
Abstract
Integrin α4β7, a CD4 independent receptor of human immunodeficiency virus-1 (HIV-1) gp120, defines a subset of CD4+T cells preferentially targeted by HIV. It is also considered as a promising therapeutic target for HIV-1 infection. Despite its role in HIV acquisition and disease progression, HIV-1-mediated integrin α4β7 signaling has not been elucidated so far. In view of this, we determined phosphoproteomic signatures of HIV-1 gp120 signaling as well as signaling mediated by the integrin α4β7 ligand, mucosal vascular addressin cell adhesion molecule-1 (MAdCAM-1), in primary CD4+ T cells. This is the first comprehensive report on MAdCAM-1 signaling, which is believed to enhance HIV-1 replication. Importantly, we identified proteins associated with both classical and nonclassical integrin functions. We observed that HIV-1 gp120 signaling is associated with proteins that have previously not been associated with HIV-1 pathogenesis and thus, need to be explored further. There was a significant overlap in proteins identified by both MAdCAM-1 and HIV-1 gp120 signaling, which most likely represents cellular processes triggered upon interaction of HIV-1 gp120 with integrin α4β7. Pathway analysis revealed enrichment of processes that could facilitate viral replication as well as viral entry through endocytosis. Although these results warrant independent replication and further validation, they suggest the presence of additional potential therapeutic targets. These results also suggest that combinatorial approaches for targeting both HIV-1 gp120 and MAdCAM-1 signaling may be necessary for efficient control of HIV-1 infection as well as novel innovation strategies in HIV therapeutics.
Introduction
Integrins are ubiquitously expressed cell surface receptors, which mediate interactions with other cells and the extracellular matrix (Anderson et al., 2014). Besides mechanical and structural functions, they also transmit signals bidirectionally and regulate numerous cellular processes, such as adhesion, migration, tissue extravasation, survival, and differentiation (Anderson et al., 2014). The molecular basis of integrin signaling is explained by an adhesome, which comprises all the proteins participating in the formation of focal adhesion complex (Zaidel-Bar et al., 2007). Integrins also serve as attachment and entry receptors for a number of viruses and the integrin-mediated signaling triggered by the bound virus can also promote viral replication (Stewart and Nemerow, 2007).
Integrin α4β7 is a leukocyte-specific integrin, which acts as a gut homing receptor (Wagner et al., 1996). Mucosal vascular addressin cell adhesion molecule-1 (MAdCAM-1), constitutively expressed on Peyer's patches, mesenteric lymph nodes, and lamina propria, is the natural ligand for integrin α4β7 (Bargatze et al., 1995; Berlin et al., 1995). It selectively interacts with integrin α4β7 and thereby helps to direct and increase leukocyte traffic to the gut-associated lymphoid tissue (Gorfu et al.., 2009). Integrin α4β7 also functions as an additional cellular receptor for human immunodeficiency virus-1 (HIV-1) (Arthos et al., 2008) and α4β7high CD4+ T cells are preferred targets for highly productive HIV-1 infection (Cicala et al., 2009).
Higher frequency of integrin α4β7-expressing CD4+ T cells is associated with increased risk of HIV acquisition and faster disease progression (Sivro et al., 2018). The importance of integrin α4β7 in the pathogenesis of HIV-1 infection is also highlighted by studies in nonhuman primates, which demonstrated the prophylactic and therapeutic potential of antibody-mediated blocking of integrin α4β7 (Byrareddy et al., 2014, 2016).
HIV-1 gp120 binds to and signals through integrin α4β7, by mimicking a tripeptide motif present on MAdCAM-1, resulting in activation of another integrin, LFA-1 (Arthos et al., 2008). The HIV-1 gp120-integrin α4β7-mediated activation of LFA-1 is believed to increase efficiency of HIV-1 infection by promoting formation of virological synapse between infected and uninfected cells (Arthos et al., 2008; Cicala et al., 2009).
Recently, MAdCAM-1 costimulation through integrin α4β7 has also been reported to modulate surface expression of HIV-1 coreceptor, CCR5, as well as to promote HIV-1 replication (Nawaz et al., 2018). Although HIV-1 gp120-mediated CD4 and chemokine signaling has been studied earlier (Wiredja et al., 2018; Wojcechowskyj et al., 2013), details of HIV-1 gp120-mediated integrin α4β7 signaling, especially the proteins participating in the signaling pathway, remain largely unknown despite evidence of the role of integrin α4β7 in HIV-1 acquisition and disease progression. Besides the information of a few components (Murata et al.., 2002), proteins involved in MAdCAM-1-mediated integrin α4β7 signaling also remain unknown to a large extent.
