
Abstract
Abstract
Genome instability has long been implicated as a salient causal factor in aging and age-related diseases such as cancer and neurodegeneration. However, the molecular mechanisms associated with genome instability remain unclear. Recent studies suggest growth signaling in the organism Saccharomyces cerevisiae and in higher eukaryotes might affect oxidative stress and aging/age-related diseases by activating DNA replication stress that causes DNA damage. In this broader integrative biology and clinical context, MDM two binding protein (MTBP) is a binding protein that has a role in activation of the tumor suppressor protein p53. Clinical studies suggest that the increase in expression of MTBP leads to reduction in survival of breast cancer patients. The functional homolog of MTBP in yeast, Sld7, is a hitherto uncharacterized protein that decreases the affinity of Sld3 (Treslin/Ticrr in humans) toward Cdc45 in the cell cycle process of S. cerevisiae. To investigate the putative function of Sld7 in chronological aging and replicative life span, we applied a network biology approach, integrating interactome and transcriptome data of budding yeast. The cell cycle and chronological aging networks of proteins in budding yeast were reconstructed. Furthermore, through meta-analysis of cell cycle and chronological aging-associated transcriptome datasets, we constructed coexpression networks and identified coexpressed gene clusters. In this study, we propose a dual role for Sld7: it participates in macromolecular complex binding in the cell cycle and has oxidoreductase activity in chronological aging in budding yeast. These potential roles of Sld7 in yeast can offer new insights on the role of MTBP in humans and consequently might lead to novel strategies for treatment of cancers and aging-related diseases.
Introduction
G
Sld7 is a limited abundance protein that regulates DNA replication initiation by decreasing the association of Sld3 with Cdc45, both of which are essential proteins in the G1 phase of cell cycle (Araki, 2010; Tanaka et al., 2011b). The depletion of Sld7 slows down DNA replication and cell growth in budding yeast (Tanaka et al., 2011b). The crystal structures of Sld3 and Sld7 were determined by X-ray crystallography, and binding of Sld7 to N-terminal of two Sld3 molecules was noted earlier (Itou et al., 2015). The quaternary structure of Sld7-Sld3 complex is suitable for binding Cdc45 and GINS onto Mcm2-7 on a replication origin (Itou et al., 2015). Sld7 is localized in the nucleus and colocalized with spindle pole (Huh et al., 2003). Hence, it might have a role in aberrant mitochondrial morphology (Altmann and Westermann, 2005). Despite the comprehensive research, the molecular function of Sld7 is still unknown.
A functional relationship between yeast Sld7 and human MDM two binding protein (MTBP) has been proposed (Boos et al., 2013). MTBP is one of the binding proteins of Treslin/Ticrr, which is the homolog protein of Sld3 in yeast. The reduction of MTBP affects human cells in the same way as the depletion of Sld7 affects growth in yeast cells. The expression of MTBP was reportedly elevated in breast and ovarian cancers (Boos et al., 2013; Grieb et al., 2014). The data from The Cancer Genome Atlas (TCGA) confirmed that 19% of breast cancer patients showed amplification and overexpression of MTBP.
Clinical research indicated that the increase in the expression of MTBP led to reduction in the survival of breast cancer patients (Bassi et al., 2016; Chen and Butte, 2016; Grieb et al., 2014; Han et al., 2016). Thus, targeting MTBP might be a novel promising therapeutic approach in the treatment of cancer. Indeed, the research in mouse haploinsufficient in MTBP revealed a decrease in growth and development of lymphoma (Odvody et al., 2010). The potential of MTBP in humans or Sld7 in yeast as a drug target needs to be explored for therapeutic purposes.
In our previous study, the gene expression level of Sld7 was found to change significantly in chronological aging-related experiments (Yucel et al., 2014). In the present study, the putative molecular functions of Sld7 in chronological life span and replicative life span (RLS) were studied by computational methods using an integrated systems biology approach. We analyzed the physical interactions of Sld7 with other proteins in the reconstructed cell cycle and chronological aging networks of budding yeast. In addition, coexpression patterns were analyzed by combining transcriptome data from several experiments with the interactome networks presented here so as to propose possible roles of Sld7 protein in aging of budding yeast. The highly interacting and significantly coexpressed gene/protein clusters around Sld7 in these networks were further examined with functional enrichment analyses. The results indicated a dual role of Sld7 in both cell cycle and chronological aging processes of budding yeast.
Materials and Methods
Network reconstruction by Selective Permissibility Algorithm
The Selective Permissibility Algorithm (Arga et al., 2007), an efficient process in reconstruction of context-specific interaction networks around a core set of molecules, was adapted to reconstruct the signaling networks of cell cycle and chronological aging in yeast.
In brief, core proteins were determined by finding the proteins annotated with cell cycle term in the Gene Ontology Database for S. cerevisiae S288c (+) organism released on September 4, 2015. The gene ontology (GO) annotation terms (molecular function/biological process/cellular component) of these core proteins were used as inputs to the algorithm and they form the annotation collection table (ACT).
The candidate proteins that have a role in the cell cycle by interacting with these core protein(s) were included only if all GO annotation terms were present in the ACT. Proteins added through this procedure became the new input proteins and the algorithm expanded the network in a cyclic way until no new interacting proteins could be added to the network (Fig. 1). At last, the high-confidence physical interactome data (confidence score >0.5) of S. cerevisiae (Karagoz and Arga, 2013) were used to find the related interactions and to reconstruct the cell cycle network (CCN).

Schematic representation of the network reconstruction algorithm, SPA for CCN1. SPA, Selective Permissibility Algorithm.
