
Abstract
Abstract
Coumarins such as warfarin are prescribed for prevention and treatment of thromboembolic disorders. Warfarin remains the most widely prescribed and an anticoagulant of choice in Africa. Warfarin use is, however, limited by interindividual variability in pharmacokinetics and a narrow therapeutic index. The difference in patients' pharmacodynamic responses to warfarin has been attributed to genetic variation in warfarin metabolism and molecular targets (e.g., CYP2C9 and VKORC1) and host–environment interactions. This expert review offers a synthesis of human genetics studies in Africans with respect to pharmacogenetics-informed warfarin dosing. We identify areas that need future research attention or could benefit from harnessing existing pharmacogenetics knowledge toward rational and optimal therapeutics with warfarin in African patients. A literature search was conducted until January 2019. A total of 343 articles were retrieved from nine African countries: Botswana, Ethiopia, Egypt, Ghana, Kenya, South Africa, Sudan, Tanzania, and Mozambique. We found 19 studies on genetics of warfarin treatment specifically among Africans. Genes examined included CYP2C9, VKORC1, CYP4F2, APOE, CALU, GGCX, and EPHX1. CYP2C9*2 and *3 alleles were highly frequent among Egyptians, while rare in other African populations. CYP2C9*5, *8, *9, and *11, and VKORC1 Asp36Tyr genetic variants explained warfarin variability in Africans better, compared to CYP2C9*2 and *3. In Africa, there is limited pharmacogenetics data on warfarin. Therefore, future research and funding commitments should be prioritized to ensure safe and effective use of warfarin in Africa. Lessons learned in Africa from the science of pharmacogenetics would inform rational therapeutics in hematology, cardiology, and surgical specialties worldwide.
Introduction
Coumarins such as warfarin, phenprocoumon, and acenocoumarol are prescribed for the prevention and treatment of thromboembolic disorders (Militaru et al., 2015; Moualla and Garcia, 2011). Furthermore, direct oral anticoagulants (DOACs) with better safety profiles, which include rivaroxaban, apixaban, dabigatran, and edoxaban, have recently been introduced in the market (Bauersachs, 2016; Hellenbart et al., 2017). In Africa, the most common indications for anticoagulation include atrial fibrillation, venous thromboembolism, and their associated complications such as stroke (Awolesi et al., 2016; Kanyi et al., 2017; Nguyen et al., 2013).
Although the prevalence of these conditions is not well defined in Africans, they form part of cardiovascular diseases that contribute to increased global disease burden and cause of death in developing economies (Danwang et al., 2017; Nguyen et al., 2013; Stambler and Ngunga, 2015). Thus, warfarin remains the most widely prescribed and an anticoagulant of choice in Africa due to extensive knowledge of its use by physicians and low cost (Stambler and Ngunga, 2015), while other anticoagulants such as phenprocoumon, acenocoumarol, and DOACs are expensive and their use is limited to patients on private health insurance and those in resource-rich countries (Harrington et al., 2013; Shah et al., 2016).
Warfarin use is beset by interindividual variability and a narrow therapeutic index, which makes it difficult to titrate the optimal starting doses to reach the international normalized ratio (INR) (Kimmel, 2008; Wadelius and Pirmohamed, 2007). Prolonged titration to identify the appropriate dose to reach INR is associated with adverse drug effects and the need for hospitalization (Hafner et al., 2002; Kimmel, 2008). Therefore, warfarin requires rapid and continuous monitoring of the INR, before reaching stable INR of 2 to 3 (Jonas and McLeod, 2009).
The differences in patients' responses to warfarin have been attributed to both environmental and genetic factors (Jorgensen et al., 2009). Environmental factors include the following: race, age, gender, body weight, comorbidities, comedications, and lifestyle variables such as diet, alcohol consumption, and smoking (Dean, 2016; Jonas and McLeod, 2009; Kamali et al., 2004). For instance, warfarin dose requirements are reported to decrease with increasing age (James et al., 1992; Kamali et al., 2004).
