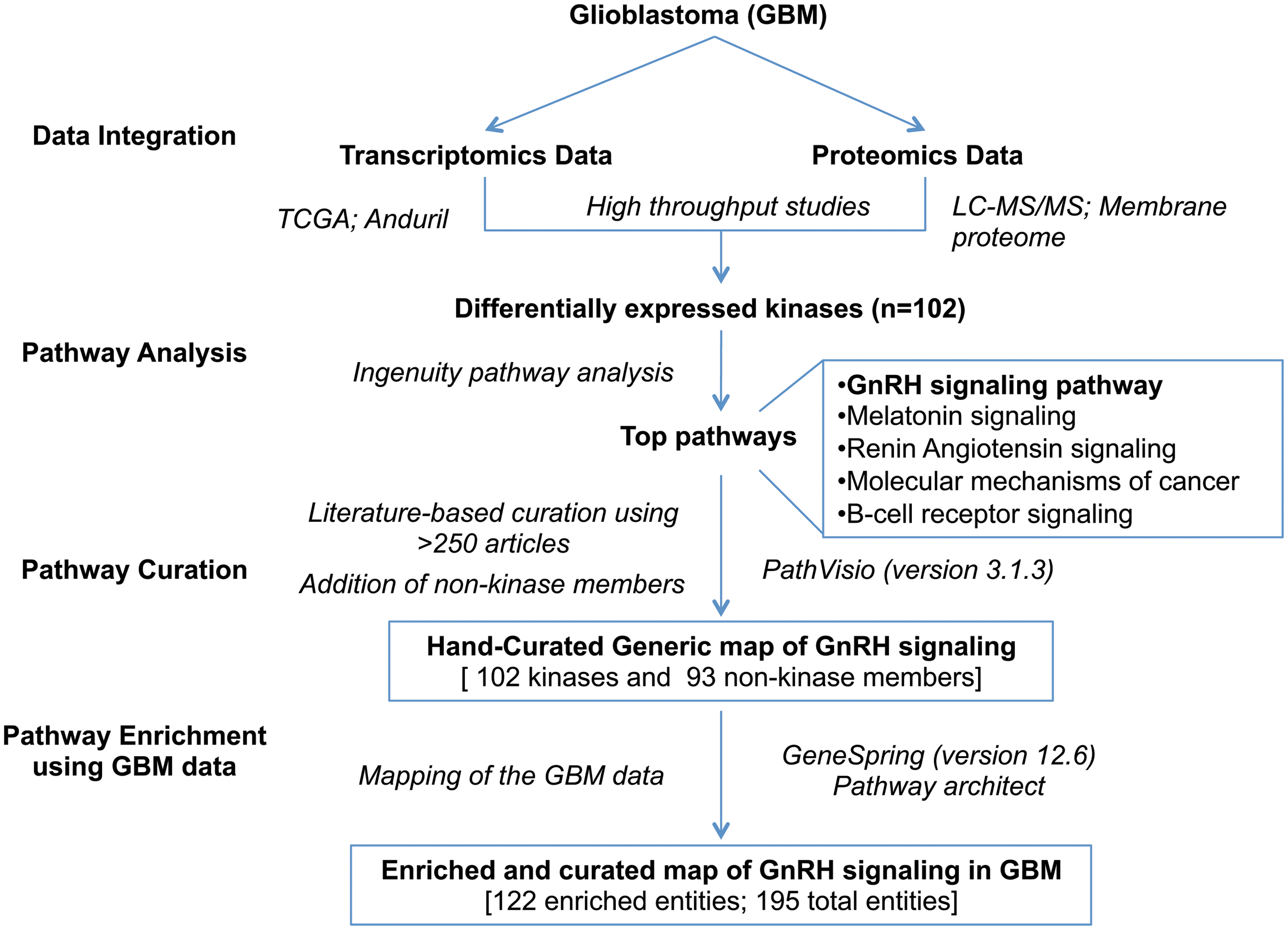
Workflow of the study and analyses. The overall workflow of the analysis is depicted in this figure.
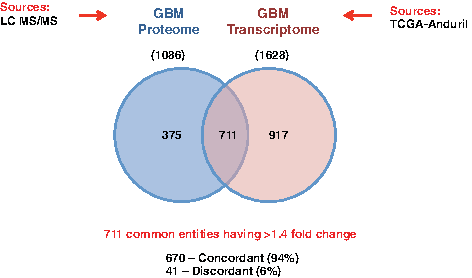
Common entities between omics datasets. This Venn diagram shows the common entities between GBM proteomic and transcriptomic datasets and the degree of concordance between them. GBM, glioblastoma multiforme.
