
Abstract
Abstract
Networked 21st century society, globalization, and communications technologies are paralleled by the rise of electromagnetic energy intensity in our environments and the growing pressure of the environtome on human biology and health. The latter is the entire complement of environmental factors, including the electromagnetic energy and the technologies that generate them, enacting on the digital citizen in the new century. Electromagnetic pulse (EMP) irradiation might have serious damaging effects not only on electronic equipment but also in the whole organism and reproductive health, through nonthermal effects and oxidative stress. We sought to determine whether EMP exposure (1) induces biological damage on reproductive health and (2) the extent to which selenium-rich Cordyceps fungi (daily coadministration) offer protection on the testicles and spermatozoa. In a preclinical randomized study, 3-week-old male BALB/c mice were repeatedly exposed to EMP (peak intensity 200 kV/m, pulse edge 3.5 ns, pulse width 15 ns, 0.1 Hz, and 400 pulses/day) 5 days per week for four consecutive weeks, with or without coadministration of daily selenium-rich Cordyceps fungi (100 mg/kg). Testicular index and spermatozoa formation were measured at baseline and 1, 7, 14, 28, and 60 day time points after EMP exposure. The group without Cordyceps cotreatment displayed decreased spermatozoa formation, shrunk seminiferous tubule diameters, and diminished antioxidative capacity at 28 and 60 days after exposure (p < 0.05). The Cordyceps daily cotreatment alleviated the testicular damage by EMP exposure, increased spermatozoa formation, and reduced apoptotic spermatogenic cells. These observations warrant further preclinical and clinical studies as an innovative approach for potential protection against electromagnetic radiation in the current age of networked society and digital citizenship.
Introduction
N
A large number of epidemiological and experimental studies have suggested that exposure to electromagnetic radiation could result in serious reproductive health issues, including decreased sperm counts, infertility, sexual dysfunction, abnormal embryonic development, congenital deformity, perinatal death, and abnormal development in utero (Agarwal et al., 2009; Kesari et al., 2010; Shahin et al., 2013). Several reports have also shown that certain parameters of electromagnetic radiation do not elicit positive biological effects (An et al., 2015; Dasdag et al., 2003; Lee et al., 2010). Our department has confirmed that EMR in certain parameters can cause learning and memory decline, reproductive hormone disorders, increased permeability of the blood–testis barrier in testicular tissue, decreased activity of antioxidant enzymes, increased antisperm antibodies and free radical product (i.e., malondialdehyde [MDA]) levels, and declined acrosome enzymatic reactions (Chen et al., 2011; Hou et al., 2012; Lin et al., 2015; Wang et al., 2008; Zeng et al., 2011).
As a consequence of advances in technology, the total amount of electromagnetic radiation exposure received by juveniles has increased dramatically in recent decades. The juvenile stage represents a critical maturation period for different body systems, particularly the male reproductive system. This system is highly sensitive to a variety of external stimuli. Studies have shown that the testis is sensitive to electromagnetic radiation. Until now, the effects of electromagnetic radiation on premature animals have rarely been reported (Vergouwen et al., 1995). Hence, this study evaluated the impact of EMP irradiation on the testicles and spermatozoa of young male mice.
Selenium is a multifunctional nutrient of great importance in China. Organic selenium is used in both humans and animals and has been shown to eliminate free radicals, facilitate the provision of antioxidative effects, strongly inhibit the generation of lipid peroxidation, enhance immune function by 20- to 30-fold, and protect cell membranes. Selenium-rich Cordyceps is an entomopathogenic fungus that has been bred to contain a high concentration of organic selenium. In addition, this fungus contains relatively higher concentrations of cordycepin, Cordyceps acid, and Cordyceps polysaccharides than Chinese Cordyceps species from the Qinghai–Tibet region. Selenium-rich Cordyceps also exhibits higher superoxide dismutase (SOD) activity (than Chinese Cordyceps species) and has been shown to display relatively high efficiencies of antioxidant scavenging of free radicals, with concomitant anticancer effects and immune system enhancement (Li et al., 2012; Nakamura et al., 2015; Wang et al., 2011, 2014; Zhong et al., 2008). Thus, to further elucidate the potential protective effects associated with selenium-rich Cordyceps, we investigated the protection of selenium-rich Cordyceps in the reproductive system following EMP exposure.
Materials and Methods
Ethics statement
All procedures for experiments performed on mice were reviewed and approved by the Animal Ethics Committee at Fourth Military Medical University (Approval ID FMMU 2007102020).
