
Abstract
Abstract
Interindividual and interethnic differences in response to antiretroviral drugs (ARVs) are influenced by genetic variation. The few genomic studies conducted among African-Americans and African ethnic groups do not reflect the extensive genetic diversity within African populations. ARVs are widely used in Africa. Therefore, genomic characterization of African populations is required before genotype-guided dosing becomes possible. The aim of this study was to determine and report on the frequency of genetic variants in genes implicated in metabolism and transport of ARVs in South African populations. The study comprised 48 self-reported South African Colored (SAC) and 296 self-reported Black African (BA) individuals. Allele and genotype frequency distributions for 93 variants contributing to metabolism and transport of ARVs were compared between groups, and other global populations. Fifty-three variants had significant differences in allele and genotype frequencies when comparing SAC and BA groups. Thirteen of these have strong clinical annotations, affecting efavirenz and tenofovir pharmacokinetics. This study provides a summary of the genetic variation within genes implicated in metabolism and transport of ARVs in indigenous South African populations. The observed differences between indigenous population groups, and between these groups and global populations, demonstrate that data generated from specific African populations cannot be used to infer genetic diversity within other populations on the continent. These results highlight the need for comprehensive characterization of genetic variation within indigenous African populations, and the clinical utility of these variants in ARV dosing for global precision medicine. Population pharmacogenetics is a nascent field of global health and warrants further research and education.
Introduction
T
In South Africa, the recommended initial regimen for treatment-naive adult patients entails administration of a single tablet containing two nucleoside reverse transcriptase inhibitors, including tenofovir (TDF) and either emtricitabine (FTC) or lamivudine (3TC), and one non-nucleoside reverse transcriptase inhibitor, such as efavirenz (EFV), dolutegravir (DTG), or rilpivirine (RPV) (Meintjes et al., 2017).
Despite the efficacy of these ARVs, some patients experience adverse effects and poor treatment outcomes when treated with standardized doses (Apostolova et al., 2015; Eron et al., 1995; Rajman et al., 2017; Ustianowski and Arends, 2015). The occurrence of adverse effects and variable treatment outcomes are often a result of interindividual variability in plasma concentrations, especially for ARVs with narrow therapeutic ranges such as EFV (Zanger and Schwab, 2013). Drug response is multifactorial and some of the aspects associated with interpatient drug concentration variability include demographical, environmental, behavioral, and genetic factors (Cressey and Lallemant, 2007).
Genetic variants in genes encoding ARV-metabolizing enzymes and transporters may affect the functionality and/or expression of these proteins, thereby altering pharmacokinetic parameters and subsequently ARV plasma concentrations (Aceti et al., 2015; Dalal et al., 2015). Further complicating the pharmacokinetics of some ARVs, such as EFV, is the mechanism of autoinduction (Habtewold et al., 2011; Ke et al., 2016; Ngaimisi et al., 2010, 2011; Yanakakis and Bumpus, 2012). This mechanism may be concentration and/or time dependent and may also be influenced by both environmental and genetic factors (Habtewold et al., 2011; Ngaimisi et al., 2011). Alterations in pharmacokinetics due to these factors likely impact long-term exposure and the extent of suppression of viral replication (Aceti et al., 2015; Dalal et al., 2015).
Although genetic variation is known to influence the metabolism and transport of ARVs, pharmacogenetic studies have mainly been performed in Caucasian and Asian populations (Nelson et al., 2013; Rajman et al., 2017; The 1000 Genomes Project Consortium, 2015). Data generated from African-American populations are sometimes used to infer genetic variation within populations on the African continent (Nelson et al., 2013; Rajman et al., 2017; The 1000 Genomes Project Consortium, 2015). In an attempt to address these concerns, (Rajman et al., 2017) a review on variation within the drug-metabolizing cytochrome P450 (CYP) gene family in African populations was conducted.