The number of interactions was high in CCN. To reduce the noise in the interactome data, the unknown GO annotation terms (component/process/function) were eliminated from ACT. As the molecular function of Sld7 is unknown, it was added manually into the core proteins of the network. The constructed network with eliminated unknown function terms was called CCN2. In addition, to query any possible role of Sld7 in the initiation of DNA replication in G1 phase of budding yeast as proposed (Tanaka et al., 2011b), a network around proteins associated with G1 phase of yeast cell cycle (CCN3) was also reconstructed.
The same procedure was followed to form chronological aging networks, CAN and CAN2, where gene products sharing the chronological cell aging GO biological process term were extracted from the manually curated literature data of Saccharomyces Genome Database, released on February 17, 2016.
Cluster identification and functional enrichment
The reconstructed networks were represented as undirected graphs (i.e., nodes represent proteins and the edges represent interactions between the proteins), visualized, and analyzed using Cytoscape (v2.8.3) (Smoot et al., 2011). To determine highly interconnected protein clusters, topological analyses were applied through MCODE plug-in (Bader and Hogue, 2003) with default settings. The clusters that involve the protein of interest, that is, Sld7, or any of its interactors (Sld3, Nup116, Mth1, Nab2, Ssa2, Srp40, and Hek2) were examined using GO TermFinder (Boyle et al., 2004) as the functional enrichment analysis tool to identify over-represented GO terms associated with proteins in clusters. Hypergeometric test was employed to evaluate the significance of the enrichments: the ratio of cluster frequency over background frequency was calculated for each GO annotation term as a percentage value to assess the significance of an enriched function with respect to that of the whole yeast genome. This procedure was followed for each cluster.
Network formation from transcriptome data
To understand the genetic interactions of Sld7, the transcriptome data recorded by Yucel et al. (2014), in an environment with and without pH control for both young and old (aged) cells, were transformed into a network, where nodes represent proteins and edges represent absolute value of the correlations of proteins being equal or higher than 0.99 (Köberlin et al., 2015).
The overview of procedures followed in the present multiomics approach is given in Figure 2.
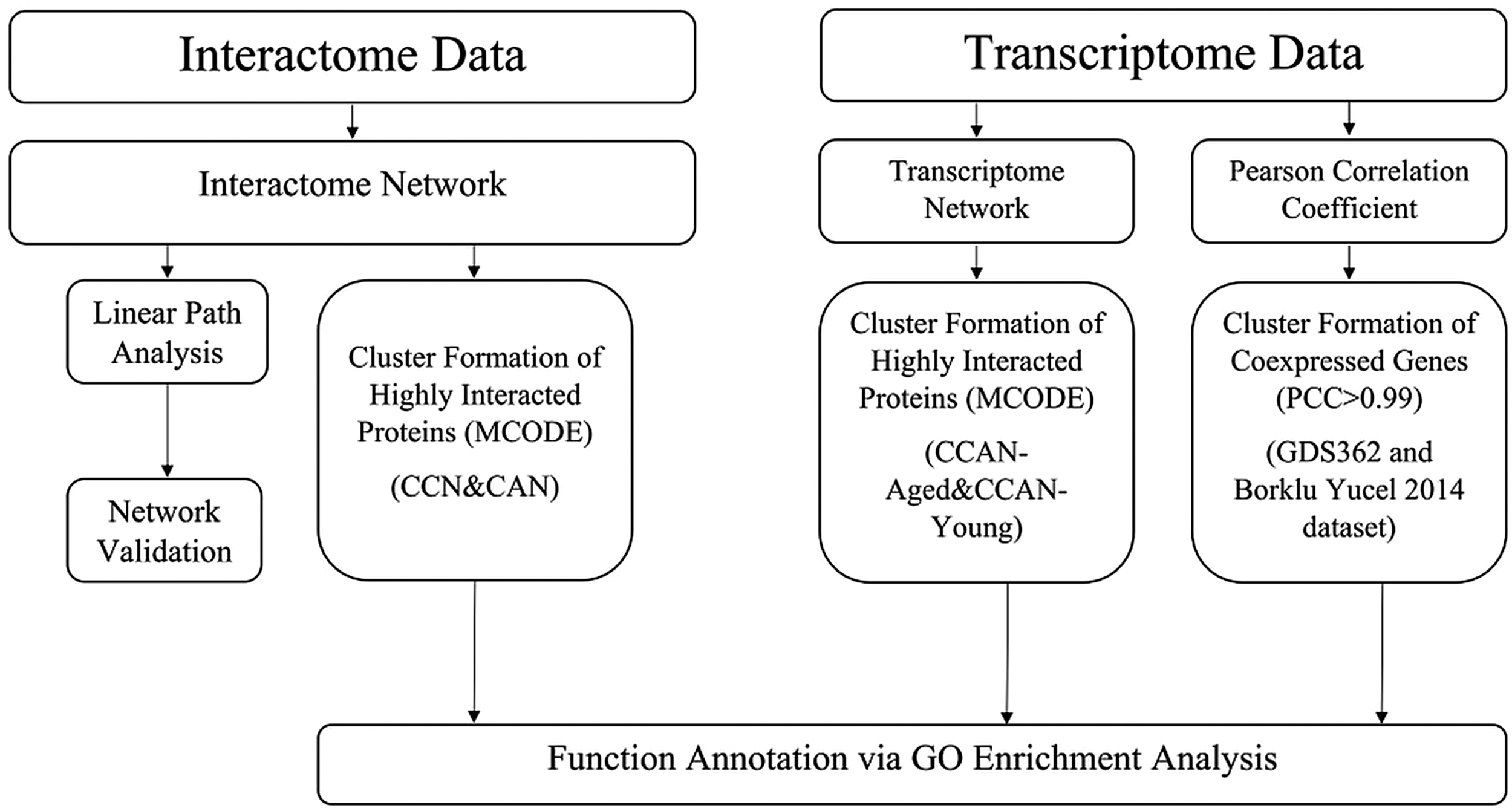
Overview of the computational methods employed to estimate the molecular function of Sld7.