Animals and study groups
A total of 120 male BALB/c mice (3 weeks old, weighing 9–11 g) were received from the Laboratory Animal Center of the Fourth Military Medical University, Xi'an, Shaanxi, China [Certification No.: scxk (Military) 2007-007]. These mice were randomly divided into three groups of 40, including a sham exposure control group (given sham irradiation and without selenium-rich Cordyceps treatment), an exposure group without selenium-rich Cordyceps treatment, and a selenium-rich Cordyceps-treated irradiation group. Each group of animals was further subdivided into the following five time points; 1 day (d), 7, 14, 28, and 60 days. Each subgroup encompassed eight mice.
Animal positioning and EMP radiation
The all-solid-state nanosecond generator that was utilized in this study was developed and tested in the Northwest Institute of Nuclear Technology in Xi'an, China (Ding et al., 2009). Selenium-rich Cordyceps powder (procured from Anhui Anshan Huayu Biology & Technology Co.) was dissolved in double-distilled water to a final concentration of 20 g/L and stored at −4°C. The dissolved selenium-rich Cordyceps powder mixed with double-distilled water was intragastrically administered on a daily basis (100 mg/kg) to mice in the selenium-rich Cordyceps-treated irradiation group before exposure each day. Mice that were exposed to this treatment regimen were housed together with mice from the other group. Four mice were kept in a 16 × 5.5 × 4 cm aperture plastic box. Both groups of animals were irradiated at a field intensity of 200 kV/m EMP, with a pulse edge of 3.5 ns, pulse width of 15 ns, repetition frequency of 0.1 Hz, and pulse frequency of 400 times/day. Mice in exposure groups were irradiated 5 days per week for four consecutive weeks. Irradiation was performed between 15:00 and 17:00 at room temperatures ranging from 22 to 24°C. Mice in the sham control group were given sham irradiation. Four sham exposure control mice were placed in the organic plastic box and left inside the EMP generator for the same duration as the two experimental groups. The sham exposure mice were subjected to similar sound stimuli (as the irradiated mice) following exposure to the same pulse frequency. Animals from each group were weighed every 4 days.
Determination of testicular index
Mice were weighed before being euthanized at the aforementioned time points. After decapitation, both testicles, from each animal, were dissected, weighed, and the testicular index was calculated (i.e., testicular index = mouse testicle weight (g)/mouse body weight (g) × 100%).
Measurement of seminiferous tubule diameter and detection of apoptosis
One dissected testicle, from each mouse, was stored at −80°C.The other testicle was fixed in Bouin's solution for 24 h and dehydrated using a graded ethanol series. The specimen was subsequently cleared with xylenes and embedded in paraffin. Coronal sections of 4 μm thickness were then obtained.
After routine hematoxylin and eosin (HE) staining, tissue sections were observed using light microscopy. The diameters of seminiferous tubules across the minor and major axes were measured for five mice at each time point that was assessed. A total of 40 seminiferous tubules were measured per mouse.
Spermatogenic cell apoptosis was detected using terminal deoxynucleotidyl transferase-mediated nick end labeling (TUNEL) fluorescent staining, according to the manufacturer's protocols. Routine dewaxing and antigen retrieval steps were subsequently performed. Digital images were taken from each stained tissue section. The percentage of TUNEL-positive cells per unit area was calculated using Leica analytical software.
Mouse sperm count and deformity rate analysis
At various time points, after the end of irradiation treatments, each animal was euthanized by decapitation and bilateral epididymis was collected. The collected tissue was shredded in 1 mL of preheated normal saline (preheated to 37°C). After a freestanding incubation step at 37°C for 20 min, the supernatant was collected and filtered through a 400 mesh sieve. A total of 20 μL of filtrate was added to 380 μL of sperm diluent (i.e., 5 g sodium bicarbonate, 1 mL formaldehyde solution, and distilled water made up to a final volume of 100 mL) to count spermatozoa using a hemocytometer. Each sample was counted four times. The smear filtrate was collected and air-dried. The filtrate was subsequently fixed in methanol for 30 min and stained in 2% eosin for 2 h. The number of head deformities from 200 integrated spermatozoa was measured from each mouse under high magnification (40 × 10), using five mice per group.