Furthermore, candidate gene studies investigating additional genes implicated in ARV metabolism and transport have shown extensive genetic diversity in various African cohorts (Čolić et al., 2015; Dandara et al., 2011; Drögemöller et al., 2010; Ikediobi et al., 2011; Mukonzo et al., 2014; Ngaimisi et al., 2013; Swart and Dandara, 2014; Swart et al., 2012a, 2012b, 2013; Wright et al., 2010). To date, few studies have attempted to compare the frequency of variants within genes known to affect metabolism and transport in indigenous Sub-Saharan African populations and these studies have focused on isolated ethnic groups and different genes or single nucleotide variants (SNV).
The aim of this study was to determine and contextualize the frequency of variants within genes implicated in metabolism and transport of ARVs in indigenous South African populations.
Materials and Methods
Study participants and sample collection
DNA samples of patients from two cohorts, recruited as part of our ongoing pharmacogenetics research, were obtained from our biorepositories. The first cohort comprised of 125 (77 Black African [Xhosa] and 48 South African Colored [SAC]), unrelated, female participants. All participants provided written informed consent (Röhrich et al., 2016). As previously described, DNA was extracted from saliva samples (22). Ethics approval was obtained from Stellenbosch University's Human Research Committee (N08/08/225).
The second cohort comprised 219 South African participants of Black African origin (101 Zulu, 45 Tswana, 42 Sotho, and 31 Xhosa). Clinical and demographic factors for this cohort have been described (Swart et al., 2013, 2015). A 5 mL whole blood sample was obtained from each subject using venipuncture, and used for plasma sample collection as well as DNA extraction as previously described (Swart and Dandara, 2014; Swart et al., 2013, 2015). All participants provided written informed consent and study approval was obtained from the University of Cape Town Health Science Faculty Research Ethics Committee, Cape Town, South Africa (HREC REF 103/2009) and the University of Witwatersrand Human Research Ethics Committee, Gauteng, South Africa (M080124).
Variant selection
The PharmGKB database (McDonagh et al., 2011) was searched using the terms “efavirenz,” “tenofovir,” “emtricitabine,” “lamivudine,” “dolutegravir,” and “rilpivirine” and all SNVs returned for each query were selected for genotyping. The specific high-throughput genotyping method employed did not allow for genotyping of all SNVs and a subset of SNVs were replaced with additional candidate SNVs (selected based on frequently being reported in pharmacogenomic literature) (Supplementary Table S1).
High-throughput genotyping
All samples were genotyped using the QuantStudio™ 12K Flex Real-Time PCR System (Applied Biosystems, CA) and a custom-designed microarray plate containing all prioritized SNVs (Supplementary Table S2). Duplicate SNV assays were included at random to serve as internal controls and to ensure genotyping accuracy.
Population structure and statistical analyses
Extensive genetic diversity exists in Sub-Saharan African populations partly because of geographic isolation, differences in cultural practices, conflicts, and colonization (Busby et al., 2016; Patin et al., 2017). As a result, genetic ancestry has been shown to be more informative than self-reported ancestry (Spector et al., 2016). Ethnicity was self-reported for all participants in this study, thus, principal component analysis (PCA) was performed to assess the reliability of this self-reporting. PCA was performed in R using the “stats” package (R Core Team, 2017). Chi-squared or Fisher's exact tests were performed to compare allele and genotype frequency distributions for SNVs of interest between groups. Bonferroni correction was used to correct for multiple testing and significance was set at p < 5 × 10−4 (0.05/93).
Results
PCA showed two clusters when all participants were included. Specifically, the self-reported SAC participants were shown to cluster separately from other participants (Fig. 1A). Further analyses of all other participants (self-reported Zulu, Tswana, Sotho, and Xhosa), excluding the self-reported SAC participants, showed no independent clustering (Fig. 1B). Based on the results of the PCA, genotype frequencies were reported separately for the SAC group (n = 48) and the combined BA grouping (n = 296).