To address these gaps in literature, we studied phosphoproteomic signatures of HIV-1 gp120 and MAdCAM-1 signaling in α4β7-expressing primary CD4+ T cells. The results of our study provide evidence for a crosstalk between these signaling pathways and identify a set of proteins representing the contribution of integrin α4β7 toward HIV-1 gp120 signaling. Exploring the role of some of these proteins, which have not been previously reported to be linked with the infection, is likely to offer a novel insight into HIV pathogenesis as well as novel pathways to innovation in HIV therapeutics.
Materials and Methods
Purification and culture of CD4+ T cells
The study was approved by the Indian Council of Medical Research–National Institute for Research in Reproductive Health Ethics Committee for clinical studies (Project No. 224/2012). Peripheral blood was obtained from four healthy individuals (two males and two females) between the ages of 26 and 40 years (median age of 33 years) after obtaining written informed consent.
Peripheral blood mononuclear cells were isolated using HiSepTM LSM 1077 density gradient separation media (HiMedia Labs, Mumbai, India) and CD4+ T cells were purified (>95%) by negative immunomagnetic separation using the EasySepTM CD4+ T cell Enrichment Kit (StemCell Technologies, Vancouver, Canada). Purified CD4+ T cells were activated with the anti-CD3 mAb, OKT-3 (eBiosciences, San Diego, CA, USA), IL-2 (20 IU/mL) (PeproTech, NJ, USA), retinoic acid (15 nM; Sigma, MO, USA), and cultured in RPMI-1640 (HiMedia Labs) containing 10% fetal bovine serum (FBS; HiMedia Labs) along with antibiotics in a humidified incubator at 37°C with 5% CO2 for 7 days.
CD4+ T cell stimulation with integrin α4β7 ligands
CD4+ T cells in culture, were deprived of IL-2 and retinoic acid, 18 h before stimulation with the integrin α4β7 ligands and maintained in RPMI with 1% FBS to minimize basal signaling. Cells were then washed with phosphate-buffered saline (PBS) and incubated in HEPES-buffered saline containing 0.5% (w/v) bovine serum albumin, 1 mM MnCl2, 100 μM CaCl2 at 4°C, and treated with either 50 nM recombinant HIV-1 gp120 (Sino Biologicals, Inc., Wayne, PA, USA), or 50 nM recombinant MAdCAM-1 (R&D Systems, Inc., Minneapolis, MN, USA) or left untreated for 20 min. Following this, the cells were maintained at 37°C for 3 min, washed with ice cold PBS, and the cell pellets were frozen and stored at −80°C until further analysis. The above treatments were performed on two independent batches of cells, which were then separately processed and treated as biological replicates.
Phosphoproteomic analysis
The frozen pellets of the HIV-1 gp120 or MAdCAM-1 treated and untreated CD4+ T cells were thawed on ice and processed as previously described (Pinto et al., 2015). Briefly, the cells were lysed, sonicated, and centrifuged to separate proteins in supernatant from cellular debris. Following protein quantitation using the bichinconinic acid assay (Pierce, Waltham, MA, USA), 700 μg protein was subjected to reduction with dithiothreitol, alkylation with iodoacetamide, precipitated using acetone, digested with trypsin, and the peptides so obtained were labeled with six-plex isobaric tandem mass tags (Thermo Scientific, Rockford, IL, USA) according to the manufacturer's instructions.
These peptides were pooled, desalted with C18 reversed-phase column (Waters Corporation, Milford, MA, USA), and lyophilized, and further fractionated by basic pH reversed-phase liquid chromatography, dried using speedvac followed by TiO2-based phosphopeptide enrichment (Deb et al., 2019; Pinto et al., 2015), and subjected to liquid chromatography/mass spectrometry/mass spectrometry analysis using an LTQ-Orbitrap fusion mass spectrometer (Thermo Fisher Scientific, Bremen, Germany) interfaced with Easy-nLC II nanoflow liquid chromatography system (Thermo Scientific, Odense, Denmark).
The mass spectrometry data were searched using MASCOT (Perkins et al., 1999) (Version 2.2.0) and SEQUEST (Eng et al., 1994) search algorithms against a human RefSeq database (O'Leary et al., 2016) using Proteome Discoverer 1.4 (Version 1.4.0.288) (Thermo Fisher Scientific) and the search parameters used were as described previously (Nirujogi et al., 2015).The false discovery rate was set to 1% at the peptide level. Phosphopeptides with >75% site localization probability were identified and considered for further analysis. The peptide abundance ratio of gp120/control and MAdCAM/control were considered for determining the hyperphosphorylated and hypophosphorylated phosphosites. An arbitrary cutoff of 1.5-fold was considered to identify the confident list of dysregulated phosphosites. Thus, the abundance ratio values of >1.5 or <0.5 in any one of the two replicates were considered to be differentially phosphorylated.
