
Abstract
Abstract
Disruptive innovations in medicine are game-changing in nature and bring about radical shifts in the way we understand human diseases, their treatment, and/or prevention. Yet, disruptive innovations in cancer drug design and development are still limited. Therapies that cure all cancer patients are in short supply or do not exist at all. Chief among the causes of this predicament is drug resistance, a mechanism that is much more dynamic than previously understood. Drug resistance has limited the initial success experienced with biomarker-guided targeted therapies as well. A major contributor to drug resistance is intratumor heterogeneity. For example, within solid tumors, there are distinct subclones of cancer cells, presenting profound complexity to cancer treatment. Well-known contributors to intratumor heterogeneity are genomic instability, the microenvironment, cellular genotype, cell plasticity, and stochastic processes. This expert review explains that for oncology drug design and development to be more innovative, we need to take into account intratumor heterogeneity. Initially thought to be the preserve of cancer cells, recent evidence points to the highly heterogeneous nature and diverse locations of stromal cells, such as cancer-associated fibroblasts (CAFs) and cancer-associated macrophages (CAMs). Distinct subpopulations of CAFs and CAMs are now known to be located immediately adjacent and distant from cancer cells, with different subpopulations exerting different effects on cancer cells. Disruptive innovation and precision medicine in clinical oncology do not have to be a distant reality, but can potentially be achieved by targeting these spatially separated and exclusive cancer cell subclones and CAF subtypes. Finally, we emphasize that disruptive innovations in drug discovery and development will likely come from drugs whose effect is not necessarily tumor shrinkage.
Introduction
C
Unless detected early, cancer beyond the primary site is difficult to control and cure. Despite our best efforts in making healthcare available to all and after spending billions of dollars in an attempt to eradicate cancer, cancer still remains as one of the deadliest disease known to mankind today. Worldwide, cancer deaths outnumber total deaths from diseases such as TB, HIV/Aids, and malaria combined (Ferlay et al., 2015).
Disruptive innovations in medicine are game-changing in nature and bring about radical shifts in the way we understand human diseases, their treatment, and/or prevention. Yet, disruptive innovations in cancer drug design and development are still limited. Therapies that cure all cancer patients are in short supply or do not exist at all. Chief among the causes of this predicament is drug resistance, a mechanism that is much more dynamic than it was previously understood. A major contributor to drug resistance is intratumor heterogeneity. For example, within solid tumors, there are distinct subclones of cancer cells, presenting profound complexity to cancer treatment. This expert review explains that for oncology drug design and development to be more innovative, we need to take into account intratumor heterogeneity.
Methodology: Search Strategy and Article Selection
PubMed, Google Scholar, and ScienceDirect were searched (from year 2000 to year 2017) for the following terms: solid tumor, cancer, genomic instability, intratumor heterogeneity, chemoresistance, targeted therapy, clonal evolution, cancer stem cells (CSCs), subclones, deep sequencing, next-generation sequencing, and drug development. These databases are specialized in human diseases, such as cancer, and innovative technologies, and mostly use English as the main language. Duplicate articles were removed and only full-text articles were considered. Our citations also show a few earlier years publications to accommodate origins of certain theories such as the clonal evolution theory by Nowell, 1976.
Cancer: Not Everyone Fits the Mold
Our understanding of cancer has improved greatly with new data showing that tumor heterogeneity contributes immensely to the lack of treatment success and the development of chemoresistance (Gerlinger et al., 2014a, 2014b, 2015; Greaves and Maley, 2012). The ability to analyze tumor heterogeneity observed in solid tumors through second-generation whole-genome sequencing has increased greatly in the last decade (Mateo et al., 2014). Traditionally, pathologists examine the whole tumor, but when it comes to making a diagnosis, they only report the highest grade (Fisher et al., 2013; Myers et al., 2012; Seoane and De Mattos-Arruda, 2014; Wang et al., 2015).
The implications of intratumor heterogeneity are profound, affecting both the diagnosis and clinical management of different cancers. During treatment, intratumor heterogeneity shows itself in the nonuniformity of drug response (Cooke et al., 2011; Turner and Reis-Filho, 2012; Yap et al., 2012). It is therefore important to identify subgroups of patients who do not respond to standard chemotherapeutic treatments. Treatment strategies utilizing targeted therapy will only eradicate a small population of cancer cells and is effective in few patients, and hence the transition toward personalized medicine.
However, several challenges remain to be addressed before personalized medicine is successful. Challenges that need to be addressed before personalized medicine can deliver on its promise include the lack of disease signature, the predictive power of disease models needs to be upped, and biomarkers are not very specific. Biomarkers must be authenticated by looking at their correlation with clinical outcomes and not just factors such as tumor shrinkage. There is a bias toward genomics at the expense of everything else, yet we know that genetic, epigenetic, and environmental factors are inseparable with regard to causing cancer. Genomics should not be the only platform used in the development of innovative chemotherapeutic agents. Putting all our “eggs in one basket”, genetics that is, will not solve issues associated with a complex disease such as cancer.
Most cancers present in old age partly due to the weakening of the suppressive tumor microenvironment, which acts by stalling growth of the tumor (Burrell and Swanton, 2014a; Burrell et al., 2013). In addition, cancer cells compete among themselves and with stromal cells for nutrients and space. Chemotherapy not only kills cancer cells but also adds a selective pressure that can result in the appearance and proliferation of chemoresistant clones (Ding et al., 2012). Furthermore, chemotherapy can induce new mutations in cancer cells, leading to cancer cells with increased malignant potential, and can initiate relapse of the cancer (Hunter et al., 2006). The tumor microenvironment can alter cancer cell response to chemotherapy. The probability of any human to acquire mutations is high, this is partly due to the vast human genome (Lynch, 2010; McGranahan and Swanton, 2015).
This is true for solid tumors, with many having several somatic mutations. Many genetic changes are harmless, while some are easily repaired through normal DNA repair mechanisms (McGranahan and Swanton, 2015; Nowell, 1976). However, a few genetic changes will confer a selective advantage, resulting in a cancer cell with increased proliferative abilities (Lawrence et al., 2014). Such a cancer cell will outcompete the other cancer cells or normal cells and become the dominant clone within the solid tumor. Studies focussing on single cancer cells have shown that most genetic mutations are not present in all cancer cells; thus cancer cells are genetically heterogeneous (Yates and Campbell, 2012).
Even though combination therapy is the standard practice, a sizable number of especially advanced cancers are incurable, partly due to the metastatic disease and drug resistance (Saunders et al., 2012). Of late, systematic therapeutic strategies aim to exploit specific aberrations within solid tumors, creating the “so called” targeted and nontargeted therapies. Such therapies include inhibitors of growth factors, inhibitors of angiogenesis, inhibitors of chemokines, antistromal drugs, and epigenetic modifiers (Kwak et al., 2010). Patients' responses to these new and innovative therapies have been varied, with mortality remaining high (Garraway and Janne, 2012).
One of the reasons this is so is the development of drug resistance in most cancers (McGranahan and Swanton, 2017; Saunders et al., 2012). Tumor heterogeneity impacts precision medicine and the development of therapeutic agents. Drug development has to take into account tumor heterogeneity to make informed decisions on the cost and therapeutic benefit of such innovations (Burrell et al., 2013; Fojo et al., 2014). Due to genomic instability, the number of mutations present in a tumor tends to increase over time; thus targeted therapy might not target mutations in all cancer cells. Some subclones might have mutations conferring drug resistance even before the start of treatment. It is also known that some dominant subclones are supported by low-occurring subclones within the solid tumor (Gerlinger et al., 2015; Nowell, 1976).
The increased expression of drug proteins and drug-metabolizing enzymes, including the cytochrome P450 superfamily and survival pathways, are some of the common cell autonomous mechanisms involved in drug resistance (Dzobo et al., 2016; Fodale et al., 2011; Fukuda and Schuetz, 2012; Senthebane et al., 2017). These cell autonomous mechanisms, in addition to tumor microenvironmental factors, compromise the efficacy of drugs in varied ways (Fukuda and Schuetz, 2012; Saunders et al., 2012; Senthebane et al., 2017). Several therapeutic strategies have been used to modify and interfere with these mechanisms and proteins. In most cases, however, drug resistance develops and the disease relapses and progresses (Chan et al., 2017; Saunders et al., 2012).