With respect to diet, an inconsistent intake of vitamin K-rich foods such as green vegetables and vegetable oils affects warfarin dose requirements (Booth, 2010). Thus, individuals with decreased vitamin K intake are said to be sensitive to warfarin, whereas those with higher vitamin K intake may be resistant to warfarin (Johnson, 2005). Warfarin activity is also affected by medications such as amiodarone and rifampin through inhibition and induction of metabolism, respectively, consequently increasing or decreasing the INR levels (Bungard et al., 2011; Greenblatt and Moltke, 2005; Juurlink, 2007). Genetic factors include variants in genes involved in warfarin metabolism and genes coding for warfarin-targeted proteins (Rost et al., 2004; Sconce et al., 2005).
Warfarin is administered as a racemic mixture of S- and R-enantiomers. The S-form is the most potent (Johnson et al., 2017; Kaminsky and Zhang, 1997). Warfarin exerts its anticoagulant effects by inhibiting the action of vitamin K epoxide reductase complex 1 (VKORC1), an enzyme involved in the interconversion of vitamin K-2,3-epoxide (vitamin K epoxide) to reduced vitamin K hydroquinone (KH2) (Hirsh et al., 1998). The S-warfarin form is principally metabolized by CYP2C9, while R-warfarin is mainly metabolized by CYP1A2 and CYP3A4, with other enzymes such as CP2C18, CYP2C19, and CYP2C8 playing minor roles (Kaminsky and Zhang, 1997; Rettie et al., 1992).
Single-nucleotide polymorphisms (SNPs) in CYP2C9, VKORC1, and CYP4F2 have largely been shown to affect warfarin dose variability (Aithal et al., 1999; Caldwell et al., 2008; Chen et al., 2017; Muszkat et al., 2007; Rost et al., 2004; Sconce et al., 2005). Furthermore, variants in CYP2C9 and VKORC1 have been consistently shown to account for up to 40% of the total variability in warfarin dose requirements among Caucasians and Asians (Johnson et al., 2017; Ozer et al., 2010; Sconce et al., 2005; Veenstra et al., 2005). Hence, data on effects of genetic variants have been incorporated together with environmental factors in developing pharmacogenetics-based warfarin dosing algorithms.
These algorithms enable better prediction of warfarin starting dose that allows quick achievement of the required INR. The algorithms mostly include the following SNPs: VKORC1 g.-1639G>A, CYP2C9 c.430C>T (*2), and CYP2C9 c.1075A>C (*3), due to their inactivating effects on carrier enzymes (Alzubiedi and Saleh, 2016; Gage et al., 2008; The International Warfarin Pharmacogenetics Consortium, 2009).
Although warfarin pharmacogenetics algorithms have been applied in Caucasians and Asians, they have not been considered in native Africans (Finkelman et al., 2011; Kimmel et al., 2013; Pirmohamed et al., 2013). Furthermore, data used to develop these algorithms are extracted mostly from Europeans and Asians (Finkelman et al., 2011; Perera et al., 2014). Therefore, it is of importance to determine variants that influence warfarin variability in African populations with an effort to reduce the mismanagement of warfarin use and its associated side effects. This expert review evaluates the human genetics studies in African patients with respect to warfarin dose–response variations. The ultimate goal is identifying areas that still need attention or could benefit in making informed recommendations on the management of warfarin in Africans.
Literature Search
A literature search was conducted from the earliest data available until January 2019 using the following databases: PubMed, Google Scholar, Academic Search Premier, and MEDLINE. The keywords used individually or in combination in the search included the following: “Warfarin AND Africa,” “Pharmacogenetics AND warfarin AND Africa,” “Warfarin algorithm AND Africa,” and “Warfarin AND Blacks.” A total of 343 articles were retrieved from the search and studies were filtered down using the study selection criteria illustrated in Figure 1. After identification and screening, 34 studies were eligible for review.