Gene ontology enrichment
Gene ontology (GO) analysis for differentially phosphorylated proteins upon HIV-1 gp120 and MAdCAM-1 stimulation was performed using FunRich Functional Enrichment Analysis Tool (Pathan et al., 2015). GO terms with p < 0.001 were considered to be significantly enriched.
Kinome map
The kinome map was built using the KinMap tool (http://kinhub.org/kinmap/index.html). Illustration reproduced courtesy of Cell Signaling Technology, Inc.
Motif analysis
Motif-x algorithm was used to analyze the predicted consensus phosphorylation motifs in the HIV-1 gp120 and MAdCAM-1 dataset. (http://motif-x.med.harvard.edu) (Chou and Schwartz, 2011; Schwartz and Gygi, 2005). The criteria used for analysis included a sequence window of 6 amino acids on either side of each of the identified phosphorylation site, the minimum motif occurrence was set to 10 with significance threshold of p < 0.02. The background dataset for this motif enrichment analysis was derived from the whole phosphoproteome identified in our analysis. One of the enriched motifs, 14-3-3 binding motif, was further analyzed using 14-3-3-Pred (Madeira et al., 2015), a webserver for prediction of 14-3-3-binding sites in proteins.
Functional interaction network around differentially phosphorylated proteins
STRING database was used to construct functional interaction networks around differentially phosphorylated proteins (https://string-db.org) (Szklarczyk et al., 2015). GO analysis was used to highlight proteins involved in relevant biological processes. An interaction score of highest confidence (0.9) and disconnected nodes were hidden.
Results
To decipher the integrin α4β7 signaling network, stimulation of primary human CD4+ T cells was carried out using both MAdCAM-1, the natural ligand for integrin α4β7, as well as HIV-1 gp120, the pathogen-derived ligand. CD4+ T cells from four healthy participants were cultured in the presence of retinoic acid to obtain integrin α4β7high CD4+ T cells that were pooled. To investigate early signaling events, changes in phosphorylation at 3 min following stimulation of α4β7high CD4+ T cells with 50 nM of either MAdCAM-1 or HIV-1 gp120 were studied by quantitative phosphoproteomics.