Obviously, a deeper understanding of the dynamic nature of the cancer disease and the development of drug resistance is needed. Most important is the role of intratumor heterogeneity in drug resistance. Within solid tumors are distinct cancer cell subclones varying in mutations and tumorigenic potential, therefore varying in responses to drugs (Wei et al., 2017; Yin et al., 2017). Overall, the phenotype of a cancer cell at any given time is the sum of the contributions of the genotype, epigenetics, transcriptome, proteome, and microenvironmental interactions (Fig. 1). It is clear that not all cancer cells “fit the mold” and cancer drug design and development must take this into account. This review explores the clinical implications of tumor heterogeneity and strategies that can be used to overcome solid tumor heterogeneity.

The cancer cell phenotype is the result of the interplay between the genome, epigenome, transcriptome, proteome, and the microenvironment. At any given time, a cancer cell exists in a certain state. The cancer cell state plays a significant role in the cancer cell drug response.
Intratumor Heterogeneity
The presence of cancer cell subpopulations differing both genetically and phenotypically and therefore having distinct biological behaviors within a tumor is what is known as tumor heterogeneity. Somatic aberrations such as chromosomal doubling, single nucleotide or base changes, and gene amplifications are a result of the increased genome instability associated with cancers (Burrell and Swanton, 2014b). Several platforms and bioinformatic softwares have been developed to elucidate the involvement of certain mutations or somatic aberrations in certain cancers (Nik-Zainal et al., 2014). The major driving force for this intratumor heterogeneity is genomic instability (Campbell et al., 2010; Sieber et al., 2003; Yachida et al., 2010). Most cancers show increased mutations, chromosomal alterations, and production of reactive oxygen species, and this leads to genetic instability (Negrini et al., 2010; Turner and Reis-Filho, 2012).
While some genetic alterations are deleterious, genetic alterations giving a specific clone a survival advantage will result in that clone being dominant only under specific conditions (Ashworth et al., 2011; Turner and Reis-Filho, 2012). Genomic instability therefore creates a pool of mutations that are then selected by factors such as microenvironmental pressures, which give rise to the heterogeneity observed in solid tumors (Burrell et al., 2013; Ding et al., 2012; Gerlinger et al., 2014a; McGranahan and Swanton, 2015; Nowell, 1976). Each region of a solid tumor is eventually associated with a specific mutational signature, reflecting both the somatic aberrations and the role of microenvironmental pressure (Ding et al., 2012; Gerlinger et al., 2015).
Several mutational signatures, usually of unknown origins, have been identified in different cancers and these illustrate the diversity of the genomic instability processes at play during carcinogenesis (Govindan et al., 2012; Yates and Campbell, 2012). Some of the mutational signatures are associated with age such as the single-nucleotide level transversion C>T, which occurs mostly at CpG islands (McGranahan and Swanton, 2015, 2017). Mutations such as C>T and C>G, usually at TpC sites, are associated with apolipoprotein B mRNA editing enzyme, catalytic polypeptide-like (APOBEC) cytidine deaminases (Burns et al., 2013; Henderson et al., 2014). Many tumors driven by the human papillomavirus show increased presence of APOBEC motifs (Henderson et al., 2014).
The source of intratumor heterogeneity and its understanding has enormous implications on how cancer can be treated and the management of cancer patients (Saunders et al., 2012; Yap et al., 2012). A mutation in an important gene in one cell is usually the beginning of cellular transformation toward cancer. This cell passes on its mutation to its daughter cells during cell division. Daughter cells, in addition to the original mutation, go on to acquire more mutations and if these mutations give the cells an advantage over other cells, they survive and ultimately outcompete other nonmutated cells (Ding et al., 2012; Gerlinger et al., 2015; Yates and Campbell, 2012). This is the basis on which the clonal evolution model of tumor evolution is based (Fig. 2A) (Polyak, 2007; Shackleton et al., 2009).

Clonal Evolution and the CSC models.
In this model, the ability to form new tumors is the same for all subclones. The other model is the CSC model (Fig. 2B) (Bonnet and Dick, 1997; Lapidot et al., 1994; Shackleton et al., 2009). In this model, the ability to form new tumors is restricted to a rare subpopulation of cancer cells. This rare subpopulation of cancer cells, which is sometimes referred to as CSCs or tumor-initiating cells, has self-renewal abilities and is generally more resistant to therapy (Al-Hajj et al., 2003; Bonnet and Dick, 1997; Dzobo et al., 2016; Lapidot et al., 1994; Li et al., 2007). These rare cells have been reported in several cancer types, including leukemia and breast, colon, head, neck, and esophageal cancer. It is important to note that these two models are not exclusive. Both models give allowance for the formation of distinct cancer cell variants, differing both genetically and phenotypically (Bonnet and Dick, 1997; Nowell, 1976).
The clinical implications of the two models are vast. For example, for treatment to be successful, treatment of cancers following the clonal evolution model will require that all cancer cells must be eradicated, whereas in the case of the CSC model, the eradication of the rare subpopulation of CSCs might suffice for successful treatment. Evidence of transdifferentiation of cancer cells into different subclones, however, calls for caution on the possible success of therapy by just removing CSCs (Shackleton et al., 2009).
Drivers of Drug Resistance in Cancer
Resistance to therapy is almost certain in patients with cancer. Cancer cell resistance to drugs can be acquired or inherent. Several cellular processes are involved in mediating these phenomena and these include increased drug transporter expression, alteration in drug metabolism, enhanced DNA repair, and alterations in apoptotic pathways (Dzobo et al., 2016; Fodale et al., 2011). These processes can also act in combination to increase the chances of cancer cell survival. Given that intratumor heterogeneity gives rise to cancer cells differing genetically and phenotypically, it is easy to see that tumor heterogeneity plays a key role in cancer cell chemotherapeutic responses. To improve patients' survival, it is imperative to understand how contributors of tumor heterogeneity such as genetic variations, the microenvironment, cellular plasticity, and stochastic processes impact drug resistance (Saunders et al., 2012).
It should be noted that tumors also display phenotypic heterogeneity caused by factors besides intratumor heterogeneity (Marusyk and Polyak, 2010; Marusyk et al., 2014; Turner and Reis-Filho, 2012). Variation in the phenotypic traits of cancer cells can also come from epigenetic sources as well as through interactions with the tumor microenvironment (Dzobo et al., 2016; Krause et al., 2010; Senthebane et al., 2017).
The ABC family of transporters pumps toxins and drugs out of cells, thereby reducing the concentration of drugs within cells to sublethal levels (Fig. 3) (Fukuda and Schuetz, 2012; Saunders et al., 2012). Breast and esophageal cancer cells have enhanced DNA repair capabilities and can easily repair DNA damage caused by drugs. The cytochrome P450 family of enzymes is one example of drug-metabolizing enzymes responsible for the biological transformation of xenobiotics, including drugs into nontoxic compounds or molecules (Kumaraguruparan et al., 2006; Nebert and Dalton, 2006; Pool-Zobel et al., 2005; Yoxall et al., 2005). An increased activity of these enzymes means drugs are easily converted into nontoxic compounds or molecules, invoking drug resistance.