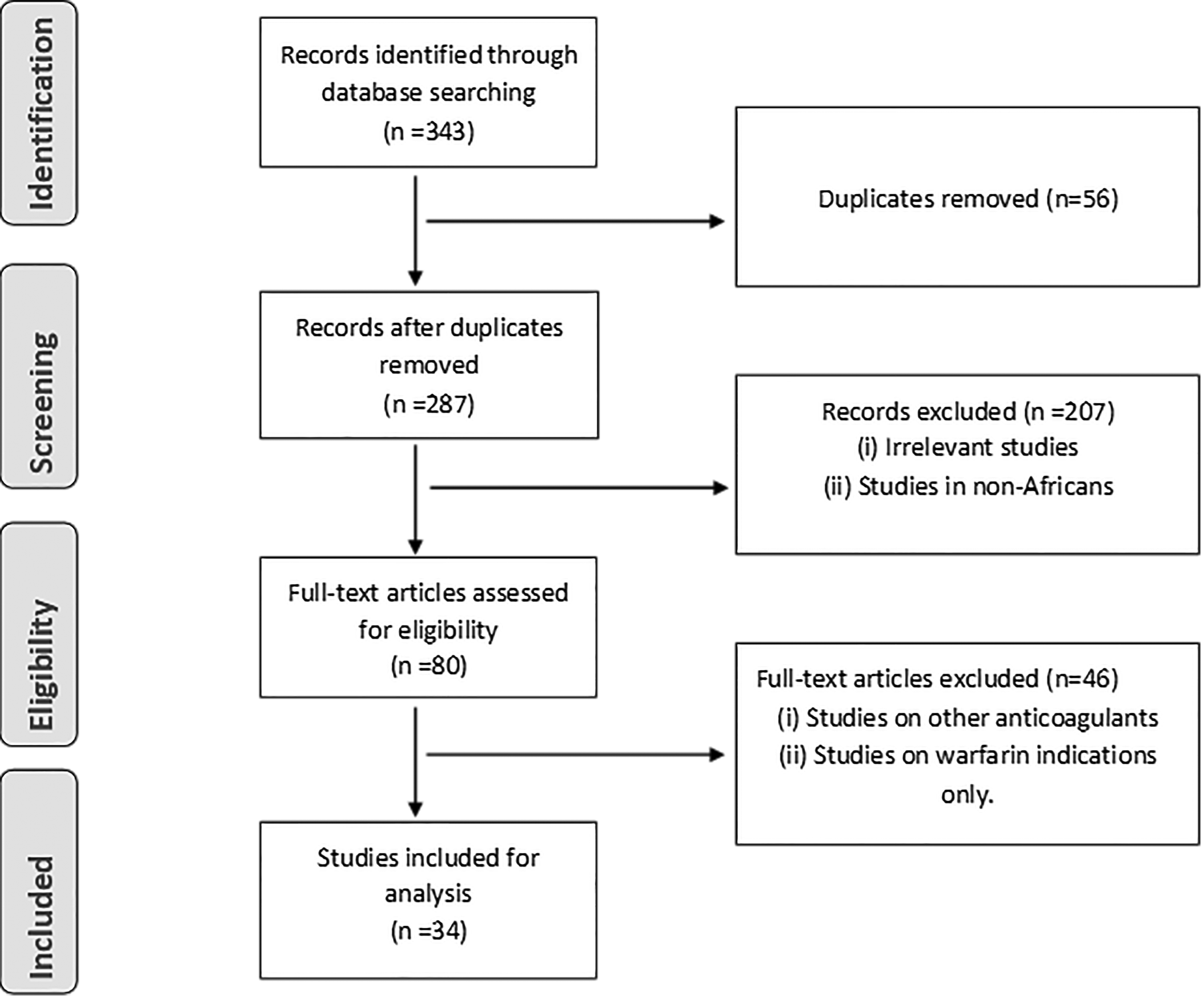
Flowchart of the literature search employed in this review.
Selection criteria
The inclusion criteria were as follows: (1) studies focusing on warfarin therapy in African populations, (2) articles focused on either genetics of warfarin alone or with both genetics and nongenetics factors, (3) research articles, and (4) articles published in English.
Articles excluded comprised review articles, studies on warfarin in non-African populations, studies focused on the use of other anticoagulants such as other coumarin derivatives and DOACs, and studies reporting on warfarin indications only.
Data extraction
For each study, the following data were extracted: the investigators, title of the study, the year of publication, study country, patients' data (e.g., demographic data), clinical data (indications and comorbidities), and studied genes.
Results
A total of 34 studies conducted in native Africans undergoing warfarin treatment were identified for the purpose of the expert review. The studies were conducted in individuals from the following nine African countries: Botswana, Ethiopia, Egypt, Ghana, Kenya, South Africa, Sudan, Tanzania, and Mozambique. The most common indication for warfarin in African patients was mechanical heart valve replacement (19–89%), followed by vascular heart disease (17–75%), atrial fibrillation (6–65%), and venous thromboembolism (4–65%) (Table 1). More than 30% of the studies were conducted in Egypt (n = 11). Genes explored included CYP2C9, VKORC1, CYP4F2, APOE, CALU, GGCX, and EPHX1 (Table 1).