The combined workflow for phosphoproteomic analysis is outlined in Figure 1A and B. A total of 1769 phosphopeptides (Supplementary Table S1) were identified in both the replicates with 2038 unique phosphosites, which corresponded to 953 proteins. We found that 71% of phosphopeptides were phosphorylated at a single site, whereas 22% phosphopeptides exhibited phosphorylation at two sites. The distribution of phosphorylation sites was found to be highest on serine residues (1828, i.e., 89%) followed by threonine (191, i.e., 9.3%) and tyrosine residues (19 i.e., 0.9%) (Fig. 1C).
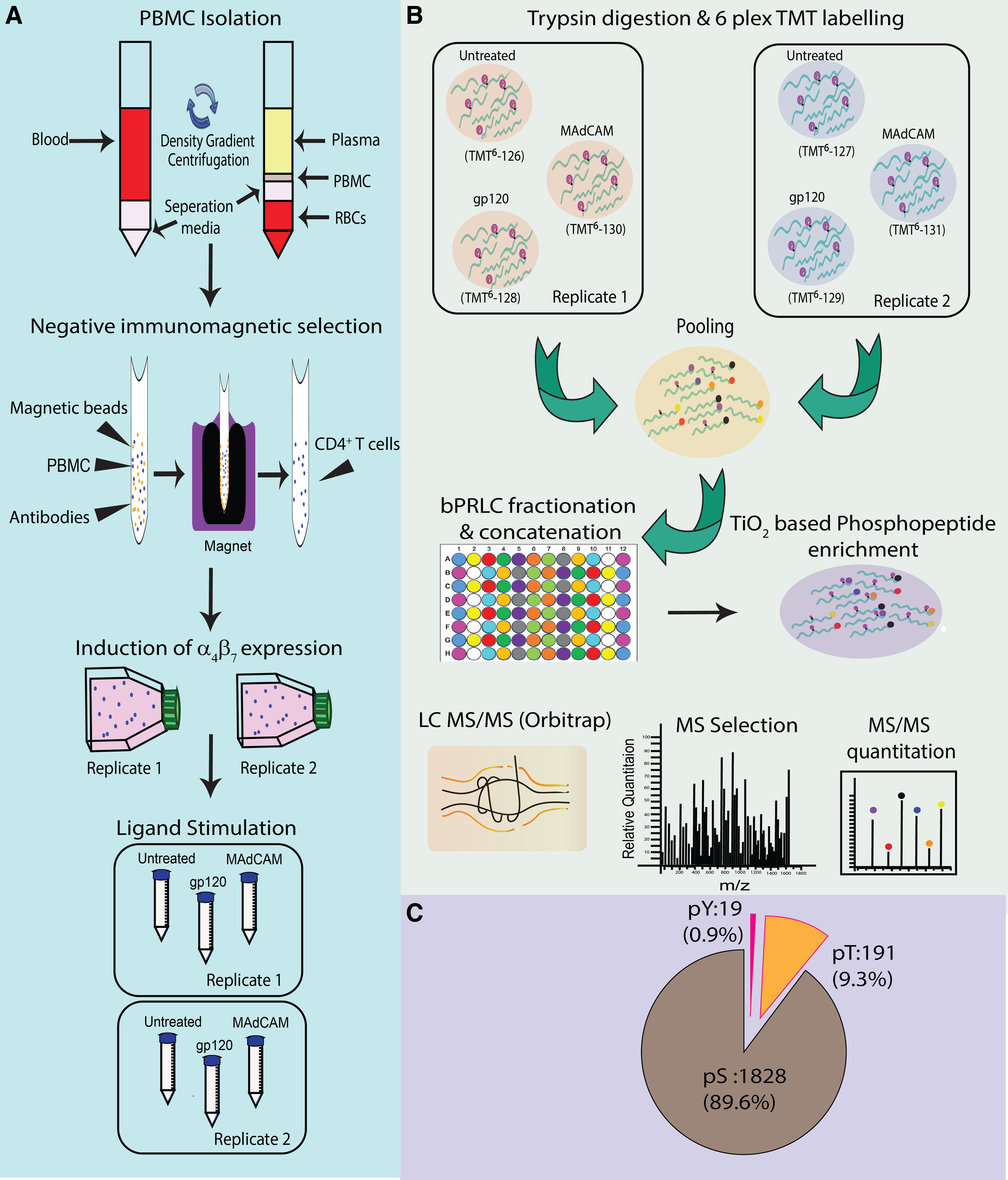
Schematic representation of workflow for quantitative phosphoproteomic analysis of HIV-1 gp120 and MAdCAM-1-triggered signaling in α4β7high CD4+ T cells.
Phosphoproteomic profile of MAdCAM-1 signaling
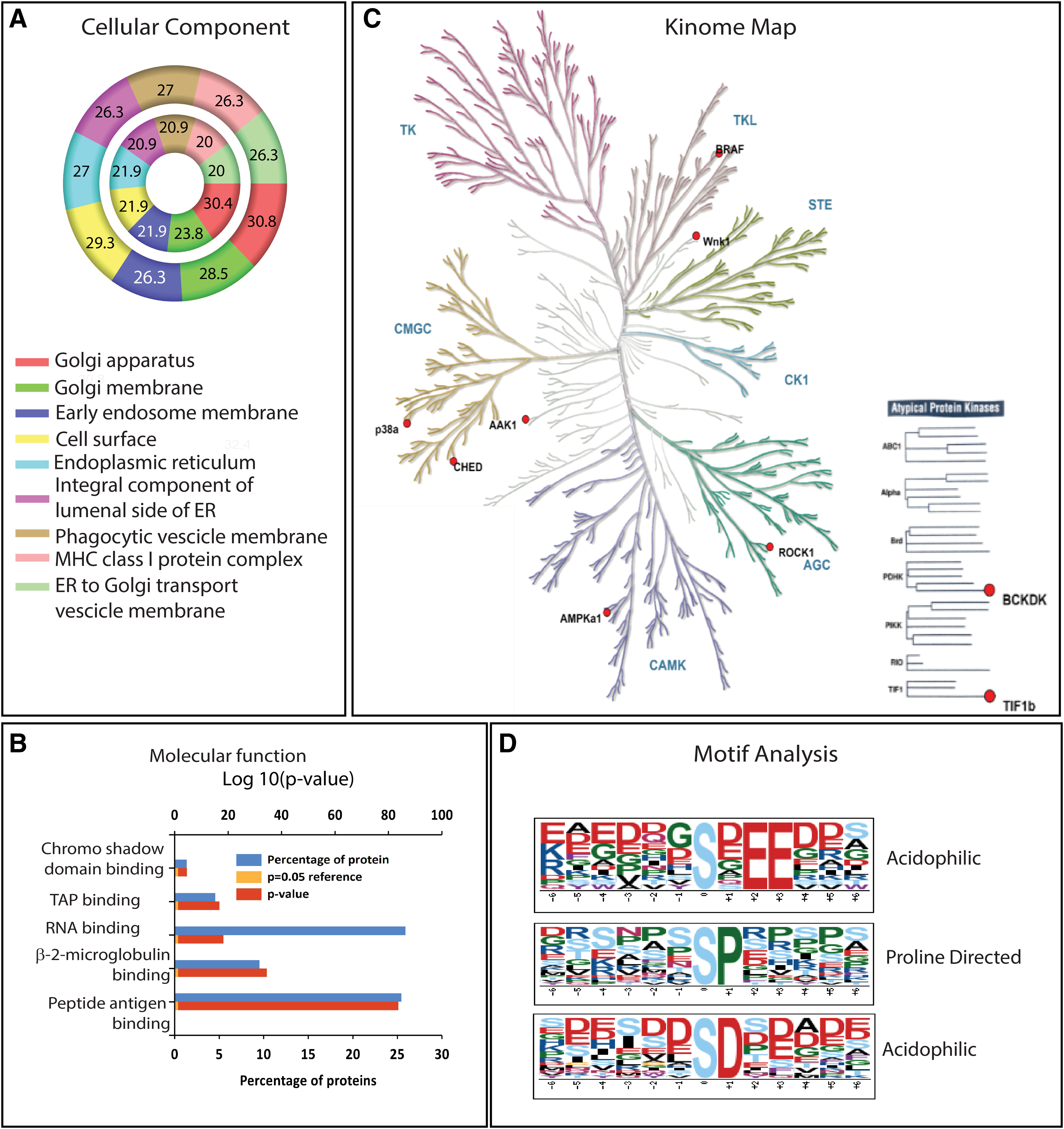
In response to MAdCAM-1 stimulation of CD4+ T cells, we identified 176 proteins, including integrin β7 to be differentially phophorylated (Supplementary Table S2). GO analysis was carried out using FunRich 3.1.3 with Uniprot Human database (UniProt release 2018_10) (Fig. 2A, B). Both hypophosphorylated and hyperphosphorylated proteins showed enrichment of GO terms for cellular components, such as cell surface, Golgi apparatus, endoplasmic reticulum, endosomes, and vesicles involved in intracellular transport.
Phosphoproteomic profile of MAdCAM-1 signaling.
Differentially phosphorylated proteins showed involvement in molecular functions, such as transporter for antigen presentation, peptide antigen and beta2-microglobulin binding, processes linked to antigen presentation through major histocompatibility complex (MHC) class I, which are exploited during viral infections (Hewitt, 2003). Differentially phosphorylated