The degree of correlation (regression value R2 = 0.725) between the common entities in the omics datasets also reveals high concordance between them as displayed in the scatter plot in Figure 3. However, when we compared the pathway analysis carried out with single ome datasets (Polisetty et al., 2012; Cancer Genome Atlas Research Network, 2008), the coverage of the altered entities in the pathways was not as uniform. Hence, we integrated the two datasets and used them for pathway analysis. This increased the coverage of entities and significance of some of the pathways. Furthermore, we selected PK group of entities that are differentially regulated in the two datasets for mapping to pathways. Kinases are well-known therapeutic targets in cancers. The GBM data were found to be enriched with several kinases. We identified a total of 102 kinases in GBM datasets; 77 in transcriptomic data and 30 in proteomic data, 26 common between them.
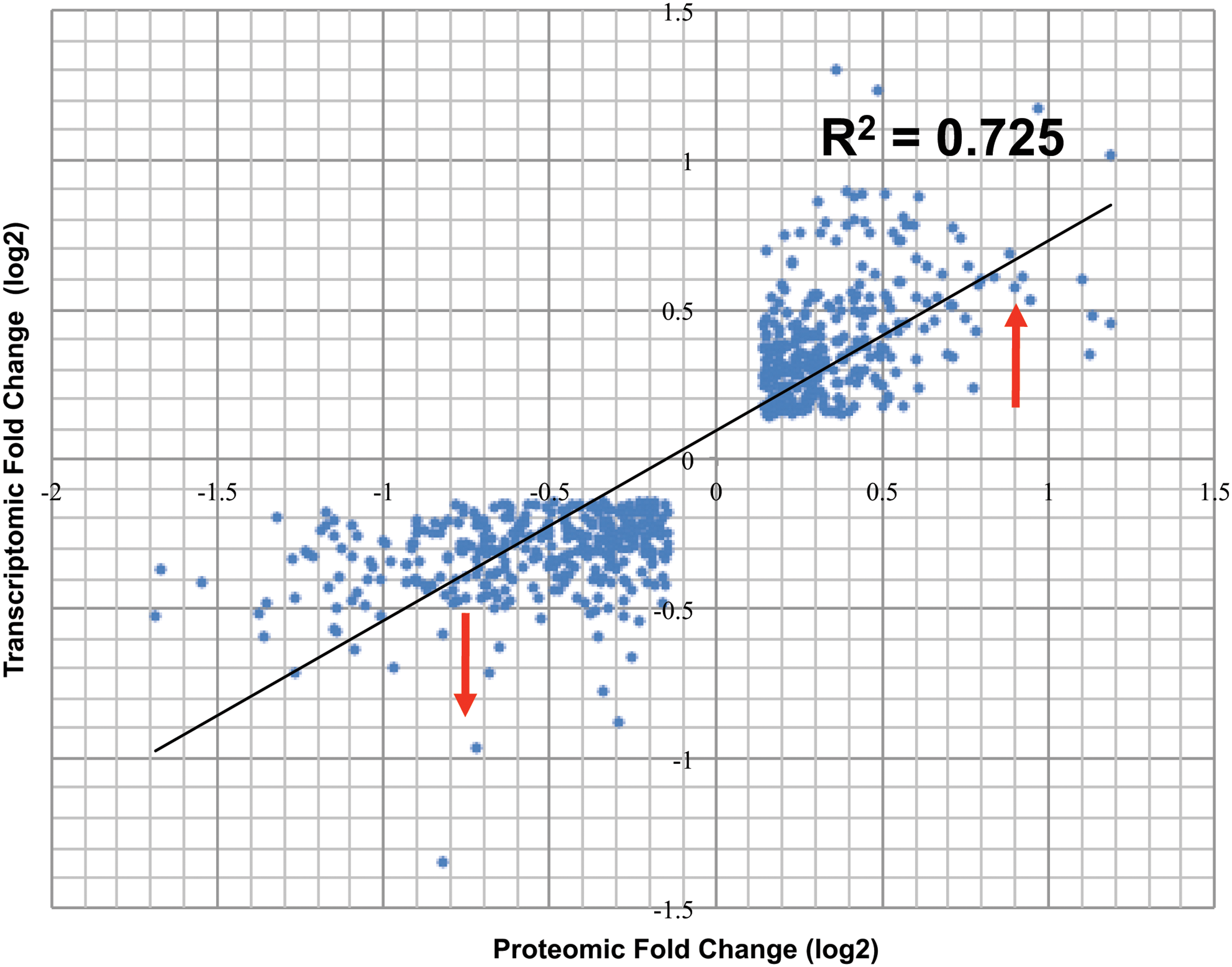
Concordance between omics datasets. Scatter plots showing the degree of correlation (R2) between the common and concordant entities (n = 670) in the two datasets.