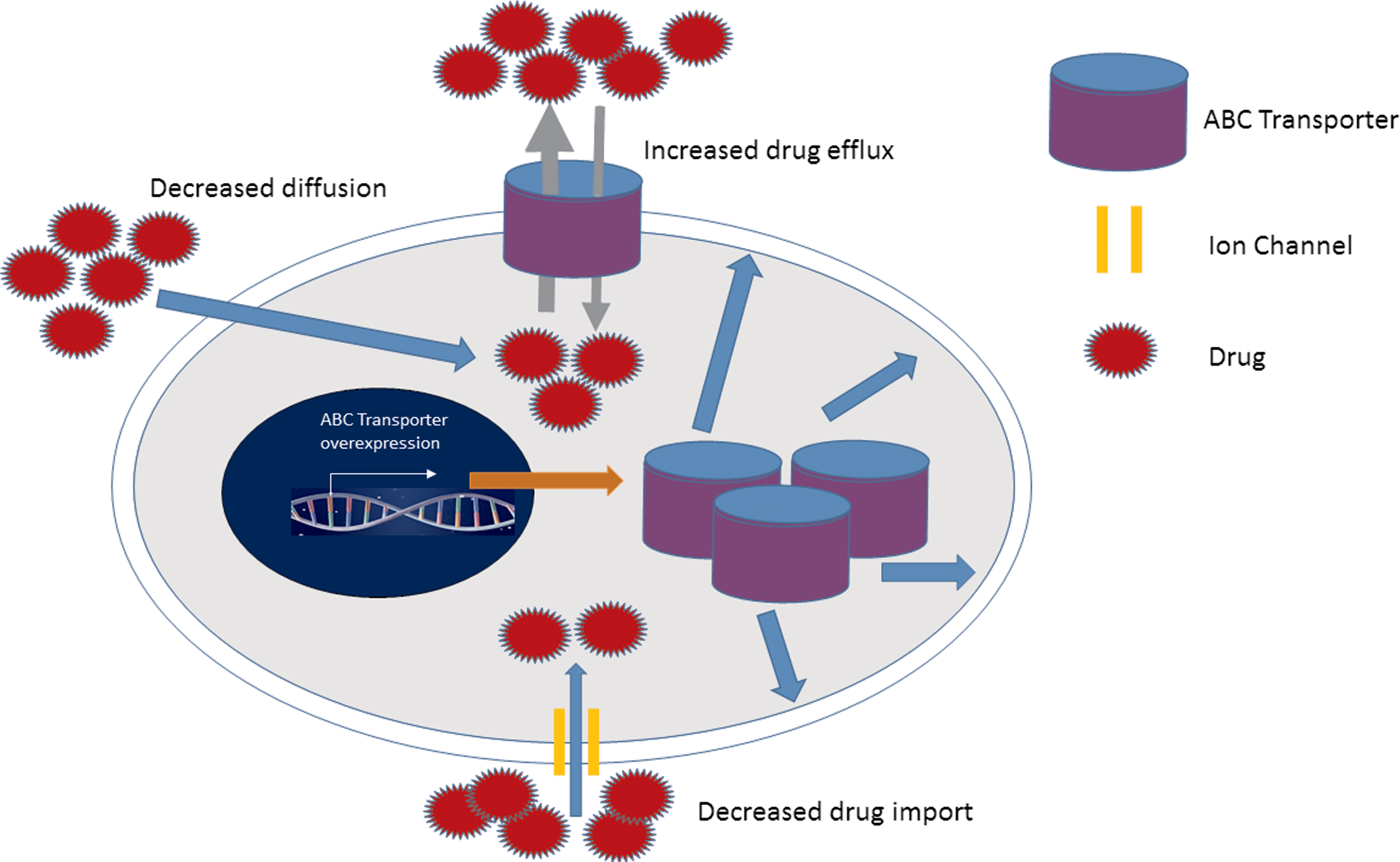
The involvement of ABC transporter proteins in chemoresistance. Increased expression of several membrane drug transporters such as the ATP ABC transporter family has been implicated in the development of chemoresistance. ABC proteins contribute toward chemoresistance through decreased drug uptake and increased drug efflux. ABC, adenosine binding cassette; ATP, adenosine triphosphate.
Last, the balance between proapoptotic and antiapoptotic processes or pathways determines whether cancer cells will be sensitive or resistant to drugs. Beside these cell autonomous mechanisms, other factors that can affect the effectiveness of drugs include the diffusion of drugs through the tumor, the size of tumor (affecting oxygen and nutrient availability), and the involvement of stromal components.
Even with thorough knowledge of the above mechanisms, resistance is still reported in many cancers (Acerbi et al., 2015; Alderton, 2017; Apte et al., 2015; Au et al., 2016; Bilen et al., 2016; Dauer et al., 2016; Di Marzo et al., 2016; Safa, 2016). CSCs have been shown to be resistant to therapy in many cancers, including breast, head, and neck and glioblastoma cancers (Bao et al., 2006, 2013; Rich and Bao, 2007; Xie et al., 2014, 2015). This has been attributed to the enhanced expression of ABC-transporter proteins used in drug secretion and to increased DNA damage repair processes (Bao et al., 2006; Brown et al., 2017; Chen et al., 2016; Lee et al., 2016; Liu et al., 2006; Mathews et al., 2011).
For tumors following the CSC model therefore, targeting these mechanisms can kill all CSCs and eradicate the cancer. For tumors showing clonal evolution, there is need to understand the subclonal architecture of the tumor and monitor changes to clone composition before, during, and after treatment. Alterations to the genome also contribute toward therapy resistance. Analysis of tumors before and after therapy clearly shows that it is possible for a minor subclone to become dominant after therapy (Anderson et al., 2011; Burrell et al., 2013; Ding et al., 2012; McGranahan and Swanton, 2017; Meacham and Morrison, 2013). Thus, the genetic differences between subclones can determine which subclone survives after therapy.
Genetic Variation
Recent data from single cancer cell analysis using “omics” techniques such as next-generation sequencing and transcriptomic analysis have shown massive intratumor heterogeneity in solid tumors for breast, pancreatic, kidney, and colon cancers (Ding et al., 2012; Saunders et al., 2012; Tirosh et al., 2016; Wei et al., 2017; Yap et al., 2012; Zhang et al., 2014). Not only are distinct cancer cell subpopulations present in solid tumors, these distinct subpopulations are also regionally located within the solid tumors (Gerlinger et al., 2014a; Saunders et al., 2012). Only a fraction of mutations is usually shared among all cancer cells within tumors. This distinct subpopulation of cancer cells differs not only in their genetic makeup but also in their sensitivities to drugs, and their ability to self-renew and initiate new tumors. (Bonnet and Dick, 1997; Burrell and Swanton, 2014a; Ding et al., 2012).
Over the years, it has been observed that after the initial response to chemotherapy, many cancer cells show resistance after being exposed to the same drug again (Fodale et al., 2011). The same scenario is observed even with new improved targeted therapies. The use of multiple drugs concurrently and second-generation drugs with broader specificity can overcome such resistance. Drug resistance during targeted therapy is clear evidence of the existence of distinct subclones within the solid tumor (Sanchez-Laorden et al., 2014; Saunders et al., 2012).
It should be noted that therapy itself can be a source of intratumor heterogeneity as cancer cells are exposed to different amounts of drugs in a solid tumor (Gerlinger et al., 2014a; McGranahan and Swanton, 2015). Usually new subclones are identified when a tumor relapses. Therapy is known to alter the subclone composition of a solid tumor, resulting either in new subclones altogether or one or two subclones outcompete the rest. These data point to the need to monitor or manage cancer patients during and after treatment.
During treatment, therapy also acts as a source of genomic instability (Johnson et al., 2014; Zhang et al., 2014). Increased mutations are usually observed in solid tumors during or after therapy compared to before treatment (Ding et al., 2012). Importantly, several aberrations such as insertions, deletions, and structural variants are associated with cisplatin treatment (Meier et al., 2014). At the single-nucleotide level, cisplatin and temozolamide treatments resulted in an increase in C>A and C>T transversions in Caenorhabditis elegans, respectively (Johnson et al., 2014; McGranahan and Swanton, 2015).
Increase in mutation load in recurrent tumors, of about 6–10 times compared to primary tumors, especially of the brain, has been reported extensively. Therapy therefore induces some mutations as well as acts as a selection barrier, influencing the evolutionary direction of the tumor (Bianchi and Ghobrial, 2014; Burrell et al., 2013; Fisher et al., 2013). The resultant tumor is sometimes more resistant to therapy and more aggressive. Therapy can also result in new mutations on dormant clones and since these were not targeted initially, they could be resistant to therapy and can therefore become the dominant clones posttherapy.