Coexpression analysis
The cell cycle and aging-related transcriptome data were downloaded from the Gene Expression Omnibus (GEO) database. To measure the strength of a linear association between two variables (i.e., the coexpression pattern of genes), the Pearson correlation coefficient (PCC), denoted by r, is used [Eq. 1].
The PCCs of the genes were calculated with respect to Sld7. The coexpressed genes were determined based on the threshold value, which should not be lower than 0.7 for accuracy. The highly coexpressed genes (or their encoded products, proteins) were accepted as a cluster and the molecular function of this cluster was examined through GO enrichment analysis.
Linear path analysis
To understand information flow through the prereplicative complex (pre-RC) in S. cerevisiae, linear path analysis was carried out using the NetSearch algorithm (Steffen et al., 2002). The starting proteins were selected as origin recognition complex (ORC; Orc1, Orc2, Orc3, Orc4, Orc5, and Orc6), the initiator proteins bind to replication origins during the cell cycle process (Araki, 2011), and the proteins that have a role in pre-RC were selected as the target proteins (Cdt1, Cdc2, Cdc6, Cdc9, Cdc7, Cdc28, and Pol1) (Araki, 2010). Different path lengths were tested starting from two steps to seven steps, and the paths involving Sld7 were examined.
Results
The biological function of a protein is usually determined by its physical interactions with other proteins (Karagoz et al., 2016). The protein interactome (i.e., protein–protein interactions [PPIs]) is a fundamental integral component of complex biological systems. PPIs in yeast are identified through various experimental methods and are stored in curated databases. Integrating with functional genomics data and gene set enrichment analyses employing genome annotations such as GO terminology, several studies were performed to assign functions to proteins through coexpression, clustering, and homology-based methods (Arga et al., 2007; Brun et al., 2003; Chua et al., 2007; Piovesan et al., 2015). In this study, we applied an integrative biology approach using both interactome and transcriptome data to unravel the complexity of biology related to the uncharacterized Sld7 protein.
Reconstruction of cell cycle and chronological aging networks—Interactomic approach
The cell cycle (CCN1) and chronological aging networks (CAN1) were reconstructed from the physical interactome data of budding yeast (Karagoz and Arga, 2013). The number of interactions was significantly high at CCN1, which includes 70% of proteins in yeast, due to inclusion of proteins with unknown function (Table 1). This leads to a computational burden in the determination of highly interacting protein clusters and decreases the chance for estimating a function for Sld7. Therefore, the networks were reduced by eliminating the unknown molecular function annotation terms (so-called CCN2 and CAN2). The protein of interest, that is, Sld7, was added manually into the reduced networks. Moreover, it was proposed that Sld7 might have a role in early firing origin in G1 phase (Tanaka et al., 2011b). Therefore, the CCN involving the interactions in G1 phase (CCN3) was also reconstructed and examined further.
Identification of highly interacting protein clusters
The highly interacting protein clusters of reconstructed networks were identified using MCODE plug-in in Cytoscape software (Table 2). To determine the putative molecular functions of Sld7, it was necessary to examine clusters that involved Sld7 or any of its physical interactors, which are Sld3, Nup116, Mth1, Nab2, Ssa2, Srp40, and Hek2 (Karagoz and Arga, 2013). Sld7 cannot be encountered in any of the clusters formed from any of these networks. However, 9 of 118 clusters included its physical interactors (Fig. 3 and Supplementary Figs. S1 and S2). Sld3, which is also a genetic interactor of Sld7 (Tanaka et al., 2011b), was the most encountered protein among the physical interactors of Sld7 and existed in three clusters (Table 2).

Interactome network divided into highly interacting protein clusters, including the Sld3 (YGL113W), which is one of the physical interactors of Sld7.
Functional enrichment analyses of protein clusters
The nine clusters of the cell cycle and chronological aging networks were individually examined through functional enrichment analyses using GO molecular function terminology as function annotation. Although 83 distinct GO terms were encountered (see Supplementary Table S1 for analysis results), protein clusters of CCNs were significantly enriched in binding activities; more specifically, macromolecular complex binding (GO: 44877), microtubule plus-end binding (GO: 51010), RNA polymerase II C-terminal domain phosphoserine binding (GO: 1990269), and ribonuclease MRP activity (GO: 171).
On the other hand, clusters of aging networks (CANs) were significantly enriched with ribonuclease MRP activity (GO: 171), heme–copper terminal oxidase activity (GO: 15002), oxidoreductase activity, acting on a heme group of donors (GO: 16675), and chromatin binding (GO: 3682). When all clusters were taken into account, binding (GO:5488) and macromolecular complex binding (GO:44877) were the most repeated terms, whereas RNA polymerase II C-terminal domain phosphoserine binding (GO: 1990269) came into prominence as the most specific function term (Table 3).
Construction of coexpressed gene clusters—Transcriptomic approach
To check the reliability of the predicted function(s) of Sld7 given in Table 3, the gene expression data were employed. The PCCs of the expressed genes with respect to Sld7 were calculated [Eq. 1] for each dataset given in Table 4. The coexpressed gene clusters were constructed considering different r-value cutoffs and were examined by functional enrichment analyses.