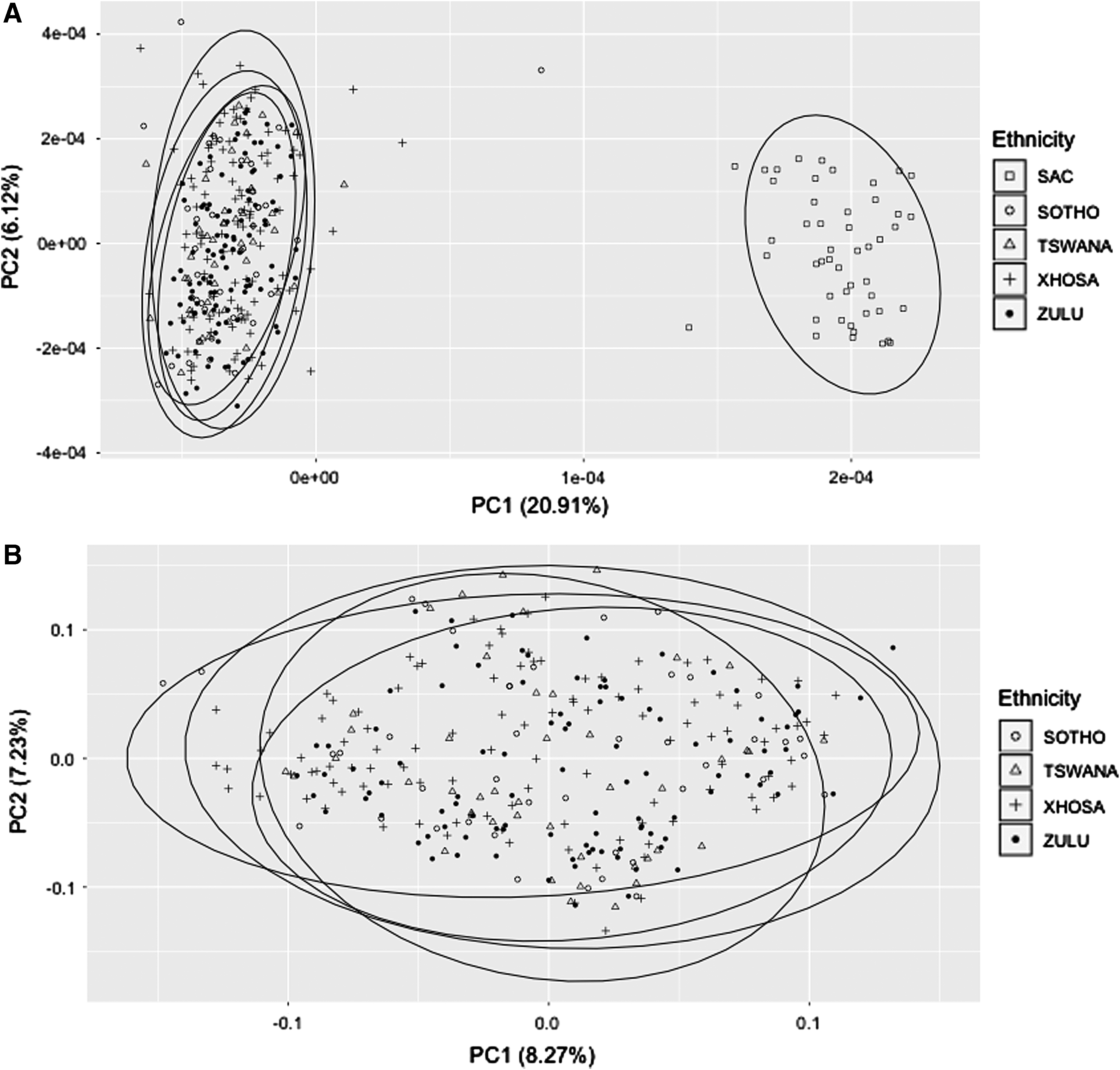
Performed population structure analyses.
Genotype and allele frequencies for the 93 SNVs included in this study are presented in Supplementary Tables S2 and S3. Among the SAC group, genotype frequencies for NR1I2 rs1054191 deviated from Hardy–Weinberg equilibrium (HWE) (p < 0.001), and genotype frequencies for ABCC2 rs7910642 deviated from HWE in the BA group (p < 0.001).