This integrated dataset was mapped to pathways using the IPA tool. All the significant pathways identified using IPA are shown in Figure 4. We observed the GnRH pathway to be the top pathway in this analysis with high significance p-value (1.27E-40). The other pathways included melatonin signaling, renin–angiotensin signaling, molecular mechanisms of cancer, and B cell receptor signaling. We also carried out analysis using the subset of genes that conform to 2.0-fold change as the threshold commonly used (74 kinases from the transcriptomic data and 26 from the proteomic data) and observed enrichment of the same pathways (data not shown). However, since even smaller changes in kinase levels or activities may impact their downstream effects, we believe the analysis using all kinases with 1.4-fold change or higher would offer a more complete view. Addition of other nonkinase members of the pathway from the GBM datasets further increased its significance value.
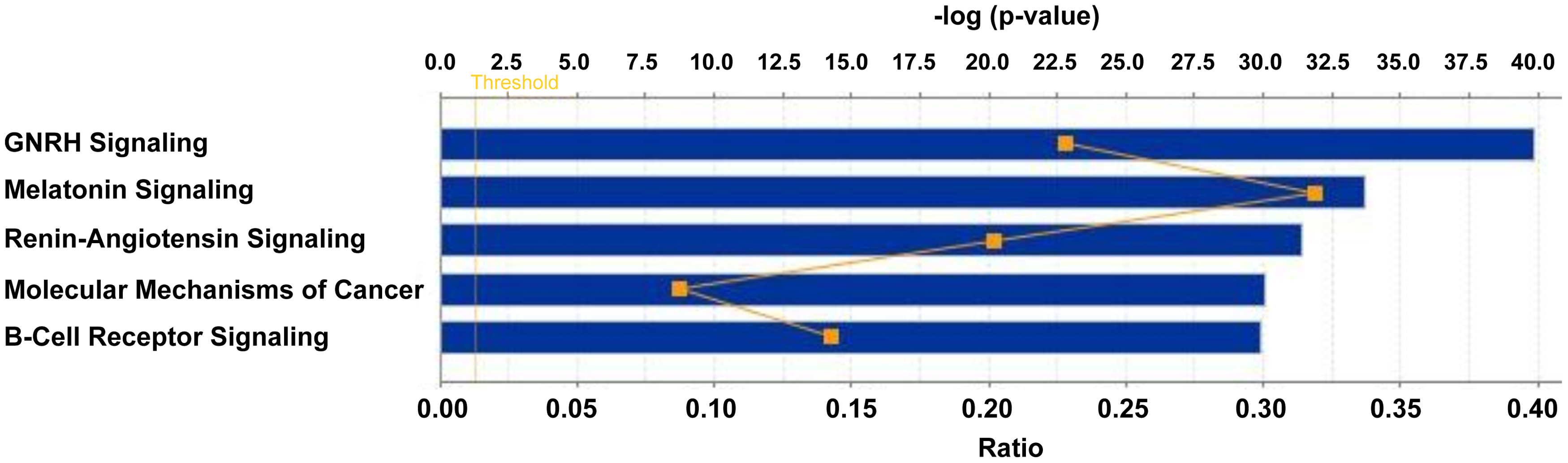
IPA of kinases dysregulated in GBM. The figure shows the top canonical pathways enriched among the dysregulated kinase data mined from GBM omics data. IPA, ingenuity pathway analysis.
To have a total view of all entities (from the integrated datasets) implicated in the pathway, we then built an updated/curated generic pathway map of the GnRH pathway, as described in the Methodology section. Over 2000 research articles pertaining to GnRH signaling were screened, 250 of them were found to provide new information useful for further annotation and curation of the existing GnRH signaling pathway. Information on PPIs, enzyme-catalyzed events, protein translocation, site-specific PTMs, and activation/inhibition events upon activation of GnRH signaling by its ligands was annotated. On annotation, a total of 195 entities are now represented on the map, including about 92 entities already represented in the KEGG map for the GnRH signaling pathway.
Thus, more than 100 entities have been newly added through updated annotations. Some of the new entries in the generic pathway include members that may provide a connecting link with wnt, calcium, integrin/focal adhesion kinase (FAK), and NFkB signaling that are reported to be the major pathways involved in GBM (Brennan et al., 2009; Leclerc et al., 2016; Natarajan et al., 2003; Polisetty et al., 2012; Cancer Genome Atlas Research Network, 2008). Wnt signaling members are now an integral part of the GnRH pathway with beta-catenins, GSK3B, and matrix metalloproteinases (MMPs) directly or indirectly regulated by GnRH (Chou et al., 2003; Gardner and Pawson, 2009; Gardner et al., 2007, 2010; Ling Poon et al., 2011). Apart from the known GPCR and mitogen-activated protein kinase (MAPK) signaling, the fully curated GnRH pathway map now includes several newly added members of the calcium signaling pathway such as L-type voltage-, ion-, or nucleotide-gated Ca2+ channels, annexins, S100 proteins, calcium/calmodulin-dependent kinases (CAMKs), and phosphatases such as calcineurin-regulating NFAT transcription factors (Binder et al., 2012; Haisenleder et al., 2003; Mulvaney et al., 1999).