Therapeutic response to cisplatin has also been observed to depend on homologous recombination, a form of copy number aberration (Birkbak et al., 2012). Copy number variation can be used to predict the chromosomal instability observed in cancer cells (Birkbak et al., 2011). Certain tumors show a loss of some chromosomes. Chromosomal fragmentation can result in pieces of DNA joining to other chromosomes during DNA repair processes (Zhang et al., 2016b). Whole-genome doubling is a common event in several cancers (Dewhurst et al., 2014; Zhang et al., 2014, 2016b). Analysis of single tumor samples has revealed that branched tumor evolution occurs in several cancer types (Anderson et al., 2011; Johnson et al., 2014; Yates and Campbell, 2012).
Tumor subclones interact with one another through paracrine mechanisms (Calbo et al., 2011). Such cooperation between clones has been reported widely and is known to activate several survival signaling pathways and the release of growth factors and cytokines (Chapman et al., 2014). It is known that certain tumor subclones are only metastatic in the presence of other subclones or cells (Calbo et al., 2011). Co-engrafting of cetuximab-resistant and cetuximab-nonresistant subclones resulted in the survival of the nonresistant subclones as well (Hobor et al., 2014). This was through the release of growth factors necessary for the survival of the nonresistant subclones.
On the other hand, cooperation between subclones can also result in tumor collapse when one subclone outcompetes another subclone on which it depends on for survival (Marusyk et al., 2014). Tumor heterogeneity therefore appears to be a necessity for the continual survival and progression of a tumor. For drug development and precision medicine, this means there is need to target even the minor subclones that are needed for the survival of the dominant subclones (McGranahan and Swanton, 2015).
From a clinical point of view, the targeting of dominant subclones, although appealing, is a futile exercise given the above observations (McGranahan and Swanton, 2015). Different cancers obviously follow different evolutionary routes (Landau et al., 2013; Wang et al., 2017a). For example, in breast and esophageal cancers, TP53 mutations are generally thought to be early events, while in clear cell renal cell carcinomas, TP53 and PTEN mutations are late events with von Hippel-Lindau gene mutation as an obligatory early event (Aiello et al., 2016; Campbell et al., 2010). KRAS and BRAF mutations are early events in colorectal cancer (Brannon et al., 2014). Different patients with the same type of cancer may display different evolutionary routes as well. In multiple myeloma, some patients show BRAF mutations as driver events, while in others, these mutations occur on the tumor's phylogenetic tree's branches as subclonal (Bolli et al., 2014).
In some patients, early somatic aberrations influence the evolutionary trajectory of a tumor (Burrell and Swanton, 2014a; Gerlinger et al., 2014b). These results appear to show that certain mutations or early events predestine the tumor to a particular evolutionary stream. Parallel evolution is shown in clear cell renal cell carcinoma with different regions of the tumor displaying distinct somatic mutations affecting the same gene or pathway (Gerlinger et al., 2014a, 2014b). A deeper understanding of the role of the microenvironment may help in predicting tumor evolutionary paths and possible treatment for such eventualities, rather than the current reactionary approach (McGranahan and Swanton, 2015, 2017; Nowell, 1976).
It is well established that chromosomal instability-induced tumor heterogeneity results in poor prognosis in many cancers (McGranahan et al., 2012). However, in many cancers, the total of driver events determines the outcome (Molenaar et al., 2015; Woll et al., 2014). Driver mutations or events being genetic aberrations provide survival advantage to cancer cells or are important for the continual cancerous behavior of the cells (Turner and Reis-Filho, 2012). The instability of the cancer genome allows subclonal changes and this results in gain and loss of certain mutations during cancer progression. Combining genomic instability and microenvironmental influence results is a complex system that needs further and thorough examination (Melchor et al., 2014).
Stochastic Processes
Stochastic processes are processes that result in transient phenotypic differences or “noise” between cells in a pool of genetically identical cells. Any cell population displays a natural variation and this can contribute to a variation in chemotherapeutic response (Albeck et al., 2008; Saunders et al., 2012; Spencer et al., 2009). Transcription is inherently random in that, each gene has two alleles, resulting in differences in gene expression. This natural phenotypic variation of cells illustrates the stochastic nature of cellular biochemical processes (Marusyk et al., 2014). A pool of genetically identical cancer cells expressing proteins associated with the apoptosis pathways showed different time points needed to induce apoptosis (Saunders et al., 2012; Spencer et al., 2009). Thus, the heterogeneity observed in this study does not rely on genetic or epigenetic mechanisms, but on differences in the expression of proteins. This has fundamental implications to therapy. Thus, given the same genetic background and physical conditions, some cancer cells will be more sensitive to chemotherapeutic agents than others.
The difference in response to chemotherapeutic agents may also arise due to the differences in stages of cell cycle at the time drugs are added. However, the stage of cell cycle accounts for about 20% of the variation, with the bulk coming from cell to cell differences in the expression of proteins of targeted pathways (Caiado et al., 2016; Cohen et al., 2008). Some cancer cells require more exposure time to drugs to undergo apoptosis. Cells with the same genome and under the same physical environment do not necessarily have the phenotype. Phenotypic heterogeneity has been shown in genetically identical cancer cells through single cell analysis (Almendro et al., 2013, 2014; Caiado et al., 2016).
Microenvironmental Factors
The interplay between genetic and environmental factors is fundamental to the pathogenesis of cancer. The tumor stroma has been shown to play a key role in the way cancer cells respond to several stimuli, including drugs (Dzobo et al., 2016; Saunders et al., 2012; Senthebane et al., 2017). Perhaps this is not surprising given that tumor stroma contributes more than half of the tumor mass. The stroma is composed of both cellular and noncellular components. These include cancer-associated fibroblasts (CAFs), macrophages, blood vessels, nerves, and the extracellular matrix (ECM).
The ECM, especially 3D ECM, is known to directly influence the behavior of cancer cells (Dzobo et al., 2016; Saunders et al., 2012; Senthebane et al., 2017). A close proximity to the ECM is known to increase the survival of cancer cells. Cells interact with the ECM mainly through receptors such as integrins (Hynes, 2013; Hynes et al., 2002). Increase in integrin expression was shown to result in the activation of antiapoptotic pathways. Thus, drug resistance could develop if cancer cells come into contact with the ECM.
Through altering and manipulating the ECM or microenvironment, it is possible to overcome drug resistance (Gopal et al., 2017). In cancers such as breast, esophageal, and lung carcinomas, there is significant infiltration by the stroma (Muranen et al., 2012). Several ECM proteins such as fibronectin, collagen type I, and laminin have been shown to be overexpressed in many cancers (Dzobo et al., 2016; Senthebane et al., 2017; Yap et al., 2012). Ligation of the same ECM proteins to their respective integrins has been shown to activate survival pathways, resulting in loss of efficacy of drugs such as cisplatin and doxorubicin (Sansing et al., 2011). Inhibitors of integrins are under development and these target the interaction between cancer cells and stromal components (Eberle et al., 2011).
Another important point to consider is the heterogeneity of cells associated with cancer. For example, CAFs are known to be distinct from those associated with normal tissues (Attieh et al., 2017; Dourado et al., 2017; New et al., 2017). For cancer-associated fibroblasts, two distinct groups appear to be present (Ohlund et al., 2017), those close to cancer cells interact directly and those distant from cancer cells interact through paracrine mechanisms (Ohlund et al., 2017). In addition, adipose and bone marrow-derived cells such as mesenchymal stem cells (MSCs) can also infiltrate tumors (Senthebane et al., 2017).
Although complex, it is imperative that we understand this complexity to achieve cure in cancer patients. The ECM and other stromal components can also form a physical barrier, limiting drugs from reaching cancer cells (Attieh et al., 2017; Senthebane et al., 2017). Altering or degrading the ECM is a plausible option, although it can also create “highways” through which cancer cells can metastasize. The tumor stroma also exerts a selection pressure on the cancer cell subclones present within the solid tumor, shaping the final genetic architecture of a tumor and therefore sensitivity to drugs (Saunders et al., 2012).