Content detection of antioxidant enzymes and lipid peroxide
The testicular weight of each mouse was accurately measured. Associated tissue samples were then shredded and homogenized while on ice. Next, the sample was centrifuged at 4000 rpm for 10 min at 4°C, and finally the supernatant was collected and stored for later measurements. Total protein concentration was determined using a Bio-Rad protein quantification kit. The total antioxidant capacity (T-AOC), SOD, MDA, and glutathione (GSH) activities or concentrations were measured using the corresponding detection kits, purchased from Nanjing Jiancheng Biological Engineering Co., Ltd. (Nanjing, China), and the manufacturers' protocols were strictly followed.
Statistical analysis
All data are expressed as mean ± SD. Data analysis was performed using the SPSS 17.0 software. Results were analyzed by repeated measures analysis of variance (ANOVA) and with post hoc paired comparisons if and when the general ANOVA model was significant.
Results
Effects of selenium-rich Cordyceps treatment on the mouse testicular index of mice exposed to EMP
Compared with the sham exposure control group, the body weights of mice in the irradiation group without selenium-rich Cordyceps treatment gradually decreased after irradiation and their testicle weights, at various time points after irradiation, were significantly reduced (Fig. 1A, B). In addition, the testicular index values for the irradiation group without selenium-rich Cordyceps were significantly lower at 28 and 60 days after irradiation (p < 0.05) compared with the sham exposure group. Upon comparison of the two exposure groups, the testicular indices of mice from the group that received intragastric selenium-rich Cordyceps treatment were significantly increased at 28 and 60 days after irradiation versus the irradiation group without selenium-rich Cordyceps (p < 0.05, Fig. 1C).
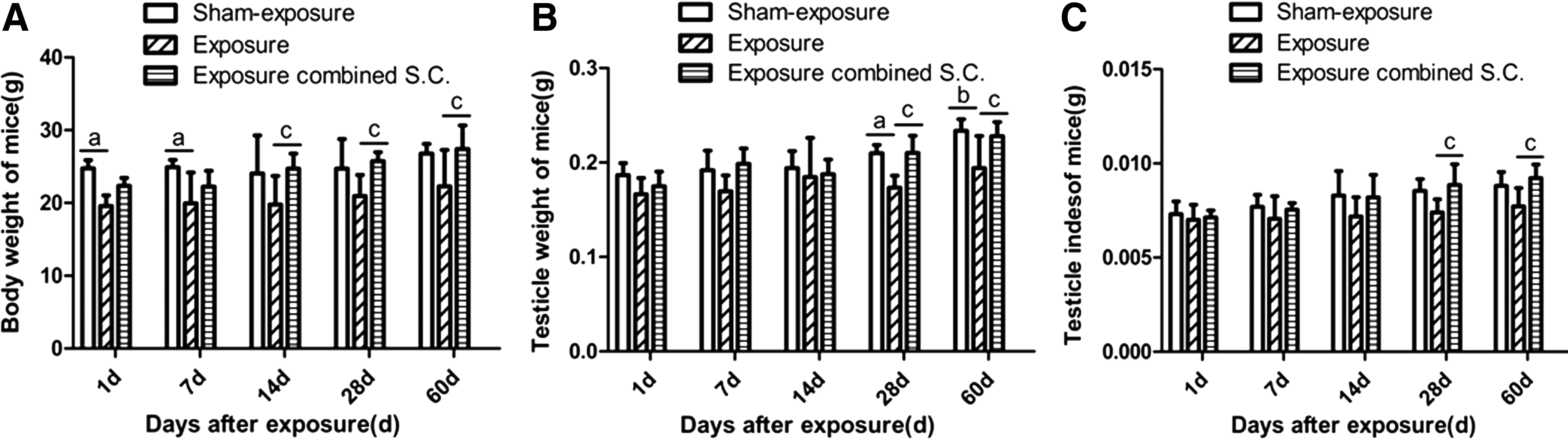
Effects of selenium-rich Cordyceps treatment on body weight, testis weight, and testis index of mice exposed to EMP. Mice were exposed to EMP 200 kV/m, 200 pulses/day, and 5 days per week for 4 weeks.
Effects of selenium-rich Cordyceps treatment on the diameter of seminiferous tubules and spermatogenic cell apoptosis in mice exposed to EMP
Microscopic observations (using the 20 × objective lens) of HE-stained specimens were used to determine the diameter of seminiferous tubules across the minor and major axes. The representative images are shown in Figure 2A.

Effects of selenium-rich Cordyceps treatment on the diameter of seminiferous tubules in mice exposed to EMP.