Key members of FAK signaling such as integrins, actins, filamins, vinculin, and others that form focal adhesion complexes in response to GnRH activation were added to the pathway (Dobkin-Bekman et al., 2009; Maudsley et al., 2007). Evidence now strongly supports the release of arachidonic acid upon GnRH stimulation, followed by induction of COX1 and COX2 intermediates through NFkB (Kiesel et al., 1986; Naidich et al., 2010), which also mediates the antiproliferative effects of GnRH (Meyer et al., 2013). Independently, GnRH is also reported to stimulate several downstream transcription factors such as Nur77, SF1, ATF3, PIN1, PITX1, EGR1, and others that have been newly added to the pathway (Hamid et al., 2008; Luo et al., 2010; Purwana et al., 2011; Xie et al., 2005). A generic pathway map of GnRH signaling was thus created using PathVisio (version 3.1.3) and is provided in Supplementary Figure S1.
The integrated omics data from GBM were then overlaid on the curated generic GnRH map using the GeneSpring Pathway Architect tool and the multi-omic analysis workflow (version 12.6). We observe an overall enrichment of about 122 entities, of which 26 kinases and 57 nonkinase members are coming from the concordant transcriptome and proteome datasets. The corresponding expression of entities at transcript and protein levels (or at both levels) in GBM clinical tissues is shown in different colored boxes in the enriched and curated map of GnRH signaling in Figure 5. The up and downregulation of entities is indicated in red and blue bulbs, respectively, next to the boxes.
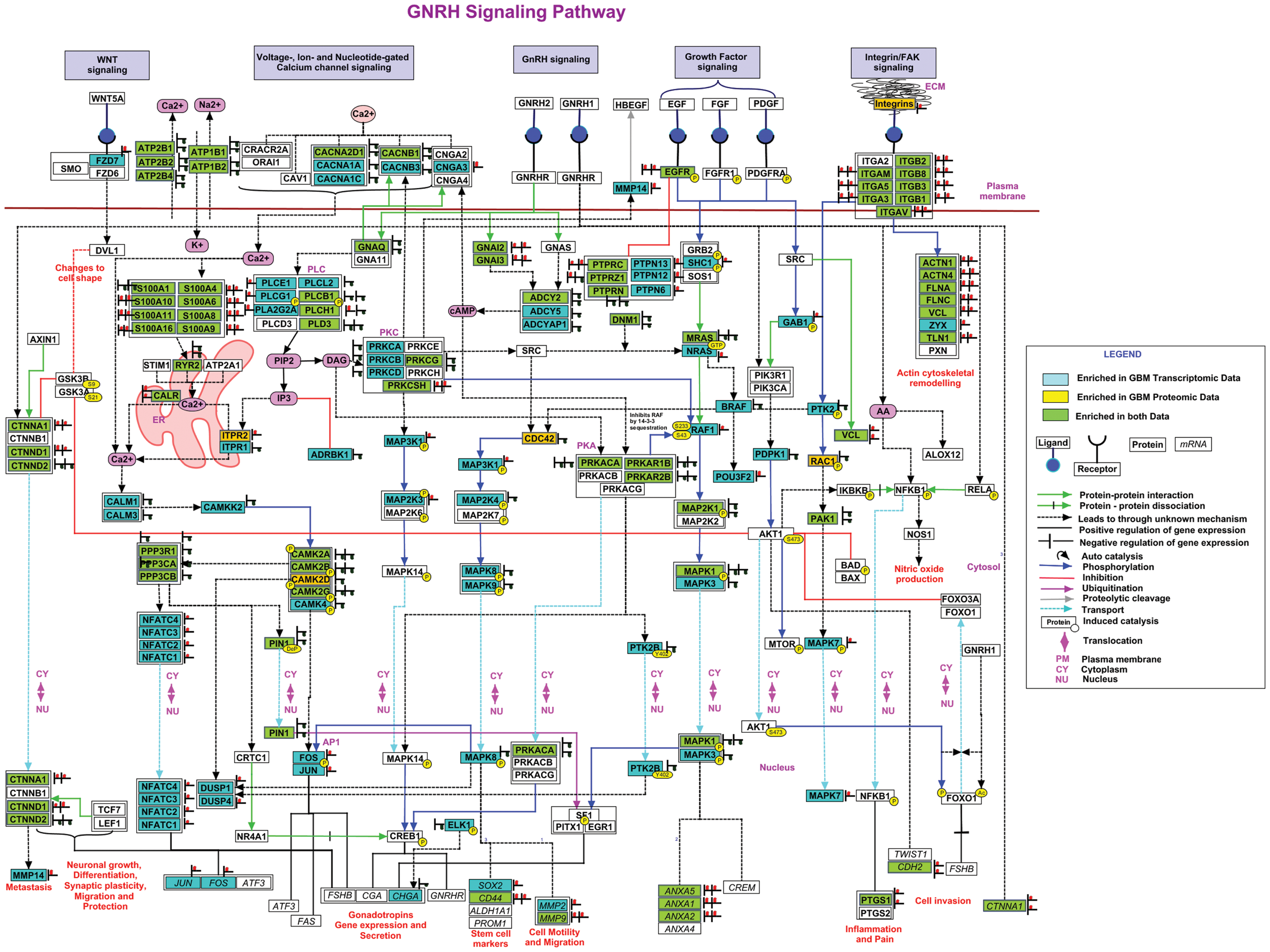
Enriched GnRH signaling pathway in GBM. Differential data from different omics datasets have been manually mapped to the generic GnRH pathway to identify the correlations between transcripts and protein entities and show the enrichment of the pathway.