Genotype frequencies for the SNVs in the SAC and BA groups are compared with frequencies from the Utah Residents with Northern and Western European Ancestry (CEU), Americans of African Ancestry in South West USA (ASW), Yoruba in Ibadan, Nigeria (YRI), Luhya in Webuye, Kenya (LWK), and Han Chinese in Beijing, China (CHB) population groups from the HapMap3 project (International HapMap Consortium, 2003; International HapMap 3 Consortium et al., 2010) (Supplementary Table S2). Furthermore, allele frequencies are compared with the five super populations of the 1000 Genomes project (African [AFR], Admixed American [AMR], East Asian [EAS], European [EUR], and South Asian [SAS]) in Supplementary Table S3 (The 1000 Genomes Project Consortium, 2015).
Of the 93 SNVs investigated, a subset of 53 unique SNVs of interest were identified based on significantly different (p < 1 × 10−8) genotype (28 SNVs) and allele (30 SNVs) frequency distributions within the SAC and BA groups (Tables 1 and 2). Comparing genotype frequencies in the SAC and BA groups showed a distinct difference in genotype profiles for CYP3A5 rs15524 and NR1I2 rs3732360 (Table 1). For these two SNVs, the BA group had genotype frequencies more like the ASW, YRI, and LWK population groups, whereas the SAC group showed distinct frequencies which tended to align more with the CEU or CHB population groups. Particularly, 11 SNVs had greater than 20% differences in major genotype frequencies when comparing distributions between the SAC and BA groups (Table 1).
No. of participants for the SAC and BA groups is indicated in parentheses. The absence of HapMap3 genotype frequency data for the selected populations is indicated with a “—.” SNVs in bold typeset are present twice in the table.
SNVs that have been assigned a PharmGKB clinical annotation and level of evidence for effect on efavirenz and/or tenofovir.
ASW, Americans of African Ancestry in South West USA; BA, Black African; CEU, Utah Residents with Northern and Western European Ancestry; CHB, Han Chinese in Beijing, China; CYP, cytochrome P450; LWK, Luhya in Webuye, Kenya; SNV, single nucleotide variant; YRI, Yoruba in Ibadan, Nigeria.
No. of participants for the SAC and BA groups is indicated in parentheses.
SNVs that have been assigned a PharmGKB clinical annotation and level of evidence for effect on efavirenz and/or tenofovir.
AFR, African; Alt, 1000 Genomes alternate allele; AMR, Admixed American; EAS, East Asian; EUR, European; Ref, 1000 Genomes reference allele; SAS, South Asian.
When considering allele frequency distributions, 19 SNVs showed reversals of alternate allele between the SAC and BA groups (Table 2). Furthermore, 11 SNVs had a greater than 20% difference in alternate allele frequencies when comparing distributions between the SAC and BA groups (Table 2). We also report on 17 SNVs that were monomorphic (Table 1).
For EFV or TDF, 13 SNVs within the subset of interest had PharmGKB clinical annotations (McDonagh et al., 2011) with corresponding assigned levels of evidence (Whirl-Carrillo et al., 2012) (Tables 1 and 2). No SNVs within the subset of interest had PharmGKB clinical annotations for 3TC, FTC, DTG, or RVP.
The CYP2B6 rs3745274 (*9) SNV had the highest assigned levels of evidence for EFV dosage (1B) and toxicity (2A), followed by CYP2B6 rs4803419 (intronic variant) and ABCC4 rs1751034 with a level of evidence of 2B for EFV and TDF pharmacokinetics, respectively. The remaining six SNVs had levels of evidence less than or equal to three for EFV pharmacokinetics (CYP3A4 rs2740574 (*1B), CYP2A6 rs28399433 (*9), NR1I3 rs3003596, CYP3A5 rs776746 (*3), and CYP2B6 rs8192719); EFV toxicity (IL10 rs1800896 and HTR2A rs6313); TDF pharmacokinetics (ABCC4 rs3742106); or TDF toxicity (ABCC10 rs9349256 and OCRL rs757639).