Figure 2B shows that the diameters of seminiferous tubules of mice from the irradiation group without selenium-rich Cordyceps treatment at 1, 7, 14, 28, and 60 days were significantly decreased compared with the sham exposure control group (p < 0.05). Upon comparison of the two irradiation groups, the diameters of seminiferous tubules from mice in the selenium-rich Cordyceps-treated irradiation group were significantly increased at 28 and 60 days after irradiation compared with those from the mice in the exposure group without selenium-rich Cordyceps treatment (p < 0.05).
TUNEL staining of testicular tissues is depicted in Figure 3. The green fluorescence in the TUNEL stains represents apoptotic cells. These cells were mainly distributed in the seminiferous tubules (Fig. 3A). The percentages of apoptotic spermatogenic cells in mice from the irradiation group without selenium-rich Cordyceps treatment were significantly higher than those from the sham exposure control group at 1 and 60 days after EMP radiation. However, the percentages of apoptotic spermatogenic cells in mice were lower than those in the sham exposure control group at 7 and 14 days after irradiation (p < 0.05, Fig. 3B). The percentage of apoptotic spermatogenic cells in the selenium-rich Cordyceps-treated irradiation group was significantly reduced at all time points after irradiation compared with the sham exposure control group (p < 0.05).The percentage of apoptotic spermatogenic cells in the selenium-rich Cordyceps-treated irradiation group was significantly lower than in the irradiation group without selenium-rich Cordyceps treatment at 14, 28, and 60 days after irradiation (p < 0.05).
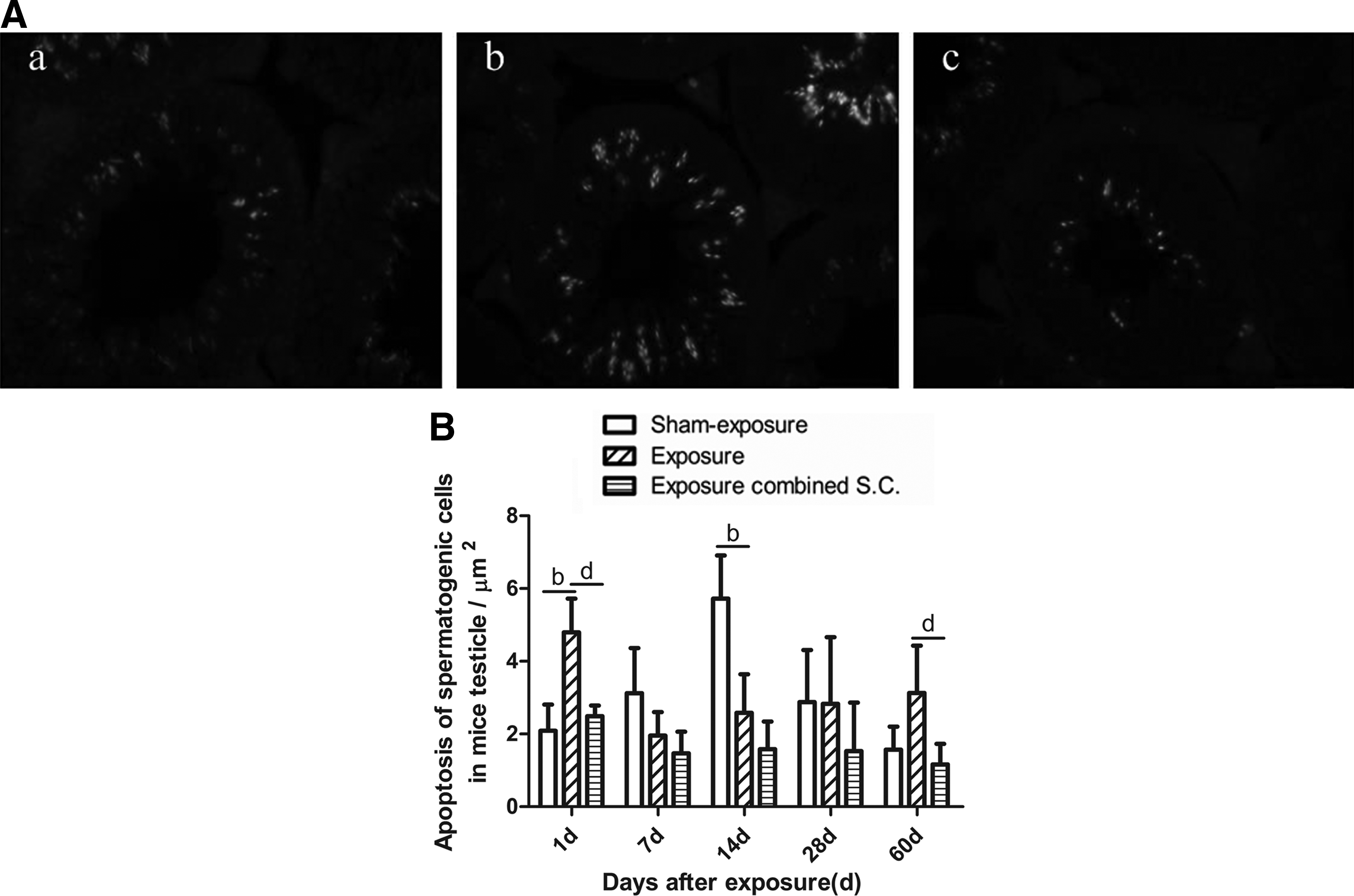
Effects of selenium-rich Cordyceps treatment on spermatogenic cell apoptosis in the testicles of mice exposed to EMP.