Thus, we demonstrate how an integrated approach using omics datasets and targeting specific families of proteins such as kinases combined with a comprehensive annotation of the pathway adds further value to the pathway view.
Discussion
GBM is one of the most devastating brain tumors with incompletely understood pathophysiology. Biomarkers that guide early and accurate diagnosis and treatment decisions would greatly support precision medicine for GBM. The current pathway-based annotations fall short to explain the role of less studied genes, identify their functional links, or identify disease-specific cross talk with other entities in the pathway. Furthermore, several pathways may lack sufficient coverage at individual ome levels due to limiting depth of analysis, technical challenges in experimentation, or due to suboptimal annotation of pathway entities. Integrated analysis of multi-omics data is thus useful to understand the cumulative changes in gene expression at multiple levels and leverage their combined potentials.
In the present study, we integrated transcriptomic and proteomic data on GBM to illustrate the concordance and correlation between differentially expressed transcript and protein datasets. This was followed by pathway analysis focused on differentially expressed group of proteins such as kinases present in the two datasets, which resulted in the GnRH signaling pathway as the top pathway altered in GBM. An exploration of GnRH signaling in current pathway resources showed that a well-annotated map of the GnRH signaling pathway was needed. Hence, based on annotations from published literature, we constructed a curated generic pathway map of GnRH signaling to capture all the reactions. The integrated GBM data were then used to enrich the hand-curated generic map of GnRH signaling.
It was intriguing that although GnRH signaling was identified in the pathway analysis for GBM based on global transcriptomic or proteomic data, it did not score high significance. This could be due to incomplete annotation of the pathway in the current resources or due to limited number of relevant entities in the individual ome datasets. Thus, it is important to first generate extensively curated maps and enrich them with all oncogenic alterations. In addition, the use of alternate approaches to enhance pathway views may increase the significance of particular pathways that has led to the identification of a less explored GnRH signaling pathway in GBM.
GnRH is a key neuroendocrine regulator of reproductive functions and secreted by a small subset of GnRH-secreting neurons in the hypothalamus. It binds to and stimulates GnRH receptors present in pituitary gonadotropes to regulate the release of the gonadotropins, luteinizing hormone, and follicle-stimulating hormone. GnRH receptors are a special type of seven-transmembrane GPCRs that primarily use G proteins for downstream signaling and activate MAP kinase cascades (Naor, 2009; Naor et al., 2000). The molecular mechanisms of GnRH signaling may be different in extrapituitary tissues and thus may be important to understand them further in the context of cancerous tissues and GBM in particular (Mermelstein et al., 2003).
Increased GnRH and GnRH receptor (GnRHR) expression has been observed in several hormone-dependent tumors such as breast, endometrial, ovarian, and prostrate tumors (Imai and Tamaya, 2000) and other nonhormone-dependent tumors, including brain tumors (Bahk et al., 2008; Montagnani Marelli et al., 2009; Moretti et al., 2008; van Groeninghen et al., 1998). GnRH receptor activation affects cell survival, proliferation, adhesion, and angiogenesis to promote tumorigenesis [Reviewed in Bahk et al. (2008)]. Unlike in pituitary cells, in extrapituitary tissues and cancers, GnRHR was shown to couple to Gi (GNAI), resulting in activation of other signaling cascades, including MAPK, PI3K, or NF-κB (Aguilar-Rojas et al., 2016). In GBM, Gi may be a major mediator of GPCR/GnRH signaling and acts through coupling to a Gi-cAMP intracellular signaling pathway (rather than the Gq/11-PLC route), resulting in reduction in cAMP levels (Limonta et al., 2003; Navarro et al., 2003). Consistent with these reports, we observed GNAI2 and GNAI3 (Gi) overexpression in GBM datasets while GNAQ (Gq/11) was downregulated. Lower levels of cAMP are also consistent with downregulation of cAMP-dependent PKs, PKA (PRKACA, PRKAR1B/2B) and PKC (PRKCG, PRKCSH), observed in our datasets.