Discussion
This study provides further evidence for extensive genetic diversity and significant genetic differences between indigenous South African populations (SAC and BA). The observed differences related to the studied pharmacogenetic targets suggest that responses to cART may differ among South African population groups, and this needs to be considered as we advance precision medicine.
The allele and genotype profiles for several SNVs within genes implicated in ARV metabolism and transport were markedly different between the SAC and BA groups, as well as when South African population groups are compared with other world population groups. PCA analysis, of the 93 SNVs investigated in this study, showed no distinct clustering between any of the Black African groups (Tswana, Sotho, Xhosa, and Zulu) investigated. This may be expected since the Black South African population groups all belong to the larger Bantu-speaking group of populations that is thought to have migrated from areas around west Africa and populated most of sub-Saharan Africa (Li et al., 2014).
This study highlights specific genetic differences within genes responsible for metabolism and transport of ARVs, and other medicines between the SAC and BA groups, which may be exploited for individualized genotype-guided ARV dosing. Differences in alternate allele and major genotype frequency distributions, were identified for 53 unique SNVs between the SAC and BA groups. Of these SNVs, 13 had corresponding PharmGKB clinical annotations (McDonagh et al., 2011) and assigned levels of evidence (Whirl-Carrillo et al., 2012); the majority of which were specific to EFV (CYP2B6 rs3745274 (*9), rs4803419 and rs8192719, CYP3A4 rs2740574 (*1B), CYP3A5 rs776746 (*3), HTR2A rs6313, IL10 rs1800896, and NR1I3 rs3003596); and four to TDF (ABCC4 rs1751034 and rs3742106, ABCC10 rs9349256 and OCRL rs757639).
CYP variation and EFV pharmacokinetics
The CYP2B6 rs3745274 (*9) SNV is known to affect EFV dosage and toxicity. Specifically, the T allele is associated with increased EFV plasma concentrations due to reduced EFV metabolism rates (Cusato et al., 2016; Hui et al., 2016; Mukonzo et al., 2013; Sinxadi et al., 2015; Swart et al., 2015). Similarly, the CYP2B6 rs8192719 T/T genotype is associated with increased plasma EFV concentrations (Sukasem et al., 2012). These SNVs had considerably higher minor genotype frequencies in the BA group (T/T: 17% and T/T: 16%) when compared with the SAC group (T/T: 4% and T/T: 4%), as well as the majority of other HapMap3 population groups investigated (Table 1; excluding the YRI HapMap3 population group).
In contrast, another CYP2B6 SNV, the intronic rs4803419, had a higher major C/C genotype frequency in the BA group (86%) when compared with the SAC group (58%). The reported C allele frequencies in the 1000 Genomes Project (The 1000 Genomes Project Consortium, 2015) in African and European superpopulations are 92% and 68%, respectively (Table 2). The C/C genotype is associated with reduced plasma concentrations of EFV (Holzinger et al., 2012; Sinxadi et al., 2015; Swart et al., 2015).
Similarly, the major genotypes for CYP3A4 rs2740574 (*1B) and CYP3A5 rs776746 (*3) were considerably more frequent in the BA group (rs2740574 C/C: 51% and rs776746 T/T: 73%) than with the SAC group (rs2740574 C/C: 31% and rs776746 T/T: 40%) (Table 1).
EFV plasma concentration is increased in individuals with the CYP3A4 rs2740574 C/C (Haas et al., 2004, 2005) and CYP3A5 rs776746 T/T (Haas et al., 2004) genotypes. Interestingly, CYP3A4 rs2740574 C allele frequency was lower in both the SAC (rs2740574 C: 52%) and BA (rs2740574 C: 44%) groups when compared with the AFR population (rs2740574 C: 77%), and higher when compared with the other 1000 Genomes populations (rs2740574 C; AMR: 11%, EAS: 0%, EUR: 3% and SAS: 4%) (Supplementary Table S3). Furthermore, the BA group (rs776746 T/T: 73%) showed similar CYP3A5 rs776746 genotype frequency distributions to the YRI (rs776746 T/T: 69%) and LWK (rs776746 T/T: 77%) populations (Table 1).