GEO, Gene Expression Omnibus.
Cell cycle-related gene expression
The cell cycle-related gene expression data were downloaded from GEO database, released on February 8, 2016 (Table 4). Unfortunately, the coexpressed gene clusters were enriched with the molecular function unknown term in all the examined datasets.
Aging-related coexpression pattern
GDS362 dataset in Table 4 was focused on deletion of Sgs1 protein, which has a role in ATP-dependent DNA helicase activity. The results of the GO enrichment analysis of the clusters with r-value cutoff of 0.90 showed that the most significantly enriched molecular function term was oxidoreductase activity, acting on a heme group of donors (GO:16675). All annotated proteins associated with this term in genome were included in this cluster (represented by a background frequency of 1).
Interestingly, the enrichment results of coexpressed gene clusters were almost the same as the results of the highly interacting protein cluster analysis involving Hek2 at CAN1. This intersection increases the possibility of the role of Sld7 in chronological aging in yeast. It was reported that the oxidative damage in DNA affects replication and causes aging (Burhans and Weinberger, 2007). That result supports the dual role of Sld7 in both chronological aging and cell cycle processes in yeast.
In our previous study, Yucel et al. (2014) recorded the gene expression data for 882 genes in an environment [YPD (yeast extract peptone dextrose) medium] with and without pH control for both young and aged cells by high-throughput microarray experiment. To determine the highly correlated genes in our aging-associated gene expression dataset (Yucel et al., 2014), the PCCs of these 882 genes with respect to SLD7 were calculated and the same procedure as that of GDS362 dataset was followed to construct and analyze coexpressed gene cluster(s).
In young cells, the coexpressed gene cluster (with an r-value of 0.96 or higher) was significantly enriched with mRNA binding (GO: 3729) and poly(A) RNA binding (GO:44822). On the other hand, in aged cells, the transcriptional factor activity, RNA polymerase II transcription factor binding (GO: 1076), term significantly came into prominence.
Reconstruction of aging-associated coexpression network (CCAN)—Transcriptomic approach
As our aging-associated gene expression dataset (Yucel et al., 2014) represented high correlations between the majority of the gene pairs, the coexpressed gene network (CCAN) was formed using the transcriptome data obtained in pH-controlled and uncontrolled environments. In this CCAN network, the positive correlations with PCC equal or higher than 0.99 represented edges and the coexpressed genes represented nodes. The constructed networks (CCAN-aged and CCAN-young) were consisting of 882 nodes and 20,332 edges and 25,304 edges, respectively. The encoded proteins in the dataset were highly correlated with each other (Fig. 4). The number of proteins that are interacting with Sld7 was given as 13 in literature (Batisse et al., 2009; Hasegawa et al., 2008; Mantiero et al., 2011; Stirling et al., 2012; Tanaka et al., 2011b; van Pel et al., 2013; Yoshida et al., 2014). However, the numbers of proteins that were coexpressed with Sld7 in aged and young cells were 97 and 65, respectively. Then, MCODE plug-in was used to divide the CCAN network into clusters, which were ranked with respect to their Z-score, representing the effectivity of the results. In case of aged cells, Sld7 and Sld3 were encountered in two different clusters (Fig. 5a, b).
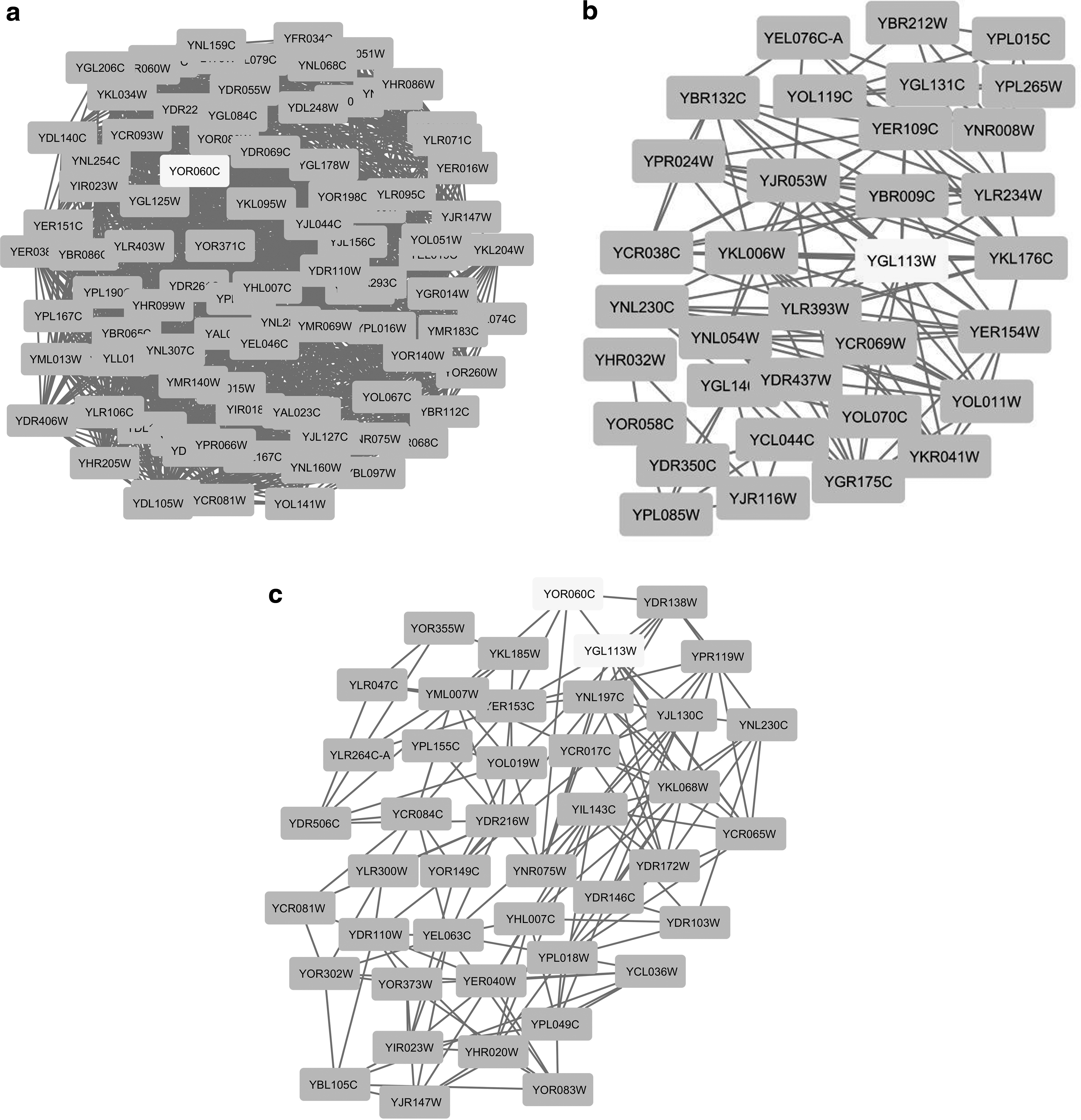