Our data show that WHCO1 esophageal cancer cells cocultured with MSCs are more resistant to paclitaxel and cisplatin than cancer cells not cocultured with MSCs (Senthebane et al., 2017). Another example of the interaction between cancer cells and stromal cells is the relationship between breast cancer cells and osteoclasts (Kaplan et al., 2006). Primary breast cancer cells are involved in the maturation of osteoclasts through the release of growth factors (Guise et al., 2006). The matured osteoclasts release growth factors such as TGF-B1, known to be involved in the homing of breast cancer cells to the bone and the promotion of breast cancer cell growth in the bone (Guise et al., 2006). Removal of the osteoclasts has been shown to increase patients' survival (Guise et al., 2006; Kaplan et al., 2006). This classical case illustrates the need to understand the complex relationship between the tumor and its microenvironment to achieve successful cure for cancer.
Metastatic cancer cells have also been shown to be genetically distinct from those at the primary tumor site; thus, the stroma at the metastatic site and the presence of distinct cancer cell subpopulation determine the chemosensitivities of metastatic and secondary tumors (Wu et al., 2012). Macrophages within the tumor stroma are also different (DeNardo et al., 2011). Tumor-suppressive M1 macrophages are known to respond well to chemotherapy as opposed to protumorigenic M2 macrophages (Bellora et al., 2017; Zhu et al., 2017). The tumor-suppressive activity of lymphocytes is also affected by the presence of tumor-associated macrophages (Burmeister et al., 2017). The advantage of using next-generation sequencing techniques to study cancer-stromal interactions is that they are able to analyze mixtures of genomes in samples made up of both tumor and stromal cells (McGranahan and Swanton, 2017).
Tumor Cell Plasticity
Cancer cell plasticity is the ability of a cell to reversibly modify its behavior. This modification is different from the usual differentiation of the same cell (Castilho et al., 2017; Martinez-Marti et al., 2017). Cancer cells display significant plasticity, which extends to how it responds to the presence of drugs (Martinez-Marti et al., 2017). Changes within the tumor cell or its microenvironment play key roles in plasticity of cancer cells. Any modification of a cancer cell behavior may end up as a change in the cancer cell's response to drugs. Examples of cell plasticity include epithelial to mesenchymal transition (EMT) and mesenchymal to epithelial (MET) processes (Nieto, 2011). The mesenchymal phenotype is associated with chemoresistance and stem cell-like behavior in lung and pancreatic cancer (Cheng et al., 2007).
Recent data indicate that cancer cells that have undergone mesenchymal transition show resistance to targeted therapy and to drugs such as paclitaxel (Cheng et al., 2007). A recent study by Gupta et al. showed that different cancer cell subpopulations sorted from breast cancer cells can differentiate into all three lineages in the same way as in unsorted cells (Gupta et al., 2011). Thus, mature cancer cells in the different subpopulations are plastic and can change their phenotypes.
Heritable changes in gene expression with no changes to the DNA sequence are termed epigenetic modifications. Epigenetic alterations, including DNA methylation and histone deacetylation, are known to occur before and during tumor initiation, and indeed throughout the process of carcinogenesis (Avgustinova and Benitah, 2016; Bodenstine et al., 2016; Eun et al., 2017; Iwadate, 2017; Vizoso and Esteller, 2015). Most epigenetic changes are inherited during cell division and play key roles during cellular differentiation (Iwadate, 2017; Rodriguez-Ubreva et al., 2017; Sikandar et al., 2017; Vizoso and Esteller, 2015; Volkel et al., 2015).
Functionally, modifications to DNA, histone, and nucleosomes can be similar to DNA mutations in terms of gene expression. Cancer cells with epigenetic modifications can switch between biological states as the modifications are reversible (Caiado et al., 2016). Altered DNA methylation, usually at CpG islands, has been shown to be a characteristic of many cancers, including esophageal, head and neck cancers (Castilho et al., 2017; Eun et al., 2017; Feinberg et al., 2016; Garg, 2017; Iwadate, 2017). DNA methylation can result in loss of gene expression and when the affected genes are tumor suppressor genes, this can promote carcinogenesis. DNA methylation patterns in cancer cells within a single tumor are not uniform. This results in cancer cells with heterogeneous DNA methylation patterns and thus gene expression. This differential expression of genes mediated by DNA methylation can result in some cancer cells outcompeting others.
In tumors, the reversibility of epigenetic changes allows for cancer cells within a single tumor and in different tumors to show gene expression heterogeneity. A study by Landau et al. (2014) showed random disordered methylation of cancer cells within individual primary chronic lymphocytic leukemia samples (Swanton and Beck, 2014). This random disordered methylation patterns were associated with transcriptional variation between cancer cells and poor prognosis in patients. Thus, random disordered methylation patterns contribute toward genetic instability and increase intratumor heterogeneity. In another study by Mazor et al. (2015, 2016), loss of DNA methylation appears to be associated with disease recurrence. Sharma et al. (2016) showed that drug-resistant cancer cells showed chromatin modifications with high expression of histone demethylase enzyme and its associated signaling (Caiado et al., 2016).
Breast cancer cells are known to differentially methylate the TGFBR2 promoter, resulting in a differential TGF pathway activity (Melo et al., 2013; Shipitsin et al., 2007). Methylation of genes such as the Wnt can also result in cancer cells with increased tumorigenic properties (de Sousa et al., 2011). Other studies have shown that breast cancer cells are known to reversibly transition between epithelial and mesenchymal states, with those in mesenchymal state being more tumorigenic (Mani et al., 2007; Meacham and Morrison, 2013). Melanoma cells reversibly switch on and off the JARID1B histone deacetylase enzyme and melanoma cells expressing JAR1D1B can sustain tumor growth (Roesch et al., 2008, 2010, 2013).
Chemotherapeutic drug resistance is another cancer cell property that is reversible. For example, resistant cancer cells can give rise to drug-sensitive daughter cells and vice versa (Kemper et al., 2014; Schwitalla, 2014; Varga et al., 2014; Ye and Weinberg, 2015). Most epigenetic changes are reversible, making the enzymes responsible for these epigenetic changes an appealing option as drug targets. Several drugs targeting epigenetic changes such as DNA methylation and histone deacetylation have been approved by the FDA to date (Verbrugge et al., 2011). Tumor cell plasticity is also displayed as morphological heterogeneity. Cancer cells within a tumor might exist in different forms from rounded cells to fibroblastic forms. The changes in shape are associated with the activation of several GTPases such as Rac and Rho (Ruiz-Ontanon et al., 2013; Sanz-Moreno et al., 2008).
Impact of Intratumor Heterogeneity on Targeted Therapy
Impressive success in cancer treatment from the use of targeted therapy significantly improved the survival of cancer patients (Haber et al., 2011; Turner and Reis-Filho, 2012). Results, however, are dependent on the type and stage of tumor. Tumor heterogeneity has enormous implications for any therapy in cancer treatment. Targeted therapy has seen patients' survival improve as it focuses on certain pathways or mechanisms critical to cancer development (Carneiro et al., 2015; Zhang et al., 2016a). This is true especially if the cancer is detected early.
Advanced cancers tend to have more cancer cell subpopulations and therefore more molecularly distinct clones, making targeted therapy of limited benefit (Carneiro et al., 2015; Zhang et al., 2016a). Targeting ubiquitous driver events common to all tumor sites and, therefore, representing the trunk of the tumor may provide a better approach for targeting and the identification of potential biomarkers (Fig. 4) (Fisher et al., 2013; Gerlinger et al., 2014a, 2015; Yap et al., 2012).