The CYP2A6 promoter SNV, rs28399433 (*9), was shown to be monomorphic (for the A/A genotype) in the SAC group, but polymorphic in the BA group. Although no HapMap3 genotype data were available for this SNV (Table 1), the reported global minor allele (C) frequency, according to The 1000 Genomes Project (The 1000 Genomes Project Consortium, 2015), is ∼12%. This SNV is implicated in altered EFV pharmacokinetics, with the A/A genotype associated with increased EFV metabolism and corresponding reduced EFV plasma concentrations (Haas et al., 2014; Leger et al., 2009; Sarfo et al., 2014; Soeria-Atmadja et al., 2017).
These results demonstrate that the allele and genotype frequency distributions of CYP SNVs known to influence EFV pharmacokinetics vary considerably between indigenous South African population groups (SAC vs. BA), and between South African and other populations. Moreover, these results emphasize the importance of pharmacogenetic studies in understudied, diverse, African populations. These data further suggest that optimal EFV dosage, based on CYP genetic markers, needs to be considered and tested individually for the SAC and BA groups and that the use of data from other African or global populations to infer EFV dosage is not recommended.
Non-CYP variation and EFV pharmacokinetics and response
Three non-CYP SNVs known to influence EFV pharmacokinetics and response were also shown to have distinct differences in allele and genotype distributions between the SAC and BA groups. The NR1I3 rs3003596 A/A genotype was more frequent in the BA group (rs3003596 A/A: 39%) than in the SAC group (rs3003596 A/A: 19%), and this genotype is associated with increased EFV plasma concentration (Swart et al., 2012b).
The other two SNVs were implicated in EFV response. The HTR2A rs6313 A allele is associated with reduced depression symptoms during EFV treatment (Sánchez Martín et al., 2013), whereas the IL10 rs1800896 C allele is associated with reduced EFV drug hypersensitivity (Rodrigues et al., 2014). For both of these SNVs, the alternate allele was shown to be higher in the BA group (rs6313 A: 47% and rs1800896 C: 67%) when compared with the SAC group (rs6313 A: 25% and rs1800896 C: 38%) (Table 2). These results further corroborate the importance of pharmacogenetic studies in underrepresented populations and population groups.
TDF pharmacokinetics and response
Four SNVs of interest (ABCC4 rs1751034 and rs3742106, ABCC10 rs9349256 and OCRL rs757639) had an identified clinical annotation (McDonagh et al., 2011) and corresponding assigned level of evidence (Whirl-Carrillo et al., 2012) specific to TDF toxicity. The ABCC4 rs1751034 T/T and rs3742106 A/A genotypes are associated with increased renal clearance (Kiser et al., 2008a, 2008b) and reduced plasma concentration of TDF (Rungtivasuwan et al., 2015), respectively, potentially resulting in reduced TDF-induced nephrotoxicity. The corresponding rs1751034 T and rs3742106 A alleles had higher frequency distributions in the SAC group (rs1751034 T: 75%, rs3742106 A: 66%), and all 1000 Genomes populations (Table 2), when compared with the BA group (rs1751034 T: 38%, rs3742106 A: 16%).
The ABCC10 rs9349256 G/G genotype is associated with increased risk for kidney tubular dysfunction (Pushpakom et al., 2011). This genotype had a higher frequency in the BA group (91%), with similar frequency to the YRI and LWK HapMap3 populations, when compared with the SAC group (65%) (Table 1). Furthermore, the frequency of this genotype in the BA (91%) and SAC group (65%) was higher than that reported in the CEU (30%) or CHB (18%) HapMap3 populations (Table 1) suggesting potential higher burden for kidney tubular dysfunction in indigenous South African populations.