EGFR, calcium, integrins, FAK, and wnt signaling have been extensively researched in GBM (Chin and Meyerson, 2008; Leclerc et al., 2016; Lee et al., 2016; Natarajan et al., 2003). Our observation that the GnRH signaling pathway could be unraveled by using alternate pathway analysis using integrated omics data on GBM permits us to discuss the cross talk of GnRH signaling with these major pathways implicated in GBM and provides leads for better understanding of molecular mechanisms. Using some key entities in the curated map of GnRH signaling, in Figure 6, we show a schematic representation that permits us to speculate cross talk between these pathways and their mechanistic implications on the tumor hallmarks.
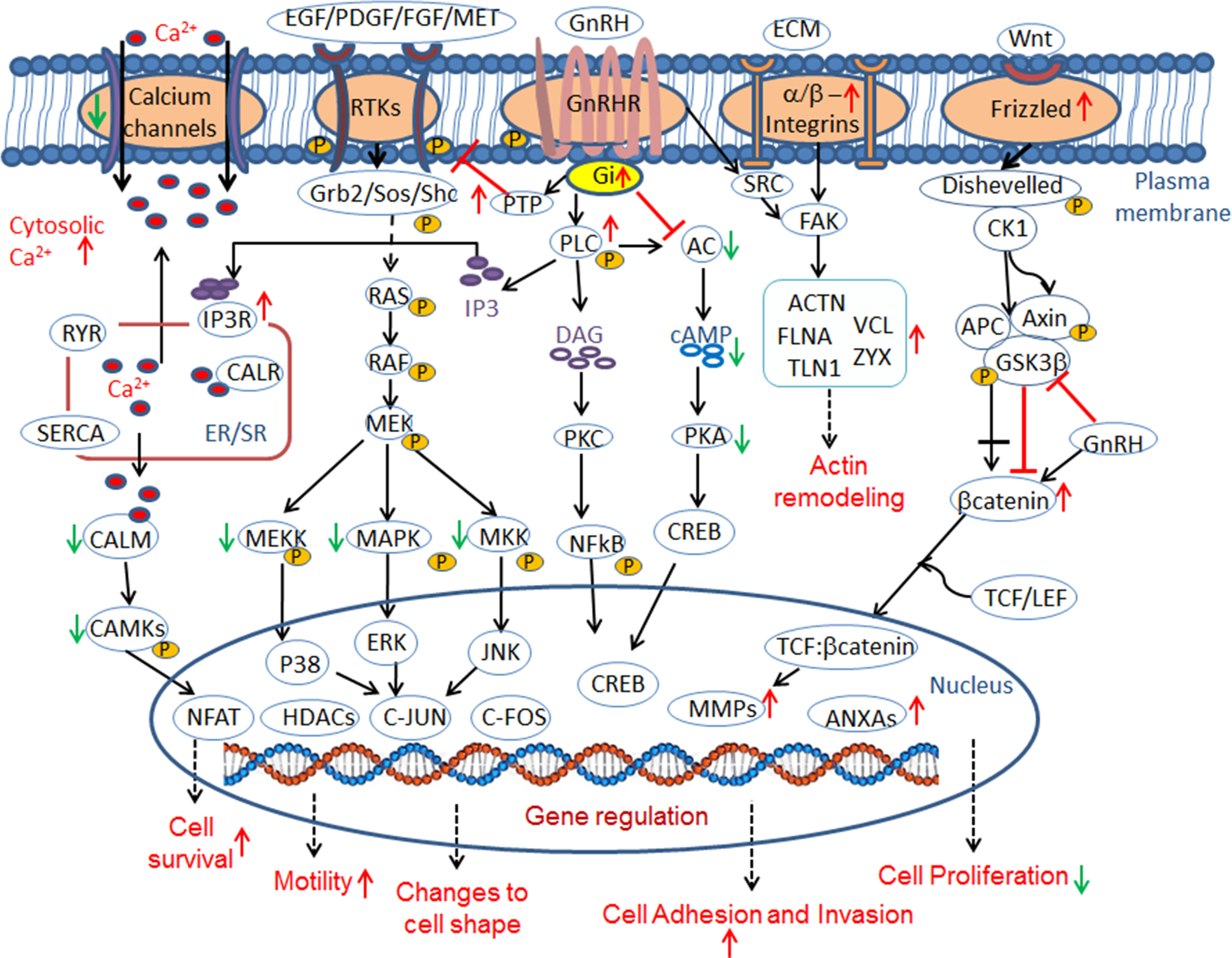
Schematic diagram of GnRH signaling. The figure represents a schematic of the reactions and cross talk in the GnRH signaling pathway.