Clinical, Demographic and Genetic Outlook in Africans Undergoing Warfarin Therapy
Median (interquartile range).
AF, atrial fibrillation; ARVs, antiretrovirals; BA, black Africans; CHD, coronary heart diseases; CHF, congestive heart failure; COPD, chronic obstructive pulmonary disease; CVD, cardiovascular diseases; DVT, deep vein thrombosis; GSH, Groote Schuur Hospital; IHD, ischemic heart disease; MA, mixed ancestry; MHVR, mechanical heart valve replacement; MPC, Mitchell's Plain Community Health Centre; MVD, mixed valve disease; PE, pulmonary embolism; SD, standard deviation; TB, tuberculosis; VHD, valvular heart diseases; VTE, venous thromboembolism.
The warfarin indications were accompanied by comorbidities such as HIV/AIDS and tuberculosis coinfections (Kanyi et al., 2017; Tarus et al., 2013). The medications used for the treatment of comorbidities pose risks of drug–drug interactions with warfarin (Schapkaitz et al., 2018). For example, some studies have shown that antiretroviral therapy potentiates an extended anticoagulation and reduces INR when coadministered with warfarin (Kanyi et al., 2017; Schapkaitz and Sithole, 2017).
The risks of warfarin use have led to recommendations for warfarin anticoagulation monitoring clinics as services to improve anticoagulation control (Louw et al., 2018; Mbokota et al., 2018). Anticoagulation control refers to when patients would spend 60% of the time within the required warfarin therapeutic range (Ahmed et al., 2017; Ebrahim et al., 2018; Manji et al., 2011). However, it is worth noting that pharmacogenetics should be accounted for when managing warfarin treatment as it has been reported to explain over 40% of warfarin variability (Sconce et al., 2005; Veenstra et al., 2005; Wadelius et al., 2009).
CYP2C9 is the main enzyme involved in the metabolism of warfarin, reducing S-warfarin to 6- and 7-hydroxywarfarin metabolites (Rettie et al., 1992). Hence, CYP2C9 is the most studied pharmacokinetic gene with regard to warfarin response (Johnson et al., 2017). Although there are several CYP2C9 variants that have been reported (www.cypalleles.ki.se; www.pharmvar.org), CYP2C9*2 and CYP2C9*3 are the most studied with regard to warfarin response, especially in Caucasians and Asians (Aithal et al., 1999; Chen et al., 2017; Kou et al., 2017; Sconce et al., 2005; Scott et al., 2008; Veenstra et al., 2005; Wadelius et al., 2009).
CYP2C9*2 and CYP2C9*3 alleles reduce enzyme activity, thereby decreasing warfarin metabolism by ∼30% and 80%, respectively (Aithal et al., 1999; Johnson et al., 2017). However, studies conducted across different African population groups (including African Americans) have reported absence or rarity of both CYP2C9*2 and CYP2C9*3 (Afilal et al., 2017; Allabi et al., 2005; Bazan et al., 2012, 2014; Dandara et al., 2011; Kudzi et al., 2009; Ndadza et al., 2019; Oliveira et al., 2009; Shrif et al., 2011) (Table 2). Hence, a lack of influence of these variants on warfarin variability is notable in populations with an African ancestry (Kudzi et al., 2016; Mitchell et al., 2011; Ndadza et al., 2019; Schapkaitz and Sithole, 2017).
Minor Allelic Distribution of Variants in African Populations in Comparison with Other World Populations
The African American, European, and Asian allele frequencies were extracted from (https://www.ncbi.nlm.nih.gov/projects/SNP).
Mozambique, bEthiopia, cBenin, dMorocco, eGambia, and fTunisia.
−, no data.
Understandably, African populations such as Egyptians and Tunisians have reported the presence of CYP2C9*2 and CYP2C9*3 (Table 2), and this can be explained by their pronounced admixture with Asian and Caucasian populations (Abdelhedi et al., 2015; El-Din et al., 2012; Ghozlan et al., 2015; Shahin et al., 2011). Thus, CYP2C9*2 and CYP2C9*3 have been significantly associated with low warfarin dose requirements in adults of Egyptian descent, explaining over 5.2–15.6% of warfarin variability (Table 3) (Bazan et al., 2014; El-Din et al., 2012; Ghozlan et al., 2015; Shahin et al., 2011).