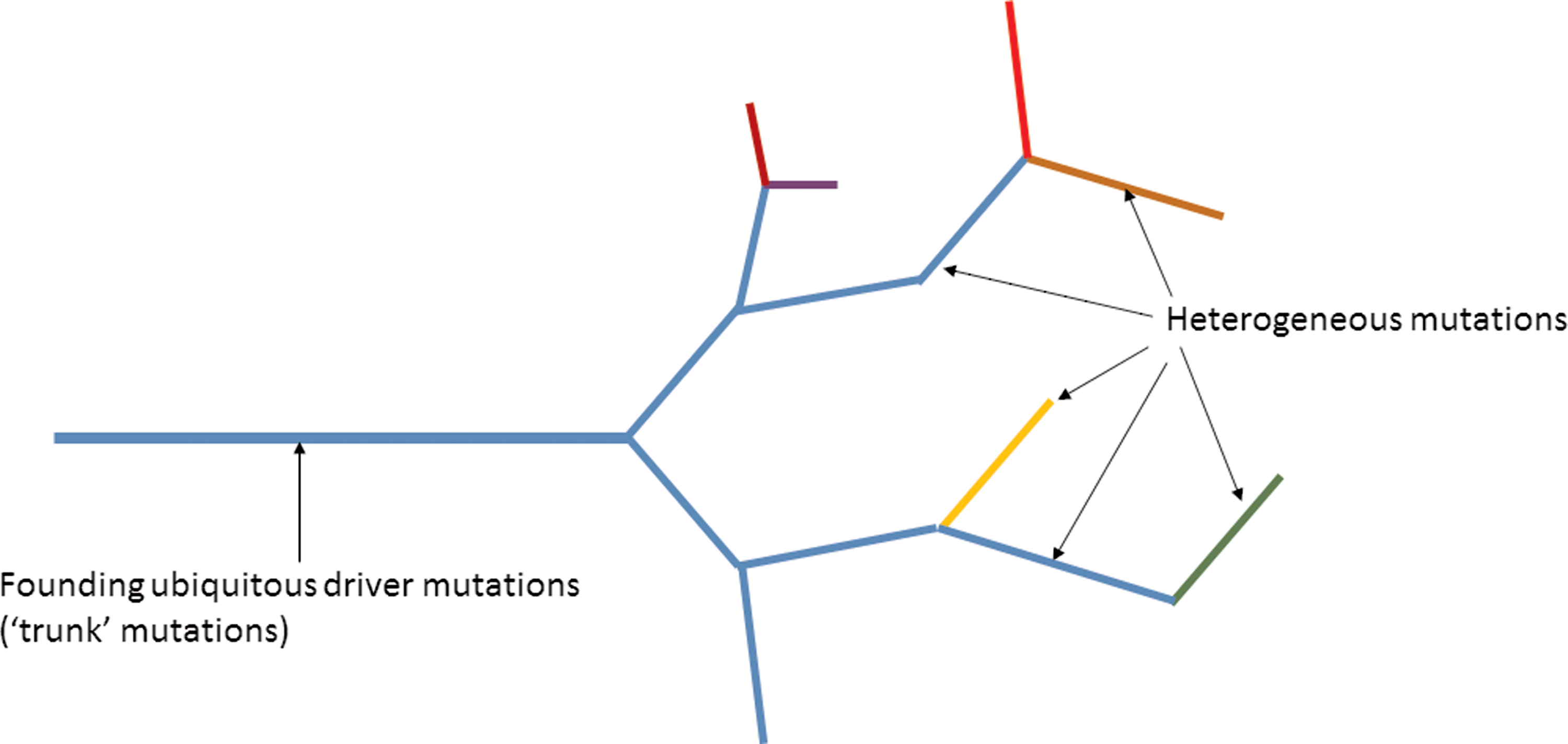
Therapeutic targeting of founding driver mutations within the “trunk” of a cancer and not the passenger mutations found in the branches of the tumor is an appealing strategy. The founding driver mutations are found in the trunk of the tumor and these are present in all the subclones. Heterogeneous mutations are found within the branches of the tumor and are found in different regions of the tumor. Targeting mutations found in all tumor cells therefore is a better strategy to eradicate cancer than targeting mutations within a fraction of tumor cells.
Tumors exhibiting many branches are difficult to treat due to the potential presence of subclones that are resistant to therapy (Cooke et al., 2011; Fisher et al., 2013; Turner and Reis-Filho, 2012; Yap et al., 2012). Therapy may actually apply selective pressure favoring the resistant subclone, resulting in the generation of new dominant clones after therapy (Fig. 5) (Ding et al., 2012; Johnson et al., 2014; Turner and Reis-Filho, 2012). In addition, microenvironmental factors and treatment selection pressure may act on the present cancer cell subclones, influencing their behaviors as drivers and passenger events (Fisher et al., 2013; Wu et al., 2012; Yap et al., 2012).
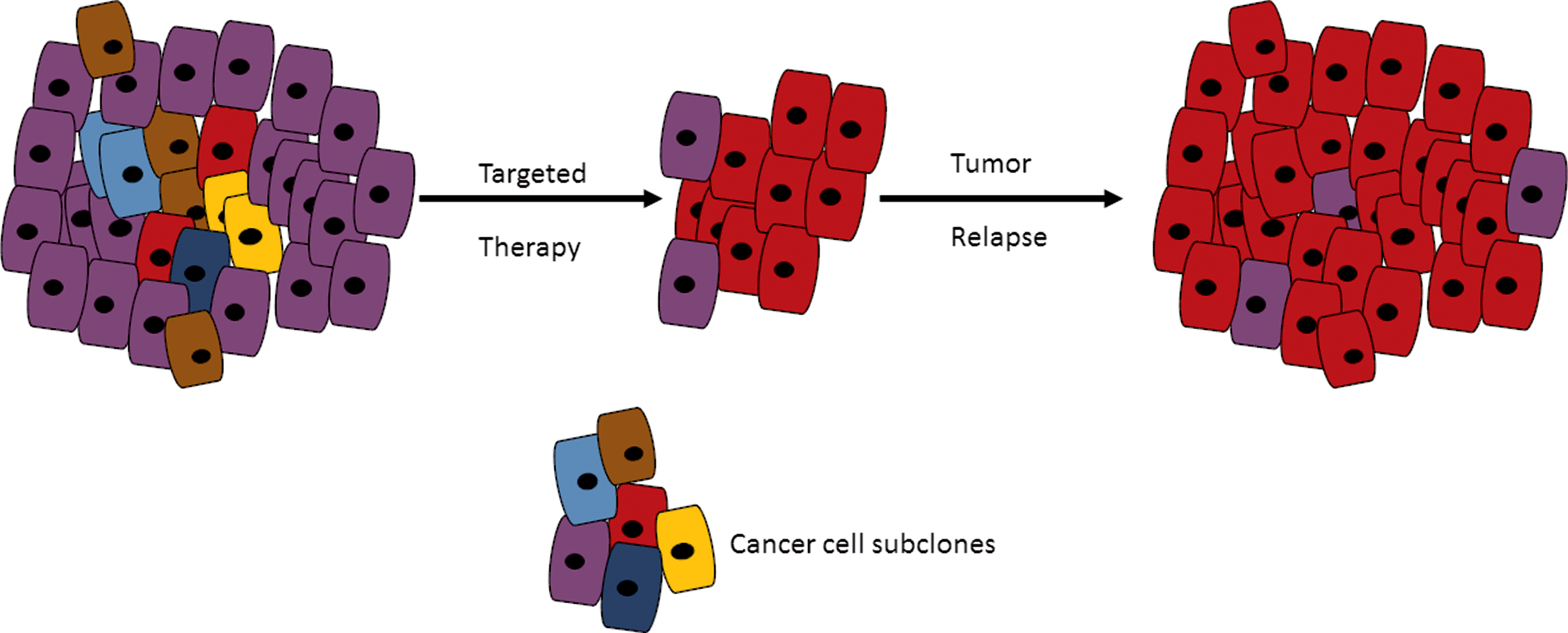
The influence of intratumor heterogeneity on tumor cell response to targeted therapy. Targeted therapy can result in a resistant subclone, occurring at low frequency in the solid tumor, to become the dominant clone after treatment. The implication of such a scenario is that all subclones must be identified in a tumor during biomarker discovery.
Indeed, it has been shown that driver events may require the presence of passenger events to continue in tumors. Biomarker discovery will have to content with the identification of even low-frequency subclones in solid tumors as these may confer drug resistance. Combination therapy is a better option to take care of most of the subclones present in tumors. Opportunities do exist before clonal expansion to treat the tumor by targeting the driver mutation (Gerlinger and Swanton, 2010). Importantly, oncologists must acknowledge that chemoresistance is a result of factors beyond just tumor heterogeneity. Chemoresistance can be caused by microenvironmental factors as well as nonmutational and epigenetic mechanisms (Lackner et al., 2012; Senthebane et al., 2017; Zhu et al., 2017).