Among 16 clusters, the cluster involving Sld7 (CCAN-aged) has the highest score (Z-score: 46.801) and the cluster involving Sld3 has the fourth highest score (Z-score: 8.73). In case of young cells, Sld7 and Sld3 were encountered in the same cluster (Fig. 5c), which has the fourth highest score (Z-score: 7.095) among 17 clusters. The functional analysis of these clusters, performed through GO enrichment analysis, pointed out the terms of transcriptional repressor activity, RNA polymerase II transcription factor binding (GO: 1191); transcription factor activity, RNA polymerase II transcription factor binding (GO: 1076); and mediator complex binding (GO: 36033) as statistically significant based on hypergeometric test.
Information flow around Sld7 within the G1 phase cell cycle network, CCN3
As Sld7 might have a role in initiation of DNA replication in G1 phase of budding yeast (Tanaka et al., 2011b), information flow around Sld7 within the reconstructed G1 phase cell cycle network (CCN3) was analyzed through the linear path analysis, which is a valuable method to determine possible signaling pathways between the proteins in an interested organism (Arga et al., 2007; Steffen et al., 2002). The six subunits of the ORC (namely, Orc1, Orc2, Orc3, Orc4, Orc5, and Orc6), which are the initiator proteins that bind to replication origins during cell cycle (Labib, 2010), were selected as input proteins, and the proteins that have a role in initiation of DNA replication in G1 phase were set as the target proteins (Cdc7, Cdc45, Dpb11, Sld2, and Pol2) (Araki, 2011). The simulations were performed using Netsearch algorithm (Steffen et al., 2002) with different path lengths up to 7, which was the network diameter of CCN3, to identify the paths involving Sld7 (see Supplementary Table S2 for detailed analysis results).
Previously, it was reported that Sld7 forms a complex with Sld3 and decreases the affinity of Sld3 toward Cdc45 (Tanaka et al., 2011b). To analyze the interaction of these proteins in the initiation of DNA replication (early firing origin), information flow from ORC to Cdc45 was analyzed in CCN3. Sld7 was encountered in the paths from all ORCs to Cdc45. Sld7 was found to be either the fourth or fifth protein in the paths involving six or seven proteins, respectively. The upstream proteins of Sld7 were either Srp40 or Hek2; the downstream protein was always Sld3. Mcm2, Mcm5, Mcm7, Dot6, or Rad53 was also encountered in these paths. The analyzed information flows support the interaction of Cdc45 with Mcm2-7 recruited by Sld3, and also confirm the validity of our computational approach.
Sld is an abbreviation of synthetic lethality with Dpb11 (Tanaka et al., 2011b), which is one of the DNA replication initiation proteins, and is responsible for the formation of CMG complex through the interaction between preloading complex (pre-LC) and Sld3 (Muramatsu et al., 2010; Tanaka and Araki, 2013). Dpb11 is phosphorylated by CDK. This phosphorylation removes Sld3 and Sld2 from Dpb11, then GINS-Polɛ binds to pre-RC and forms the preinitiation complex (pre-IC). To estimate the possible role of Sld7 associated with Dpb11, the linear paths from ORC to Dpb11 were analyzed. We identified 287 paths starting with Orc1, Orc2, Orc3, Orc6, or Mcm2-7 and involving Sld7, which was encountered as the third or fourth protein in all the paths with seven proteins. The upstream protein that interacted with Sld7 was Mth1, Srp40, Sld3, or Hek2; the downstream protein was Sld3. The protein that was encountered in these paths was Mis1, Hta2, Ssb1, Hmo1, Mth1, Ssa1, Pex13, Psf1, or Ssa2. The analyzed paths were supported by the reported information flow (Masumoto et al., 2002).