Warfarin Dose–Response Variation Explained by Genetic and Environmental Factors
Black Africans and bMixed ancestry.
BMI, body mass index; BSA, body surface area; TIA, transient ischemic attack.
Due to the rare nature of CYP2C9*2 and CYP2C9*3 in Africans, focus has now shifted to alternative CYP2C9 variants. Other CYP2C9 variants that are being explored in Africans include the following: CYP2C9*5, *6, *8, and*11 (Dickmann et al., 2001; Limdi et al., 2015). CYP2C9*5, *6, *8, and *11 highly contribute to warfarin response in Africans as their occurrence has been associated with a reduced warfarin response in both African Americans and native Africans (Cavallari et al., 2010; Mitchell et al., 2011; Perera et al., 2011; Shahin et al., 2011; Shrif et al., 2011). The combined analysis of these variants with either CYP2C9*2 or *3 has been shown to improve warfarin dose predictions by explaining ∼5% and 6% to 7% in Sudanese and African Americans, respectively (Cavallari et al., 2010; Perera et al., 2011; Shrif et al., 2011) (Table 3).
The inclusion of these variants improve warfarin dosing model from 30% to 36% in African Americans (Cavallari et al., 2010). Furthermore, the Clinical Pharmacogenetics Implementation Consortium guideline recommends the warfarin pharmacogenetics-based dosing to be utilized in African Americans only when information on the CYP2C9*5, *6, *8, and*11 genotypes is available (Johnson et al., 2017).
VKORC1 is the principal pharmacodynamic gene of interest in warfarin response. Genetic variation in VKORC1 has been described to be the major contributor to warfarin variability, explaining ∼15–34% of the variability (Ozer et al., 2010; Sconce et al., 2005; Veenstra et al., 2005). VKORC1 g.-1639G>A is the most studied VKORC1 SNP and occurs in the VKORC1 promoter region. VKORC1 g.-1639A is associated with reduced level of expression of the VKORC1, subsequently reducing warfarin dose requirements by over 30% (D'Andrea et al., 2005; Johnson et al., 2017; Wadelius et al., 2009).
The occurrence of VKORC1 g.-1639A variant varies among different African population groups, occurring at a high allele frequency of 46–51%, 37%, and 31% in Egyptians, Sudanese, and South Africans of mixed ancestry, respectively, compared to Black South Africans (12%), Ghanaians (6%), and Mozambicans (3.5%) (Bazan et al., 2014; Ghozlan et al., 2015; Kudzi et al., 2016; Ndadza et al., 2019; Shahin et al., 2011; Shrif et al., 2011; Vargens et al., 2011) (Table 2).
The influence of VKORC1 g.-1639G>A on warfarin response is reportedly low among Africans compared to non-African populations, as a 10–13% and 4–7% warfarin variability have been reported in Egyptians and African Americans, respectively, compared to the 15–34% reported in Caucasians and Asians (Cavallari et al., 2010; Ghozlan et al., 2015; Limdi et al., 2010; Shahin et al., 2011). VKORC1 g.-1639G>A occurs at high linkage disequilibrium (LD) with C1173T (rs9934438), G1542C (rs8050894), C2255T (rs2359612), and T4931C (rs7196161) (collectively defined as VKORC1*2) in Caucasians and Asians (Lee et al., 2006; Rieder et al., 2005); however, in Africans, the LD is only strong between VKORC1 g.-1639G>A and C1173T (rs9934438) (r2 = 0.95) (Limdi et al., 2008, 2010; Schelleman et al., 2007; Shrif et al., 2011).
Hence, in other African studies, VKORC1 C1173T is the most targeted SNP, as it is also associated with low warfarin dose requirements (Bazan et al., 2014; Ekladious et al., 2013; El-Din et al., 2012; Ghozlan et al., 2015; Shahin et al., 2011; Shrif et al., 2011).