Effects of selenium-rich Cordyceps treatment on spermatozoa number and percentage of spermatogenic deformities in mice exposed to EMP
Upon comparison with the sham exposure control group, the spermatozoa numbers in the irradiation group without selenium-rich Cordyceps treatment were significantly reduced at 7, 14, 28, and 60 days after EMP radiation (Fig. 4A). The selenium-rich Cordyceps-treated irradiation group had a significantly lower number of spermatozoa than the sham exposure control group at 7 and 14 days after EMP radiation (p < 0.05). The spermatozoa numbers in the selenium-rich Cordyceps-treated irradiation group were higher at all time points compared with the irradiation group without selenium-rich Cordyceps treatment; there were significant differences at 28 and 60 days after exposure (p < 0.05).
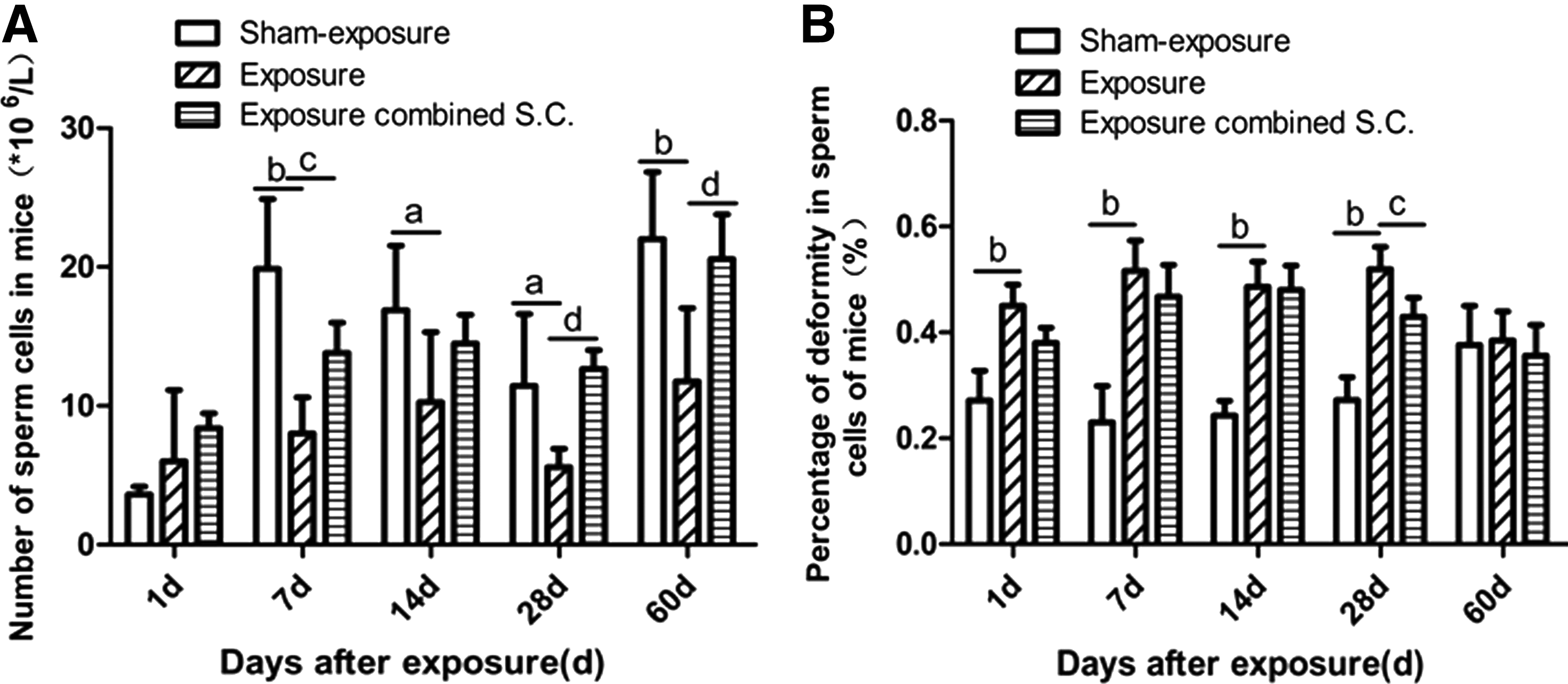
Effects of selenium-rich Cordyceps treatment on sperm number and deformity percentage of mice exposed to EMP.