Sld2 was one of the checkpoint proteins, which binds to Dpb11 and forms pre-LC complex, which is required for the initiation of DNA replication (Masumoto et al., 2002). It also has an essential role for CDK-dependent activation, which is related to chromosomal DNA replication (Muramatsu et al., 2010). To estimate the possible role of Sld7 associated with Sld2, the linear paths from ORC and minichromosome maintenance (MCM) to Sld2 were also analyzed. We identified 114 paths starting with Orc1, Orc2, Orc3, Orc6, or Mcm2-7 and involving Sld7 (Supplementary Table S3). Sld7 was encountered as the fourth protein in all the paths involving seven proteins. The upstream protein that interacted with Sld7 was Srp40, Sld3, or Hek2; the downstream protein was Sld3 or Mth1. The protein that interacted with Sld2 was Mis1, Hta2, Ssb1, Hmo1, Cdc28, or Clb5. The other protein that was encountered in these paths was Ssa1, Pex13, Psf1, or Dpb11. The analyzed paths were confirmed by the information flow reported in Masumoto et al. (2002).
Pol2 (Pol ɛ) is required for the initiation of DNA replication during mitotic DNA synthesis (Araki, 2011). Some of the paths starting with Orc1, Orc2, or Orc6 involved Sld7 (Supplementary Table S3). The paths of Orc2–Pol2 and Orc6–Pol2 involved the same proteins in the same order except the first two proteins (Orc and second one), but the proteins were different in the path of Orc1–Pol2. Sld7 was again encountered as the fourth protein and all the paths involved seven proteins. The upstream and downstream proteins that directly interacted with Sld7 were always Srp40 and Sld3, respectively. The protein interacting with Pol2 was always Psf1, one of the subunits of GINS complex. The other protein that was encountered in the paths was Hta2, Ssb1, or Hmo1.
Cdc7 is a kinase protein, the catalytic subunit of DDK (Dbf4-dependent kinase) (Aparicio, 2013). It is required for origin firing and replication fork progression in mitotic S phase through phosphorylation of Mcm2-7 complexes and Cdc45 (Labib, 2010). Sixteen paths starting with Orc1, Orc2, or Orc6 involved Sld7 (Supplementary Table S3). The Cdc7-interacting protein in these paths was Mis1, Hta2, Ssb1, or Hmo1. Sld7 was the fourth protein in all these paths, and its upstream protein (third or fourth) was either Srp40 or Hek2; the downstream protein was always Sld3. Rad53 (Sld6) or Mcm4 also existed in these paths.
Sld7-Sld3-Cdc45 complex was reported to bind to Mcm2 and Mcm7 (Bruck and Kaplan, 2015). Therefore, Mcm2-7 was chosen as the starting protein and the pre-LC proteins, which were Sld2, Pol ɛ, and Dpb11, were set as the target proteins. In all the paths involving Sld7, it interacted with Sld3. However, Cdc45-Sld3-Sld7 complex was only encountered in the paths starting with Mcm2 or Mcm7 and extending to Sld2 or Dpb11 (Supplementary Table S3).
The paths from Mcm2-7 to Cdc28, which is a cyclin-dependent kinase (CDK) protein and responsible for the regulation of chromosome dynamics, were also analyzed. The paths starting with Mcm2, Mcm3, Mcm5, and Mcm7 had Sld7. The information flow in these paths was found to be identical. Sld7 was encountered as third protein in the paths, interacting with Sld3 and Mth1 as upstream and downstream proteins, respectively. Cdc28 phosphorylates Dbp11 and separates pre-LC from the pre-IC. The signaling flow, found here from Mcm- to Cdc28, is supported by the information given in literature (Labib, 2010).
Rad53 has a genetic interaction with Sld7 and it physically interacts with Sld3 and Cdc7 in the analyzed linear paths. Rad53 (Sld6) is an essential protein for inheritance and it has a regulatory role in nuclear and mitochondrial DNA-related processes. DDK (Cdc7-Dfb4) and CDK (Cdc28) inhibit Rad53 activity in dependent pathways. The mutation of rad53-cdc7 caused synthetic lethality (Dohrmann and Sclafani, 2006). Rad53 phosphorylates Sld3 to inhibit the initiation of replication (Zegerman and Diffley, 2007). The linear paths found here supported the interaction of Rad53 with Sld3 and Cdc7 on computational basis.
The abovementioned results are in agreement with the published mechanisms occurring in cell cycle and DNA replication (Labib, 2010) and confirm that the networks were correctly reconstructed. In the initiation of DNA replication process, 160 different proteins were encountered in the linear paths, including starter and target proteins. In total, Sld7 was encountered in 1022 different pathways.
Among the proteins in these linear paths of information flow, Dot6, Hta2, and Hmo1 have a role in DNA binding (Bauerle et al., 2006; Wullschleger et al., 2006; Zhu et al., 2009), that is, they help gene products' noncovalent binding to DNA. Clb5 is responsible for transition from G1 phase to S phase (Schwob and Nasmyth, 1993). Psf1 is one of the subunits of GINS complex (Takayama et al., 2003). Ssa1 and Ssa2 are responsible for ATPase activity (Boorstein et al., 1994; Needham and Masison, 2008). These proteins can also be considered as assist proteins in binding the main proteins to each other, as indicated by the information flow of our linear path analysis. So far, an assisting role has not been mentioned in literature for any of these proteins at the initiation of DNA replication.
Discussion
In the present study, the putative molecular functions of Sld7 were studied by computational methods using an integrated approach of transcriptomics and interactomics. In the published literature, Sld7 was hitherto understood as one of the nonessential proteins with a role in initiation of DNA replication in budding yeast (Tanaka et al., 2011a, 2011b). However, in our previous experimental work on the effects of medium acidity on chronological aging using BY4742 strain of S. cerevisiae, the transcriptomics analysis revealed that the expression level of Sld7 was 0.5 (log2-fold change) under pH-controlled conditions, being in the top 10 among 882 genes (Yucel et al., 2014). This raises the question on a putative and critically important role of the Sld7 protein in aging.