There are other VKORC1 polymorphisms that have been associated with warfarin resistance and these include the following: VKORC1 g.9041G>A located in the 3′ untranslated region and VKORC1 Asp36Tyr and Val66Met both located in the VKORC1 coding regions. VKORC1 g.9041A occurs at a high frequency in Africans (43–52%) and Europeans (35–37%) compared to Asians (10–17%) (Rieder et al., 2005; Takahashi et al., 2006). However, the influence of VKORC1 g.9041G>A on warfarin response is inconsistent among Africans, as it has been significantly associated with a high warfarin dose in Sudanese and black South Africans (Mitchell et al., 2011; Shrif et al., 2011), whereas a nonsignificant difference in maintenance dose between the VKORC1 g.9041G>A genotypes has been reported in African Americans (Cavallari et al., 2010; Schelleman et al., 2010).
VKORC1 Asp36Tyr has been reported in Ethiopians, Kenyans, Sudanese, and Egyptians at a frequency of 15%, 6%, 6%, and 2.5%, respectively (Shahin et al., 2013; Sominsky et al., 2014). However, it is reported to be absent or rare in black South Africans and West African descendants such as Ghanaians and African Americans (Mitchell et al., 2011; Shahin et al., 2013). The presence of VKORC1 Asp36Tyr has been reported to increase warfarin variability by 5.5% in Egyptians (Shahin et al., 2013). VKORC1 Val66Met has been described in Brazilians of African descent, who are warfarin resistant; furthermore, a Tanzanian male who was taking 21 mg/day of warfarin was reported to be heterozygous for VKORC1 Val66Met (Lewis et al., 2016; Orsi et al., 2010).
EPHX1, CALU, GGCX, APOE, and CYP4F2 are other pharmacodynamic genes that have been described. These genes have not been extensively studied in native Africans, as they have been mainly explored in Egyptians (Hamadeh et al., 2016; Issac et al., 2014; Shahin et al., 2011). EPHX1 rs1057140, CALU rs2290228, GGCX rs699664, GGCX rs12714145 (C>T), and CYP4F2 p.V433M have been reported to have a lack of influence on warfarin dose in Egyptians and African Americans (Bress et al., 2012; Limdi et al., 2015; Schelleman et al., 2010; Shahin et al., 2011; Voora et al., 2010).
APOE-ɛ2 has been associated with a low warfarin dose requirement, explaining 2.5% of warfarin variability in Africans (Kimmel et al., 2008; Shahin et al., 2011), while CALU rs339097 has been associated with an increased warfarin dose requirement and accounted for 5.7% of warfarin variability in African Americans and Egyptians (Ramirez et al., 2012; Shahin et al., 2011). A microsatellite (CAA) tandem repeats in intron 6 of GGCX (rs10654848) has been described to play a role in warfarin response. The GGCX (CAA) rs10654848 was associated with an increased warfarin dose in African Americans, possessing at least 16 CAA repeats and further explained 2% of the warfarin variability (Cavallari et al., 2012), while in Egyptians, only one individual carried the 16 CAA repeats with a higher warfarin dose of 70 mg/week (Hamadeh et al., 2016).
Furthermore, the increased warfarin dose tended to be higher with the increase in number of CAA repeats (<15 vs. ≥15 CAA repeats); however, the difference was not statistically significant (p = 0.16) (Hamadeh et al., 2016).
Although data on warfarin pharmacogenetics are still lacking in Africans, the few studies conducted have shown that inclusion of additional variants other than CYP2C9*2 and *3 and VKORC1 g.-1639A improve prediction of warfarin variability in Africans (Table 3). Therefore, further identification of novel variants through application of molecular techniques that detect multiple variants simultaneously such as Mass Arrays and whole exome analysis from next generation sequencing (NGS) is warranted. Technological advancement in the pharmacogenetics of warfarin in African Americans has allowed identification of novel and rare variants important for warfarin response. These include the rs12777823G>A polymorphism that was identified in the CYP2C cluster upstream of CYP2C18.
The rs12777823G>A was identified by Perera et al. (2013) in a genome-wide association study (GWAS) and was found to reduce warfarin dose requirement by 10–25%, exclusively in African Americans. The effect of the rs12777823G>A polymorphism was further confirmed by our group, wherein this SNP reduced warfarin dose requirement in Black South Africans, thereby explaining 12% of warfarin variability (Ndadza et al., 2019). Another novel variant (rs7856096) was identified in a study that applied whole exome sequencing. The rs7856096 is located upstream of exon 2 of the folylpolyglutamate synthase gene and it improves the variance explained by the International Warfarin Pharmacogenetics Consortium (IWPC) dose equation of 30% with an addition of 3.3% in African Americans (Daneshjou et al., 2014).