Upon comparison with the sham exposure control group, the spermatogenic deformity rates for the irradiation group without selenium-rich Cordyceps treatment were significantly higher at 1, 7, 14, and 28 days after irradiation (p < 0.05, Fig. 4B). No significant difference was observed at 60 days after irradiation. Although the spermatogenic deformity rates of the selenium-rich Cordyceps-treated irradiation group were lower compared with the irradiation group without selenium-rich Cordyceps treatment at various time points, the observed differences were not significant.
Effects of selenium-rich Cordyceps treatment on the activity of antioxidant enzyme and MDA levels in the testicles of mice exposed to EMP
Animals in the exposure group without selenium-rich Cordyceps treatment had significantly lower T-AOC activity in the testicles, at various time points after irradiation, compared with the sham exposure control group (Fig. 5A, p < 0.05). The testicular T-AOC activity in mice from the selenium-rich Cordyceps-treated irradiation group was significantly higher compared with the irradiation group without selenium-rich Cordyceps treatment (p < 0.05).

Effects of selenium-rich Cordyceps treatment on the activity of antioxidant enzyme and MDA levels in the testicles of mice exposed to EMP.
The SOD activity in mice from the exposure group without selenium-rich Cordyceps treatment was significantly lower than for the sham exposure control group at 1, 7, and 14 days after irradiation (Fig. 5B). There were no significant differences between these two groups at the other time points that were assessed. The SOD activity in the selenium-rich Cordyceps-treated irradiation group was significantly higher compared with the irradiation group without selenium-rich Cordyceps treatment at various time points after irradiation (p < 0.05).
The postirradiation GSH activity in the testicles of mice from the irradiation group without selenium-rich Cordyceps treatment was significantly lower than the sham exposure control group (p < 0.05 at 1 and 60 days after radiation; p < 0.01 at 7, 14, and 30 days after radiation, Fig. 5C). The postirradiation GSH activity in the testicles of mice from the selenium-rich Cordyceps-treated irradiation group was significantly higher than that in the irradiation group without selenium-rich Cordyceps treatment (p < 0.05).
Finally, MDA levels in testicles from mice in the irradiation group without selenium-rich Cordyceps treatment were significantly higher at various time points after irradiation (p < 0.01) compared with the sham exposure control group (Fig. 5D). The MDA levels in testicles of mice from the selenium-rich Cordyceps-treated irradiation group showed no significant difference compared with the sham exposure control group. The MDA levels in testicles of mice in the selenium-rich Cordyceps-treated irradiation group were significantly lower compared with the irradiation without selenium-rich Cordyceps treatment (p < 0.05).
Discussion
As the electromagnetic radiation load is rapidly increasing in our globalized societies and communication age, there is a need to understand the putative biological effects of such continued environmental exposures. In the present study, we aimed to develop new insights to address this research question.
Three-week-old male BALB/c mice were utilized to evaluate the effects of EMP radiation and selenium-rich Cordyceps on the testicles and spermatozoa of young male mice. Our results showed that irradiation using a 200 kV/m field intensity and 400 times EMP for four consecutive weeks significantly reduced testicular weight in young male mice. Lower testicular weights were observed throughout the study and these values in radiation group were significantly lower than the weights observed in mice from the sham exposure control group. Moreover, the testicular indices and the diameters of seminiferous tubules in mice from the irradiation group without selenium-rich Cordyceps treatment, at 60 days after irradiation, were significantly lower and smaller, respectively, than those observed in the control group. These observations indicate that EMP radiation exposure can suppress growth in the reproductive organs of young male mice. The large cumulative dose of EMP radiation that was utilized in this study resulted in irreversible damage in tissue structures, thereby affecting function. Testicles predominantly comprise Leydig cells and the epithelium of seminiferous tubules. The seminiferous tubules are responsible for spermatozoa formation. In this study, a reduction in testicular weight, testicular index values, and the diameter of seminiferous tubules was observed following EMP irradiation. These observations suggest that EMP irradiation affects the quality, production, and development of spermatozoa.