The cell is prepared for initiation of DNA replication in G1 phase, and then proceeds to S phase, in which DNA replication occurs. The readiness of the cell is controlled through checkpoints in G2 phase before M phase, in which cell divides (Cooper, 2000). The initiation of DNA replication consists of two steps, which are origin licensing and activation. ORCs legitimize the replication origin at double-stranded DNA (dsDNA). Cdc6 and Cdt1 bind to ORC to load inactive MCM proteins onto replication origin (Aparicio, 2013) and then leave dsDNA.
The loaded complex and loading steps were called prereplication complex (pre-RC) and licensing, respectively. Pre-LC, which involves Dpb11, Sld2, GINS (partner of Sld5, Sld1, Sld2, and Sld3), and Pol ɛ, binds to CDK-phosphorylated Sld3 (Tanaka et al., 2007; Zegerman and Diffley, 2007). Cdc45 moves along the replication fork during replication of DNA, whereas pre-LC and Sld3 do not (Tanaka and Araki, 2013). Sld7 decreases the affinity of Sld3 to Cdc45 in early firing origin (Tanaka et al., 2011b). The GINS binds to Mcm2-7 to form Cdc45-Mcm-GINS (CMG) complex in late origin (Bruck and Kaplan, 2015), which is called replication fork helicase or pre-IC. The CMG complex is activated by DDK and then unwinds dsDNA in S-phase (Fig. 6).

The initiation of DNA replication mechanism in budding yeast:
We analyzed the physical interactions of Sld7 with other proteins in the reconstructed cell cycle and CANs of budding yeast. Several possible scenarios presented in the literature on the function of Sld7 were tested by analyzing the information flow patterns in the reconstructed networks. In addition, the coexpression patterns were analyzed by combining transcriptome data from several experiments with the interactome networks presented here to propose possible roles of Sld7 protein in aging of budding yeast. In these networks, the highly interacting protein clusters and significantly coexpressed gene clusters around Sld7 were further examined by functional enrichment analyses. The computational methods used here (Fig. 2) to annotate a function to Sld7 point out a dual role both in chronological and replicative (cell cycle) aging processes.
The comparison of the results of enriched clusters revealed that among 107 different terms, the most encountered function annotation term was macromolecular complex binding (GO: 44877), which means a protein interacting selectively and noncovalently with any macromolecular complex. Sld7 recruits GINS to Mcm2-7 by binding onto pre-LC, a macromolecular complex (Takayama et al., 2003), and decreases the affinity of Sld3 to Cdc45 in early firing origin in late G1 phase. This binding allows the formation of CMG complex (active helicase) that initiates DNA replication by unwinding dsDNA.
The examined information flows from Mcm2-7 to Sld2 and Dpb11 supports the theory about the role of Sld7 as macromolecular complex binding by its complex formation with Sld3 and Cdc45 and disassociating their interactions. In addition, Sld7 was encountered in paths from ORC to Cdc7, which is responsible for activation of DNA initiation by phosphorylating Mcm2-7 for early and late origins (Labib, 2010).
The upstream and downstream proteins that were encountered in analyzed paths include Mis1, Hta2, Ssb1, Hmo1, Clb5, Cdc28, Ssa1, Sld3, Srp40, Hek2, Mth1, Psf1, and Pex13, some of which have a role in DNA binding, localize generally in the nucleus, cytoplasm, and DNA initiation complex, and they are here suggested to function as assist proteins in DNA replication. The molecular functions of some of these proteins, such as Srp40, Mth1, and Psf1, are also unknown. These proteins might have role(s) in the formation of complexes in G1 phase of cell cycle in budding yeast.
Other most encountered function annotation terms were transcriptional factor activity, RNA polymerase II transcription factor binding (GO: 1076); RNA polymerase II C-terminal domain phosphoserine binding (GO: 1990269); and mRNA binding (GO: 3729). The regulator protein of Sld7, Med2 (mediator complex), is a subunit of RNA polymerase II and it is an essential protein for transcriptional regulation in yeast (Venters et al., 2011).
The genetic interactor of Sld7, Cdc7, has molecular functions of transcriptional factor activity, RNA polymerase II transcription factor binding (GO: 1076), and RNA polymerase II C-terminal domain phosphoserine binding (GO: 1990269). The molecular functions of Hek2 and Nab2, the physical interactors of Sld7, are mRNA binding and rRNA binding, respectively (Irie at al., 2002; Mitchell et al., 2013). The transcription process and rRNA binding occur in the nucleus and ribosome, respectively. Similarly, the localization of Sld7 was reported as nucleus (Hazbun et al., 2003; Huh et al., 2003) and nuclear envelope-ER (Hazbun et al., 2003). Sld7 also colocalizes with spindle pole body (Huh et al., 2003). The results of GO enrichment analyses indicate that one of the putative functions of Sld7 might be related to transcriptional repressor activity binding to RNA polymerase directly.
If Sld7 is not expressed during the cell cycle, the CMG complex, which is an active DNA helicase at replication fork (Tanaka et al., 2011b), cannot be formed and DNA replication cannot be initiated. During nutrient depletion, the cells' arrest in G1 phase causes chronological senescence (Burhans and Weinberger, 2007). We did experiments in 1 nL microbioreactors, where the sld7Δ mutant cells were grown in both glucose-rich (2% glucose) and calorie-restricted (0.2% glucose) environment. The duration of G1 phase is observed to increase in calorie-restricted medium (unpublished data; Table 5).
YNB = yeast nitrogen base; YPD = yeast extract peptone dextrose.