Global pathway analysis of transcriptomic data indicates RTK signaling as the most enriched pathway in GBM consisting of overexpression of EGFR, PDGFR, Met, and others (Dong et al., 2010; Cancer Genome Atlas Research Network, 2008). The GnRH pathway may be involved in the regulation of EGFR and downstream MAP kinase cascades through GPCR-mediated transactivation of EGFR (Shah et al., 2003) or through deactivation by phosphotyrosine phosphatase (PTP) route (Grundker et al., 2001).
GnRH also has an important role in cell migration and is involved in transcriptional activation of several matrix metalloproteases—MMP2, MMP9, MMP14, and MMP26 (Ling Poon et al., 2011; Liu et al., 2010; Peng et al., 2016)—with three of them (MMP 2, MMP9, and MMP14) found to be upregulated in GBM datasets. Interestingly, expression of both MMP2 and MMP9 was also found to be required for EGFR transactivation by GnRH (Roelle et al., 2003). Given that GBM is a highly invasive phenotype, it may harbor distinct subpopulations of cells exhibiting invasive and proliferative types with their concurrent regulation at any point of time (Gao et al., 2005; Hecht et al., 2015).
Proteomic analysis of GBM from our laboratory has shown calcium (Ca2+) homeostasis and Ca2+ signaling to be major processes altered in GBM (Polisetty et al., 2012). Several calcium regulatory proteins and sensors such as S100 proteins and annexins that sequester Ca2+ are also found to be significantly upregulated in GBM and act as both intracellular regulators and extracellular signaling molecules. Deregulation of cytosolic Ca2+ levels is known to trigger proliferation or apoptosis of cancer cells and changes to cell shape and motility (Mermelstein et al., 2003; Roderick and Cook, 2008). Cytosolic Ca2+ signals and many Ca2+-permeable ion channels are also needed for optimal cell migration (Wondergem et al., 2008). Calcium is also known to activate PKC and downstream calmodulin-dependent (CAM) kinase pathways whose active state levels are again controlled by Ca2+ (Benard et al., 2001; Kraus et al., 2001). We observe a downregulation of CAMKs and MAPKs in GBM datasets, all probably implying a downregulation of their targets (HDACs and NFATs) leading to reduced apoptosis.
GnRH signaling is implicated in the regulation of Ca2+ levels through activation of PLC (two of the phospholipases, PLCE1 and PLCG1, are upregulated in GBM datasets), which catalyzes IP3 production that in turn activates its cognate receptor in the ER, leading to the release of intracellular stores of calcium (Ca2+) to cytosol. ITPR2, an IP3 receptor on the ER, is upregulated in GBM. Furthermore, the release of arachidonic acid under GnRH stimulation and consequent increased prostaglandin synthesis may be linked to deregulation of calcium homeostasis (Kiesel et al., 1986; Naidich et al., 2010; Polisetty et al., 2012).
Wnt signaling is a well-recognized pathway involved in cellular growth, differentiation, cell adhesion, and also induces motility and migration through upregulation of MMP expression. Wnt receptor, Frizzled (FZD3), and its paralogs (FZD2, FZD5, FZD6, FZD7), as well as several β-catenins (CTNNA1, CTNND1), the major mediators of Wnt signaling, are found to be upregulated in GBM dataset, suggesting activation of Wnt signaling. Recently, key intermediaries of Wnt signaling have been added to the diverse repertoire of GnRH signaling targets (Gardner et al., 2007, 2010; Gardner and Pawson, 2009; Salisbury et al., 2007, 2008). Upregulation of wnt pathway entities, particularly β-catenin in GBM, may suggest its contribution to both proliferation and motility presumably associated with different pathway modes (Nager et al., 2012) operating in invasive and proliferative subpopulation of cells present during tumor progression (Gao et al., 2005; Hecht et al., 2015). GnRH treatment was found to inhibit glycogen synthase kinase 3 (GSK-3B) and regulate degradation of β-catenin, resulting in its nuclear accumulation and consequent activation of its target genes that include several MMPs (Goto et al., 1999; Ling Poon et al., 2011).