Personalized medicine employing “omics” techniques such as next-generation sequencing as stratification tools does not consider the frequencies of subclones (McGranahan and Swanton, 2015). Several regimes targeting, for example, the PI3K pathway, do so against evidence showing that several members of this pathway are subclonal in many cancers (Basset and Guillermet-Guibert, 2017; Katoh and Katoh, 2017; Mann et al., 2016). It is imperative that we understand the complex nature of the interactions between subclones in cancers for clinical decisions as the targeting of dominant driver mutations is no longer sufficient (Yap et al., 2012).
It is also important to understand subclonal dynamics during treatment and posttherapy. Therapy, if continued in the presence of a resistant subclone, can actually result in the acceleration of tumor progression or the activation of survival pathways in cancer cells. Some subclones that are barely detectable at presentation can drive resistance to therapy. Thus, combinatorial therapy strategies, including the use of both chemotherapy and immunotherapy, may be the only option during treatment to counteract the existence of resistant subclones present in a tumor (Ding et al., 2012). Attempts at treating all subclones in a solid tumor and the use of combination therapy, however, bring issues such as overtreatment resulting in toxicity and economics into play, and these may limit the utility of “omics” techniques and combination therapy (Turner and Reis-Filho, 2012).
Targeted therapy is reliant on predictive biomarkers that can predict the response of tumors to drugs. Drug resistance, however, is multifactorial and as such, it is challenging to predict drug response based on just biomarkers. This is true since tumor heterogeneity and sampling bias can affect the biomarkers discovered or identified. Low frequency occurring clones may be considered insignificant during biomarker discovery, but may determine drug resistance. Currently, biomarkers obtained using cancer cells are obtained from the primary tumor direct targeted therapy (Punt et al., 2017; Turner and Reis-Filho, 2012; Zhang et al., 2016a).
To predict the therapeutic response, almost all clones within a tumor must be studied, especially the low-frequency clones. Results from deep sequencing and treatment strategies also appear to show that highly heterogeneous tumors are more likely to become drug resistant than less heterogeneous tumors (Ellis et al., 2012; Lee et al., 2011). Conventional cancer treatments such as chemotherapy and surgery can aid targeted therapies through tumor debulking (Turner and Reis-Filho, 2012).
Impact of Intratumor Heterogeneity on Cancer Drug Development
The development of cancer therapeutics takes years and is an expensive process (Stratton et al., 2009; Yap et al., 2010, 2012). Currently, the cost of drug discovery is estimated to be in excess of $1.0 billion dollars (Paul et al., 2010). The estimated number of preclinical programs needed to produce just one drug is upward of 30 and this is beside the fact that most drug candidates fail in clinical trials (Taube et al., 2009; Yap et al., 2010). Better models to evaluate anticancer drug candidates must involve pharmacodynamic, pharmacokinetic, and pharmacogenomic biomarkers.
In addition, the regulatory process is such a quagmire. Due to patient-to-patient differences in the mutational drivers of cancer and the existence of intratumor heterogeneity, drug discovery processes and clinical trials are insufficient in predicting patients' response to most drugs. Targeted therapy, the basis of personalized medicine, promises to be more effective at eradicating cancer as it targets specific mutations (Yap et al., 2010). Interpatient differences have a profound effect on pharmacodynamics and pharmacokinetics and these influence drug responses and toxicities. With respect to clinical trials, there has been a shift from treating all patients randomly to strategies taking into account a patient's molecular profile and then administration of appropriate therapy (Baiu et al., 2017; Eichenauer and Engert, 2017; Groisberg et al., 2017; Losanno and Gridelli, 2017).
The use of humanized mice models has revolutionized clinical trials in that mice that are genetically engineered to recapitulate the human cancer are now being used (Maugeri and Blasimme, 2011). By better reflecting the human disease, these models are churning out very useful data before the start of the human trials. In addition, certain experiments not ethically suitable for humans can now be tried in humanized mice models before their trial in humans. For personalized medicine to be successful, molecular profiling of tumors in real time during and after treatment and identification of key molecular drivers are important.
The availability of gene alteration screening platforms allows for the manipulation of several mutations at the same time during treatment to guide chemotherapeutic drug choices for particular patients (Andrews et al., 2010; Gao et al., 2013; Nakagawa et al., 2015). The use of a single patient limits the amount of pharmacodynamic and pharmacokinetic data obtained from such studies. Combination therapies, including well-characterized targeted therapy, must form new cocktails of different strategies used in the treatment of cancers. Key to the success of modern cancer therapy is the continual measurement of specific biomarkers during treatment and any changes brought about by the therapy itself.
Clinical Management of Intratumor Heterogeneity
It is important to develop methods that allow estimation of the potential contribution of cancer cell-stroma interaction, plasticity, and specific mutation to drug resistance. It has been reported that within a tumor, the fraction of mutation similarity is around 5% (Sharma et al., 2016; Taylor et al., 2011). The current best method to estimate intratumor heterogeneity is single-cell sequencing and only with such information can targeted therapy succeed. Targeting multiple targets from several cancer cell subpopulations may result in good clinical response. Major barriers to curative therapies remain, with cancer cell plasticity and genome instability as the main culprits (Saunders et al., 2012). Modifiers of cancer cell plasticity are definitely a good option to consider. Small molecules and proteins that can either inhibit cellular transition or inhibit the acquisition of the chemoresistant phenotype come into mind.
Experimental model systems that capture the complexity of solid tumors, in terms of genetic subpopulations, cancer cell-stroma interactions, plasticity, and stochastic processes, must also be developed (Wang et al., 2017b; Zhao et al., 2017). These models must be able to capture the intratumor heterogeneity exhibited in solid tumors. Combining both targeted and nontargeted therapies might also reduce the cancer burden. In addition, alteration or modification of the tumor stroma can be complementary to both targeted and nontargeted therapies (Bahrami et al., 2017; Kota et al., 2017; Majety et al., 2017; Wong et al., 2017). The development of system biology-based platforms that can track changing tumor heterogeneity during treatment and predict drug sensitivity is paramount. To be able to do this requires personalized medicine to be cost-effective and accessible to all patients.
Our deeper understanding of carcinogenesis will have to precede any novel diagnostic and therapeutic advancement. The discovery of new biomarkers and treatment strategies is borne out of the realization that chemoresistance is an enormous problem in oncology. New strategies need to focus on common features of clones such as controlling tumor growth and migration, and not necessarily eliminating every cancer cell as this has a tendency for selecting for resistant subclones (Aiello et al., 2016; Basset and Guillermet-Guibert, 2017; Katoh and Katoh, 2017; Wang et al., 2017a). The dosages of therapy may also be changed as the tumor responds (Gatenby et al., 2009). Thus, tumor-sensitive subclones can be maintained within the tumor, so that there is no selection of resistant subclones (Duan et al., 2017). These new innovative approaches are still in the development stages.
Overall, our tools for detecting and quantifying tumor heterogeneity must be sharpened to be able to measure the interplay between subclones within tumors. Intratumor heterogeneity brings another level of complexity to the development of diagnostics and the use of biomarkers. Driver genes and therefore driver mutations may increase as the tumor becomes more advanced. This will require sequencing several regions of the same tumor and the development of technologies to measure circulating tumor cells. Circulating tumor cells act as a readily available “liquid biopsy” and are a powerful tool used in noninvasive strategies of monitoring tumor dynamics in patients (Hodgkinson et al., 2014; Rothwell et al., 2016). Noninvasive strategies employing imaging to quantify tumor heterogeneity, and track disease progression and drug response must be developed (Gatenby et al., 2013).
Screening approaches to allow for the detection of cancer disease at early stages when tumor size and clonal diversity are still low are important. Many driver mutations may not be pathogenic at all in some individuals, with some mutations being detected in older individuals without the presence of disease (Busque et al., 2012). It appears certain events such as chromosomal rearrangements and doublings are necessary events before the development of disease in colon and head and neck cancers (McGranahan and Swanton, 2015).