Finally, the OCRL rs757639 C/C genotype was associated with increased creatine clearance in patients treated with TDF (Dahlin et al., 2015). The corresponding rs757639 C allele showed a lower frequency distribution in the BA group (rs757639 C: 12%) when compared with the SAC group (rs757639 C: 59%) and other 1000 Genomes populations (Table 2). These results highlight the differences in potential genetic burden for TDF toxicity between the indigenous South African populations, as well as between South African and other global populations. Furthermore, these results underscore the importance of pharmacogenetic studies in poorly represented and genetically diverse populations such as those indigenous to South Africa (SAC and BA groups).
RVP pharmacokinetics
Although no clinical annotations (McDonagh et al., 2011) and corresponding assigned levels of evidence (Whirl-Carrillo et al., 2012) for RVP were identified for the SNVs of interest, one SNV (CYP3A5 rs776746 - *3) was recently associated with variability in RVP clearance rates (Aouri et al., 2017). Specifically, individuals with the rs776746 C allele had reduced RVP clearance (Aouri et al., 2017). These results need to be replicated and the clinical utility of this association remains to be determined. It is interesting to note that the frequency of the rs776746 C/C genotype is considerably higher in the SAC group (23%), and CEU and CHB populations, than in the BA group (2%) or other African populations (YRI and LWK) (Table 1). Future studies are required to investigate how the high frequency of this genotype could alter the pharmacokinetics of RVP when administered to SAC, CEU, or CHB individuals.
In addition to the SNVs described above, several other genotype distribution differences between the SAC and BA groups were observed, including the reversal of major genotypes for CYP3A5 rs15524 and NR1I2 rs3732360 (Table 1), and the reversal of alternate alleles for several SNVs (Table 2). To date, no clinical annotations (McDonagh et al., 2011) and corresponding assigned levels of evidence (Whirl-Carrillo et al., 2012) specific to EFV, TDF, FTC, 3TC, DTG, or RVP have been described for these SNVs. However, genetic variants investigated in this study are of broader interest because of their presence in genes involved in metabolism and transport of many commonly used medications, not just ARVs. Further pharmacogenetic studies should focus on accessing the clinical utility of these SNVs in medication dosing, for ARV and other medications, within indigenous South African populations.
One limitation of this study is the small number of SAC participants. A larger cohort of SAC participants is required to validate these research findings and ensure the accuracy and generalizability of described genotype distributions. In addition, novel genetic variants are frequently identified among individuals within South African populations (Choudhury et al., 2017). Future studies should consider advanced genotyping methodologies, such as whole-exome, whole-gene, or targeted gene sequencing in larger cohorts to have a complete understanding of the genetic differences between indigenous groups of South Africa.
This study provides a comprehensive summary of the genetic variation within genes implicated in the metabolism and transport of ARVs, specifically EFV, TDF, FTC, 3TC, DTG, and RVP, in indigenous South African populations. The observed differences between South African population groups, and between these groups and global populations, demonstrate that data generated from specific African populations cannot be used to infer genetic diversity within other populations on the African continent. These results highlight the need to better characterize genetic variation within indigenous African and South African populations and to determine the clinical utility of these variants in ARV dosing to provide accurate pharmacogenetic interventions. Furthermore, although the focus of this study was ARVs, these findings may be beneficial to other pharmacogenetic studies in South African populations.
Footnotes
Acknowledgments
The work reported here was supported by the following grants provided by the South African National Research Foundation (NRF): K.O.C. was funded by the Scarce Skills Postdoctoral Fellowship (grant no. 96833), L.W. was funded by the Competitive Program for Rated Researchers (grant no. 93498) and the Bioinformatics and Functional Genomics Program (grant no. 93681). MS was funded by the University of Cape Town Carnegie Corporation Developing the Next Generation of Academics Program (infectious diseases research focus) Scholarship and the South African Medical Research Council. C.D. and L.W. received South African Medical Research Council Self-Initiated Research Grants. The opinions expressed, and conclusions arrived at are those of the authors and are not necessarily attributed to the funding sources.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.