In our computational analysis, the enriched molecular functions of clusters, obtained using aging coexpression dataset (GDS362; Fry et al., 2003) and CAN (Yucel et al., 2014), are heme–copper terminal oxidase activity (GO: 15002); oxidoreductase activity, acting on a heme group of donors (GO: 16675); and cytochrome-c oxidase activity (GO: 4129). The damaged DNA induces the genes involved in cellular redox homeostasis in yeast (Tkach et al., 2012) and leads to increased ROS concentration, and thus prevents cell proliferation, and causes cell death (Martindale and Holbrook, 2002).
Oxidative stress forces cells to autophagic cell death, where cells begin to shrink, bleb, and die (Zakeri et al., 1995). It was predicted (although still unclear) that the main source of oxidative stress was mitochondria (Burhans and Weinberger, 2007). The cellular component of the proteins, Cox4, Cox5, Cox6, Cox8, Cox9, Cox12, and Cox13, responsible for these oxidation-related functions and present in highly correlated clusters of coexpressed genes, are mitochondria.
Altmann and Westermann (2005) did a study about the essential cellular genes/proteins in mitochondrial morphogenesis in yeast and suggested that the proteins, erg29, fcf1, pga2, and sld7, with unknown molecular function might be contributing to mitochondrial inheritance directly or indirectly. Hence, in addition to its role in cell cycle, Sld7 might have a role in keeping the level of ROS low and enhancing life span of the cell by reducing oxidative damage in mitochondrial DNA. In literature, the other protein with a dual role in both chronological and RLS is Tor1, and similar to Sld7, it is also responsible for macromolecular complex binding in yeast (Wullschleger et al., 2006).
Conclusion and Outlook
Sld7 was hitherto considered a nonessential and poorly characterized protein that interacts with Sld3 and decreases its affinity toward Cdc45 in early firing origin in G1 phase of cell cycle in budding yeast (Araki, 2010; Tanaka et al., 2011b). On the other hand, the role of Sld7 might resemble MTBP, which is one of the binding proteins of Treslin/Ticrr (homolog protein of Sld3) in human cells. Depletion of both proteins causes delay in S phase (Boos et al., 2013). The significant change in the gene expression level of Sld7 in chronological aging-related experiments (Yucel et al., 2014) motivated us to investigate the role of Sld7 in chronological aging process of budding yeast in addition to its function in cell cycle. The interactome and transcriptome data of the budding yeast were used to construct the networks (CCN, CAN, CCAN) and the clusters that were formed from these networks were examined through GO enrichment analyses.
The information flow, analyzed here through DNA replication initiation, supports that Sld7 has a role in G1 phase and forms a complex with Sld3 and Cdc45 (Sld4), being in agreement with literature (Tanaka et al., 2011b). The most repeated GO molecular function term, macromolecular complex binding (GO: 44877), might be the function of Sld7 in budding yeast, that is, binding of GINS (part of pre-LC complex) to Mcm2-7 and Cdc45 to form CMG complex, which initiates DNA replication. In the case of the depletion of Sld7, the GINS-Pol ɛ complex may not be able to bind to pre-RC, and hence pre-IC could not be readily formed. Consequently, the cell cycle is stuck in G1 phase for a while, causing replicative aging (Burhans and Weinberger, 2007).
The other molecular function terms acquired from the network, reconstructed from aging-related transcriptome data, were heme–copper terminal oxidase activity (GO: 15002); oxidoreductase activity, acting on a heme group of donors (GO: 16675); and cytochrome-c oxidase activity (GO: 4129). The proteins responsible for these GO terms are generally localized in mitochondria (Renvoisé et al., 2014). Sld7 might have a role in mitochondria (Altmann and Westerman, 2005). Therefore, one can hypothesize that Sld7 might be expressed in both mtDNA and DNA. The role of Sld7 might be related to the decrease in activity of ROS by preventing oxidation damages to DNA.
We propose that Sld7 might slow down aging process of the cell by keeping the level of ROS low and preventing oxidative damage of DNA. In the light of this information, we conclude that the enriched function terms point out a dual role for Sld7 in both cell cycle and chronological aging processes in yeast.
In higher eukaryotes, the proteins that have a role in the initiation of DNA replication are replication-stimulating protein (Treslin) (homolog of Sld3) and MTBP (putative homolog of Sld7) with topoisomerase-II beta-binding protein 1 (TopBP1) (homolog of Dpb11) (Boos et al., 2013). The expression level of MTBP has been found to be elevated in several cancer types (Alam et al., 2012; Bassi et al., 2016; Boos et al., 2013; Chen and Butte, 2016; Grieb et al., 2014; Han et al., 2016). In human cells, regulation of tumor suppressor protein p53 is controlled through MDM2, which inhibits the activity of p53. The regulation of this protein depends on a variety of cellular stresses that cause DNA damage, which in turn may cause abnormality or cancer (Leri et al., 1999; Smith et al., 2000). Once p53 is activated under stress conditions, the cell cycle is arrested in G1 phase and the system tries to repair the damaged DNA or leads to apoptosis to eliminate damaged cells (Chen et al., 1996; Haupt et al., 1996). However, if MDM2 is overexpressed, it blocks p53 function and leads to tumor development. Thus, it can be suggested that MTBP has a role in regulation of p53 indirectly through the MDM2 pathway (Alam et al., 2012).
Elucidation of the role of Sld7 in yeast can help to understand the role of its putative human homolog MTBP better as well as to shed light on the development of novel strategies for the treatment of cancers and aging-related diseases.
Footnotes
Acknowledgment
The authors gratefully acknowledge the financial support of Scientific Research Fund of Boğaziçi University through project R9701.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.