Results from the present study showed that irradiation conditions may reduce testicular weight, testicular index values, and the diameter of seminiferous tubules in young male BALB/c mice. For example, compared with the sham exposure control group, the number of spermatozoa in mice from the irradiation group without selenium-rich Cordyceps treatment was significantly reduced at 7, 14, 30, and 60 days after irradiation. Furthermore, the deformity rates of spermatozoa in this group were significantly increased at 1 and 30 days after irradiation compared with the sham exposure control group. Results from TUNEL staining revealed that the percentage of apoptotic spermatogenic cells in mice from the irradiation group without selenium-rich Cordyceps treatment was significantly enhanced at 1 and 60 days after irradiation compared with the sham exposure control group. The percentage of apoptotic spermatogenic cells in mice from the latter group was significantly lower than those found in the sham control group at 7 and 14 days postirradiation.
The TUNEL staining method that was utilized to assess apoptosis has some technique-specific limitations and can result in the false-positive staining of tissue that is undergoing excessive cell division. TUNEL staining results in the labeling of DNA fragment termini and a large number of termini belonging to complete DNA molecules are exposed during DNA synthesis and DNA replication in mitotic cells. As a result, these termini can be inadvertently labeled as apoptotic cells. The significantly higher number of apoptotic spermatogenic cells in mice from the sham exposure control group at 7 days (8.5 weeks of age) and 14 days (9.5 weeks of age) is most likely due to spermatogenic cell maturation following the previous spermatogenic cycle (this occurs at ∼5 weeks of age in mice). In addition, the initiation of mitosis before the next spermatogenic cycle might result in increased false-positive cells in the sham control group.
Spermatogenesis is a sophisticated example of a continuously synchronized and spatially ordered differentiation step (Stern et al., 1983). Due to the synchronized timing of stem cell differentiation into spermatozoa, transverse sections of seminiferous tubules will always exhibit typical cellular grouping patterns from the basement membrane to the tubular lumen. These patterns reflect the continuous growth of spermatozoa along the seminiferous tubules, a phenomenon that can trigger periodic variations in spermatozoa formation. Our results clearly reflect these variations in the seminiferous cycle. Mice exposed to EMP experienced a 1- to 2-week delay in the spermatogenesis cycle compared with the sham exposure control group. Moreover, these analyses indicate that the number of spermatozoa produced during spermatogenesis is only 25% to 27% of theoretical production output. Spermatogenic cell apoptosis has been widely reported in normal spermatozoa formation. As a result of apoptosis, a balanced number of spermatozoa are maintained in the seminiferous tubules of in vivo models. This process ensures the normal development and maturation of spermatozoa while also facilitating the maintenance of an appropriate number of mature spermatozoa. Furthermore, apoptosis is necessary to ensure normal physiological function in testicles. A study by Hanci et al. reported that 900-MHz electromagnetic field (EMF) applied during the prenatal period resulted in damage to seminiferous tubules in rat pup testes. Hanci et al. (2013) also observed a reduction in both seminiferous tubule diameter and epithelial thickness. Ozlem et al. exposed postnatal day 2 male albino Wistar rats to an EMF of 1.8 or 0.9 GHz following cell phone exposure. The rats were exposed to the EMF for 2 h per day for 90 consecutive days. This study reported that animals that were exposed to 1.8 GHz EMF had severe vacuolization and apoptosis in their seminiferous epithelium (Ozlem et al., 2012).
Studies have shown that multiple electromagnetic radiation exposures can directly or indirectly induce oxidative stress in organisms, through nonthermal effects that cause secondary damage effects. These nonthermal effects are considered to be the main mechanisms by which electromagnetic radiation produces adverse biological effects (Nakamura et al., 2003; Schastnyĭ et al., 1996; Turkozer et al., 2008). During and shortly after exposure to electromagnetic radiation, oxygen free radical levels are significantly increased. In addition, the expression of a variety of antioxidant enzymes is upregulated to protect the body against peroxidation injury (Al-Damegh, 2012; Ozturk et al., 2003). It has been reported that several antioxidant drugs can play a protective role against electromagnetic radiation. Chen et al. (2011) utilized a field intensity of 400 kV/m and 200 times EMP to irradiate mice. Their results showed that a variety of antioxidant enzymes in brain tissues were reduced, while levels of the peroxidation injury marker, MDA, were increased, resulting in learning and memory deficits in these mice. In addition, in this study, TMG was shown to play a protective role against EMP damage (Chen et al., 2011).