Outlook
The field of pharmacogenomics entails the utilization of an individual's genetic composition to personalize therapeutic regimens and improve treatment outcomes. The application of pharmacogenomics is important in drugs such as warfarin that are difficult to use due to their narrow therapeutic range and interindividual variability. The Food and Drug Administration (FDA) has recommended pharmacogenetics testing in warfarin dosing (Gage and Lesko, 2008). Since the FDA recommendation, there has been an increased development in the pharmacogenetics of warfarin. Studies have identified environmental and genetics variants that explain over 50–60% of warfarin variability (Johnson et al., 2017; Zhang et al., 2011).
Several groups, including Gage et al. (2008) and the IWPC, have developed pharmacogenetics-based algorithms for use in warfarin dosing. Native Africans are not represented in the development of warfarin pharmacogenetics algorithms because most data available are obtained from Caucasians and Asians. Extrapolating data from other population groups to Africans is not appropriate as Africans are genetically diverse and allelic frequencies vary between different population groups (Ramsay, 2012; Tishkoff et al., 2009). Furthermore, there is a heightened admixture in the African continent (Luis et al., 2004; Rotimi et al., 2017).
The large genetic diversity in native Africans is highlighted in this expert review as different allelic frequencies varied among African populations. For example, CYP2C9*2 and CYP2C*3 are frequent among Egyptians, while rare in black South Africans and Ghanaians (Table 2). Therefore, the contribution of each genetic variation on warfarin response differs from one population to the next, with specific variants playing significant roles in particular populations.
The variability in the influence of specific variants on warfarin response among Africans is also driven by the nonhomogeneous nature of other population groups such as Egyptians and Sudanese. Egyptian and Sudanese gene pools are contributed by sub-Saharan Africans, Europeans, and Asians. Hence, genetic polymorphisms affecting warfarin response in Egyptians are skewed toward both Europeans and Africans. This is highlighted by the increased contribution of CYP2C9 c.430C>T (*2), CYP2C9 c.1075A>C (*3), and VKORC1 g.-1639G>A in warfarin variability in Egyptians. Yet, these polymorphisms play an insignificant role in sub-Saharan African populations. Thus, data on warfarin pharmacogenetics in one African population cannot be generalized or be used to infer on other African populations that have not been studied previously.
Africa comprised low- and middle-income economies, suggesting that there are few or limited resources and infrastructure to conduct expensive genomic research. Hence, warfarin pharmacogenetics studies are sparse and limited to <15% of the African countries. Furthermore, studies undertaken in Africans are mostly of candidate genes of variants described in either Europeans or Asians. Candidate gene studies limit detection of novel or rare SNPs that are only functional in Africans, such as the rs12777823G>A identified through a GWAS and was described to explain 5% of warfarin variability exclusively in African Americans.
Candidate gene studies undertaken in Africans mostly focus on CYP2C9, VKORC1, and CYP4F2, and to a lesser extent, APOE, CALU, and GGCX. However, there are over 30 genes that are involved in warfarin disposition, including CYP1A2, CYP3A4, CP2C18, CYP2C19, and CYP2C8 (Wadelius et al., 2007). Variants in some of the abovementioned genes have been described in Africans independent of warfarin response. For instance, CYP2C8*2 has been widely explored in Africans and is occurs at an allele frequency ranging from 9% to 24% (Adehin et al., 2017; Alessandrini et al., 2013; Motshoge et al., 2016; Röwer et al., 2005). Furthermore, at least one copy of the mutant allele exhibits decreased CYP2C8 enzyme activity in the metabolism of anticancer (paclitaxel) and antimalarial (amodiaquine) drugs (Adehin et al., 2017).
Consideration of variants in genes that have not been previously studied could increase the warfarin variability explained in Africans. The importance of inclusion of variants less studied or with no contribution in Europeans and Asians was highlighted in the review (Cavallari and Perera, 2012), as inclusion of variants such as CYP2C9*5, *8, *9, and *11, VKORC1 Asp36Tyr, and VKORC1 L120L increased the warfarin variability explained by commonly studied variants such as CYP2C9*2, CYP2C9*3, and VKORC1 g.-1639A (Table 3).