Actin remodeling in GBM resulting from upregulation of α, β integrins, and MMPs has been documented (Musumeci et al., 2015; Schittenhelm et al., 2016). Actin remodeling and changes in extracellular matrix (ECM) are typically triggered by integrin receptors and several of them are (ITGB1, ITGB2, ITGB3, ITGB8, ITGA3, ITGA5, ITGA6, ITGAM, and ITGAV) found to be upregulated in GBM and known to alter cellular behavior through recruitment of non-RTKs (FAK; PTK2) and Src. FAK signaling downstream of integrin receptor engagement is reported to be central to the predominant mesenchymal mode of migration in GBM (Zhong et al., 2010). GnRH-activated Src kinases can directly phosphorylate FAK, FAK that is recruited to focal adhesion complexes on the plasma membrane by integrin heterodimers following integrin engagement of ECM proteins, and act on their substrates—actin cytoskeletal proteins (Paxillin; PXN, Actinin; ACTN1/4, Zyxin; ZYX, Vinculin; VCL, Filamins; FLNA, FLNC), resulting in changes in cell shape and promoting cell motility ultimately leading to tumor invasion and progression (Davidson et al., 2004; Dobkin-Bekman et al., 2009; Park et al., 2009). Several of these are all upregulated at both protein and transcript levels in GBM. These factors also seem to interact with GnRH receptor (Dobkin-Bekman et al., 2009; Park et al., 2009), implying the role of GnRH signaling in cell migration and invasion through cytoskeletal reorganization and increased matrix adherence (Davidson et al., 2004).
GnRH signaling as a possible therapeutic target in cancer has already been suggested and put together our observations strongly support this possibility in the context of GBM. GnRH I and GnRH II analogs—both agonists and antagonist—have been extensively explored in vitro and in vivo (Dondi et al., 1998; Fister et al., 2007, 2009; Grundker et al., 2001; Moretti et al., 2002; Rick and Schally, 2014; Sharma et al., 2005; van Groeninghen et al., 1998; Vincze et al., 1991). Interestingly, treatment of GBM cell lines by either GnRH analog (agonists or antagonists) resulted in the reduction of gonadotropin secretion and significant decrease in cell proliferation (Montagnani Marelli et al., 2009) yielding antiproliferative effects, although possibly by different routes.
Antagonists were found to block GnRH and promote proapoptotic effects through activation of P38 MAPK, JNK, and Bax, ultimately leading to tumor growth arrest (Fister et al., 2007, 2009; Grundker et al., 2001). On the other hand, sustained stimulation by agonists caused desensitization of the receptor-mediating antiproliferative effects through activation of PTPs that inhibit mitogenic signal of growth factors (such as EGFR). Expression of GnRH and GnRH receptor has been reported in GBM cell lines and tissue samples at both mRNA and protein levels and treatment with agonists resulted in antiproliferative activity (Limonta et al., 2003; Montagnani Marelli et al., 2009; Navarro et al., 2003). There is also evidence that the analogs can cross the blood–brain barrier, indicating suitability for treatment of malignant glioblastomas (Jaeger et al., 2005). Given this, further understanding of the molecular interplay involving GnRH signaling pathways would be important to identify potential target molecules for therapy.
Conclusions and Expert Outlook
Pathway mapping of the omics data provides a sequential flow of signaling information and places the omics data in a biological context providing systems-level insights. In this study, we used an approach to identify deregulated pathways by selecting specific protein families such as kinases from the multi-omics integrated datasets of GBM that were mapped to pathways, which revealed the GnRH signaling pathway to be significantly altered. With this lead, a complete GnRH pathway map was curated and GBM data were remapped to the curated pathway map, which resulted in its further enrichment with additional pathway mediators observed in GBM omics datasets. We also found possible cross talk of GnRH signaling with EGFR, calcium, Wnt, and FAK signaling involved in cell proliferation, invasion, and adhesion in GBM. Taken together, the results raise the potential of GnRH signaling as a target for further study.
Additionally, this study demonstrates the promise of multi-omics research and analyses to better understand complex cancers such as GBM. The regulatory molecular targets in the examined pathways have the potential to be exploited in GnRH receptor-positive GBMs as putative diagnostic and therapeutic candidates in the future. We suggest continued efforts and research investments on the multi-omics basis of common complex cancers.
Footnotes
Acknowledgments
The work reported here was funded under the project (BIC/12(9)/2012 Dated: 11/02/2013) from the Indian Council of Medical Research (ICMR) and Government of India to Ravi Sirdeshmukh. S.J. is a senior research fellow in this project and is registered as a PhD student at the Manipal University, Manipal. The authors gratefully acknowledge Pranali Sonpatki from Mazumdar Shaw Center for Translational Research and Pavithra Rajagopalan from Institute of Bioinformatics for help in making figures.
Author Disclosure Statement
The authors declare no conflicting financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.