Zeng et al. (2011) reported that exposure to EMP pulse irradiation with a field intensity of 100 kV/m and a frequency of 4 × 105 pulses resulted in fluctuations in serum hormone levels, a reduction in SOD activity in testicular tissues, and significantly increased levels of MDA within a short period of time. Xu et al. reported that 1.8 GHz of radio frequency electromagnetic irradiation induced increased γH2AX formation in Chinese hamster ovary (CHO) cells and HSF cells. These results indicated the activation of oxidative stress (Xu et al., 2013). In this study, the T-AOC, SOD, and GSH activities or concentrations in the testicular tissue of mice from the irradiation group without selenium-rich Cordyceps treatment were significantly decreased after irradiation. Testicular MDA levels in mice from the irradiation group without selenium-rich Cordyceps treatment were significantly increased after irradiation. Our results suggest that prolonged exposure to EMP radiation affected the balance in the antioxidant enzymatic system in testicular tissues, resulting in a decrease in T-AOC, SOD, and GSH activities. EMP radiation led to inefficient and delayed removal of excessive free radicals in the body, thereby significantly increasing the content of the MDA metabolite.
The selenium-rich Cordyceps used in this study contains concentrations of total selenium and organic selenium that are higher than ordinary Cordyceps by ∼70 times and 100 times, respectively (Ji et al., 2014; Lang et al., 2012). In this study, the organic selenium content accounted for 97% of the total selenium concentration. The selenium polysaccharide in selenium-rich Cordyceps has a scavenging effect on hydroxyl radicals. This polysaccharide can reduce or prevent the occurrence of lipid peroxidation, thereby playing a protective role in the cell membrane. Selenium-rich Cordyceps is rich in SOD, an enzyme that inhibits free radical oxidation (Dong et al., 2012; Zheng et al., 2014). Our study also confirmed that intragastric treatment of selenium-rich Cordyceps increased the activity of T-AOC, SOD, and GSH. Selenium-rich Cordyceps treatment also increased free radical scavenging capacity following irradiation. Therefore, the MDA content in testicular tissues from mice in the selenium-rich Cordyceps-treated irradiation group was significantly reduced. In this study, selenium-rich Cordyceps also alleviated testicular damage caused by EMP irradiation exposure by increasing the testicular index, reducing the number of apoptotic spermatogenic cells, and increasing spermatozoa formation in male mice. This analysis reveals new strategies that can be harnessed to assess the biological effects of EMP on reproductive systems.
Finally, it is important to point out that there are limitations associated with this study. First, although the Cordyceps strain that was used in this study is known to produce increased amounts of total and organic selenium, we cannot conclude that selenium production was the catalyst for the observed biological effects. Second, the analysis did not investigate how variations in EMP intensity and exposure might differentially affect reproductive systems. Third, we only analyzed a subset of parameters that might be deemed important in assessing reproductive capacities.
Conclusions and Future Outlook
This preclinical study reveals new methods that can be used to evaluate the biological effects of electromagnetic radiation on reproductive health. We observed that selenium-rich Cordyceps can play a significant role in protection against EMP irradiation-induced spermatic damage. Furthermore, this report is the first to elucidate the potential of selenium-rich Cordyceps in protecting reproductive systems against EMP exposure. Our results provide an innovative approach that facilitates the study of protection in relation to electromagnetic radiation-induced injuries.
Future studies should be conducted to investigate the effect of selenium-rich Cordyceps on testosterone production and sperm mobility following EMP exposure. In all, these observations warrant further preclinical and clinical studies as an innovative approach for potential protection against electromagnetic radiation in the current age of networked society and digital citizenship.
Footnotes
Acknowledgments
This work was financially supported by grants from the Ministry of Science and Technology (2011CB503704), the National Natural Science Foundation of China (Nos. 51677189, 31470829, and 81272490), the International Science & Technology Cooperation Program of China (No. 2010DFA31900), and the National Natural Science Foundation of Shaanxi (No. 2015JM8454).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.