There is need for bigger studies involving many patients, including postmortem analyses to study cancer subclones over time. Currently, there are little data on how tumor heterogeneity affects management of patients in clinics (Arvold et al., 2016; Buckner et al., 2017; Wright et al., 2017). A good alternative source to gain more knowledge about cancer molecular signature is to use blood (Alizadeh et al., 2015). With the use of blood, there is no need to obtain a biopsy sample from the tumor itself (Chen et al., 2017; Wu et al., 2015). Circulating tumor cells and cell-free DNA have shown that this method provides powerful information on cancer, especially of the breast, colon, and lung (Gallo et al., 2017). Tumor heterogeneity is likely to continue influencing several aspects of cancer research, the development of techniques to study tumor biology and how patients are treated.
Exploiting Tumor Heterogeneity
How both intratumor and intertumor heterogeneity affect tumor evolution and progression is only being elucidated now (Alizadeh et al., 2015; Burrell and Swanton, 2014b; Ding et al., 2012; Gerlinger et al., 2015). Scientists have been amazed at the extent of tumor heterogeneity observed even in a single tumor, let alone tumors from different patients. The source of heterogeneity and the extent to which heterogeneity affects different cancers are some of the confounding issues under investigation. In addition, some of the current questions include how tumor heterogeneity affects drug design and therapeutic strategies adopted for different patients. Tumor heterogeneity is the result of interplay between cancer cell genome, epigenome, protein expression, and the microenvironment (Burrell and Swanton, 2014b). Functional screening must be done together with measurement of epigenetic, signaling, transcriptional, and metabolic alterations to reveal the source of phenotypes driving the process of carcinogenesis.
Although tumor heterogeneity is the trait or behavior that gives tumors the ability to overcome selection pressures, it can also present an opportunity that can be taken advantage of therapeutically. Treatment of cancer cell subclones can be done in such a way as to use the initial treatment as a selection tool, so that the remaining subclones can be eradicated by a second drug or therapy (Schmoll et al., 2012; Wilke et al., 2014). Recent data suggest that excessive genomic instability may result in tumor collapse, suggesting that drugs that induce further genomic instability could result in tumor eradication (Fisher et al., 2013).
It is an enormous challenge to determine the role tumor heterogeneity plays in tumor progression and metastasis as most tumor models are not representative of the physical environment under which tumors become heterogeneous in vivo (Alizadeh et al., 2015; Shackleton et al., 2009). Even humanized mouse models, genetically engineered to recapitulate human tumors, tend to be homogenous and driven by fewer genetic aberrations (Alizadeh et al., 2015). In addition, several crucial aspects of tumor biology such as tumor load and the ability to metastasize are not represented properly in most models (Alizadeh et al., 2015; Coughlan et al., 2012). Genome editing techniques such as CRISPR/Cas9 may be able to recapitulate the necessary complexity needed in these tumors to recapitulate human tumor heterogeneity (Hamacher and Bauer, 2017; Song et al., 2017; Wang et al., 2017c).
In addition, humanized mouse models lack the necessary microenvironmental and immune components that affect clonal selection in humans. Only when we can fully incorporate these important features of human tumors into these models can we be able to really understand human tumor evolution and human tumor heterogeneity. Alternative strategies involve the use of tumor slice cultures. As with biopsy samples, tumor slice cultures only present a fraction of the heterogeneity present in any tumor (Finnberg et al., 2017; Grabiec et al., 2017). The culturing of a tumor slice in itself may add external factors that can alter the heterogeneity within the slice.
Organoids are only useful in recapitulating the process of tumorigenesis in human cells (Devarasetty et al., 2017; Diermeier and Spector, 2017; Senthebane et al., 2017; Yan et al., 2017). The expansion of cancer models such as organoid cultures is especially important in order for the promise of personalized medicine in cancer treatment to be fulfilled. To achieve this, the coming together of several international researchers, in addition to freely sharing of data by researchers, is very important. Computational models can also create formulas and hypotheses that can be tried and tested in the laboratory (Alizadeh et al., 2015). As all models are imperfect, the gold standard remains testing tumor heterogeneity in patients.
The interaction between the genome, epigenome, transcriptome, proteome, and the microenvironment determines the phenotype of any cell (Alizadeh et al., 2015). It is possible that genetically distinct cells could have the same phenotype, while genetically identical cells could have a different phenotype due to the influence of microenvironment and epigenetic factors (Alizadeh et al., 2015; Dzobo et al., 2016; Senthebane et al., 2017). During the process of carcinogenesis, changes in epigenetic mechanisms can eventually cause genetic mutations, while some genetic mutations in epigenetic regulator genes can result in epigenetic changes (Brait et al., 2009; Gelato et al., 2016; Klosin et al., 2017; Li et al., 2017).
Epigenetic modifications or changes play a role in the eventual phenotype of a cancer cell and might be useful in indicating the evolutionary history of a cancer (Brait et al., 2009; Gelato et al., 2016). Some epigenetic modifications might be useful in predicting how tumors respond to chemotherapeutic agents (Castilho et al., 2017; Chen et al., 2014). Due to the plasticity of epigenetic modifications in cancer, these may be important in understanding processes such as drug resistance, EMT and metastasis (Slattery et al., 2008). The influence of the microenvironment is understood only after a detailed analysis of the genome, epigenome, transcriptome, proteome, infiltrating immune cells, fibroblasts, and metabolites.
Conclusions
This review evaluated the sources of tumor heterogeneity and the implications on cancer drug design and development. The phenotypic properties of cancer cells are a result of not only genetic and epigenetic factors but also microenvironmental influence. This presents a major obstacle to drug development and eventual cancer eradication. Next-generation sequencing techniques and single cell analysis have equivocally shown that tumors consist of distinct clones differing in all manners imaginable. This coexistence of many clones within a tumor has profound implications for diagnosis and therapeutic response to drugs. The presence of these distinct subclones within solid tumors might explain the failure of targeted therapies.
Currently, it is possible to use latest technologies such as microarray analysis to predict therapy responsiveness of patients. For example, gene expression signature associated with the treatment of breast cancer with tamoxifen and combination therapy for colon cancer has been developed and used successfully (Caiado et al., 2016; Newcomb et al., 1999; Notas et al., 2015; Ojo et al., 2015; Singh et al., 2011; Zhang et al., 2013; Ziv et al., 1994, 1996).
Tumor heterogeneity has resulted in combination therapy being the only plausible treatment option and does affect the practise of personalized medicine. Multiple biopsy samples must be taken from different regions of the tumor and during treatment to understand the evolution of the tumor over time. Targeting driver mutations within the trunk of the tumor is a promising strategy. Given that the extent of tumor heterogeneity observed in a tumor is associated with poor prognosis, this calls for the unbiased analysis of primary tumors.
Cutting edge techniques such as fluorescence-activated cell sorting of tumor clones and single cell analysis of tumor clones must be adopted, especially to resolve intratumor heterogeneity. Strategies such as the use of circulating tumor DNA analysis might help avoid the ethical issue of taking multiple biopsy samples. Tumor heterogeneity must be taken into account during the design of tumor models and drug development. This is crucial since drug resistance mechanisms can be of nongenetic origin. In this regard, the development of epigenetic inhibitors and modulators such as histone deacetylase inhibitors and other small molecules as anticancer drugs must be supported.
We argue that innovative cancer drug design and development must be of combinatorial nature and not focus only on cancer cell properties such as proliferation. Such drugs must target stromal components, and tumor cell plasticity and migration as these are common to all cancer cells. The truth is that the “one size fits all” strategy used in cancer treatment is no longer acceptable. Delineating the complexity of tumors, especially intratumor and intertumor heterogeneity, although difficult, will allow the identification of critical interactions and signaling pathways needed for the design of effective therapeutic interventions, leading to the eradication of cancer.
Definitions
Footnotes
Acknowledgments
This work was supported by the National Research Foundation (NRF) of South Africa, the International Centre for Genetic Engineering and Biotechnology (ICGEB), the South African Medical Research Council, and the University of Cape Town. The funders played no role in the preparation of the article. We would like to thank Professor Vural Özdemir for constructive editorial feedback.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.