In addition to consideration of African-specific genetic variants, it is also important to consider inclusion of demographic, environmental, and social variables when developing pharmacogenetics dosing algorithms. This is highlighted on this expert review, as the inclusion of age, body weight, and concurrent medications improved warfarin variability explained by genetic factors (Table 3). However, there are no data on the effect of nutrition and usage of herbal medications on warfarin response in Africans. Consideration of nutrition is of paramount importance as vitamin K-rich diet has an effect on warfarin activity.
Furthermore, in some African populations, particularly in the rural areas, there is high consumption of traditional and indigenous vegetables such as the African green leafy vegetables (African spinach), which are the main source of the vitamin K and could ultimately affect warfarin response. Although studies have considered the effect of other conventional medications on warfarin response, it is worth noting that in most African countries, in addition to prescribed medications that are administered for diseases, patients also coadminister herbal medications or supplements with their prescribed drugs. The course of herbs and prescribed medication also potentiates herb–drug interaction and adverse effects.
Similar incidences of herb–drug interactions with consequential adverse effects have been reported in other populations. An example is the induction St. John's wort (a herbal) of CYP3A4, CYP1A2, and CYP2C9 activity, thereby resulting in increased warfarin clearance and reduced effects (Jiang et al., 2004). The challenge in African populations is that despite the widely accepted view that patients are coadministering herbal products with their medications, no conscious effort has been made to study the effects of these herbs on warfarin indications and bleeding events.
Having considered the limitations that are hindering development of warfarin pharmacogenetics in Africans, several recommendations can be made to improve on the accumulation of locally useful data that can assist in warfarin management. Collaboration among researchers from across Africa is essential, as this will enhance sample sizes and African countries studied. Furthermore, extension of collaboration with researchers from developed countries can leverage financial support for carrying out more extensive GWAS or NGS-anchored studies. This is exemplified by the International Warfarin Pharmacogenetics Consortium (2009), which developed a warfarin pharmacogenetics-based algorithm through collaboration of 21 research groups from 9 countries and 4 continents, thereby attaining genetic data from a total of 5700 patients.
Enhanced sample size and financial support will further enable studies such as GWAS to be undertaken. This would also allow application of technologically advanced molecular techniques such as MassArray platforms and whole exome sequencing by NGS. Technologically advanced molecular approaches enable the detection of novel and rare variants that have not been reported with regard to warfarin response and are specific for Africans. The increase in pharmacogenetics data on warfarin use in Africans will potentially allow for the development of pharmacogenetics algorithm best suited for Africans and possible future clinical trials.
Concluding Remarks
Warfarin is an essential drug in the management of thromboembolic disorders and will be more important clinically if optimal dosing algorithms are also available. Implementation of pharmacogenetics testing has the potential of allowing safe and quality anticoagulation. In Africa, there are limited pharmacogenetics data on warfarin; therefore, research in this regard should be prioritized to ensure safe and quality anticoagulation. However, the genetic diversity and the heightened admixture in Africans should be taken into consideration to generate data that will be beneficial to all. Collaboration and utilization of techniques that create and harness Big Data simultaneously are desirable and would accelerate discoveries in African pharmacogenetics studies.
Finally, new practices are emerging that impact on how we do science so as to cultivate innovations that are meaningful locally, and thus sustainable and equitable. In particular, there are attempts to understand the broader sociotechnical context as well as human values that cocreate scientific outcomes and innovation products. One example is the emerging field of responsible innovation (RI) that can help identify the human values that shape and steer collaboration among scientific groups, for collaboration is not a simple or a value-neutral practice. Greater competency of scientists in understanding the human values embedded in scientific decisions and choices would contribute to RI and help remedy the “two cultures divide” among laboratory sciences and critical social sciences (Bechtold et al., 2017; de Boer et al., 2018).
Such broader skills, if included in scientific training, would also help decipher and make transparent the embedded opaque values that drive scientific collaborations. Thus, the practices and principles of RI are often considered “sociotechnical integration research” and enable collaborations that are enriched by interdisciplinary or multidisciplinary perspectives. Ultimately, this would allow for robust and enduring responses to the broader social and equity dimensions of science in the 21st century as we generate, analyze, and contextualize Big Data to meaningfully improve the health of African patients and populations.
Footnotes
Acknowledgments
The Pharmacogenomics and Drug Metabolism Group is funded by the National Research Foundation (NRF) of South Africa and Medical Research Council of South Africa. The project on warfarin pharmacogenomics is particularly funded by the NRF.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.