proteins also showed significant enrichment of biological processes, such as T cell activation (CD5, CCDC88B, RASAL3, SASH3, and FANCD2) and T cell proliferation (CD6, actin-binding protein CORO1A). The kinases, which were differentially phosphorylated following MAdCAM-1 stimulation, are represented in the kinome map (Fig. 2C and Supplementary Table S3). Since kinases recognize specific motifs around which phosphorylation takes place, we used the motif-x algorithm to identify phosphorylation motifs of MAdCAM-1-mediated signaling.
Motif-X analysis revealed the presence of nine distinct phosphorylation motifs enlisted in Supplementary Table S4. Logos for identified proline-directed and acidophilic motifs are depicted in Figure 2D. Acidophilic motifs, such as pSD and pSxEE, are known to be phosphorylated by CK2 kinases (Songyang et al., 1996) recruited by B-Raf (Ritt et al., 2007), a scaffolding protein kinase found to be hyperphosphorylated at S729 residue. Phosphorylation of B-Raf at S729 by AMPK facilitates binding of 14-3-3 protein homodimers (Martin and Marais, 2013; Shen et al., 2013). Interestingly, differential phosphorylation of YWHAZ, a 14-3-3 protein, was also observed following MAdCAM-1 signaling.
Additionally, WNK1 Kinase, which has been recently shown to play a critical role in T cell adhesion and migration, was also found to be differentially phosphorylated (Köchl et al., 2016). Other differentially phosphorylated kinases includes cholinesterase-related cell division controller, which is a` cyclin-dependent kinase required for RNA splicing. ROCK-1, which is a key regulator of actin cytoskeleton and AAK,1 which plays an important role in integrin trafficking during clathrin-mediated endocytosis contributes toward integrin recycling during cell migration (Nishimura and Kaibuchi, 2007; Sorensen and Conner, 2008).
Overall, we observed that MAdCAM-1 signaling through integrin α4β7 involves kinases and proteins participating in integrin-associated functions, such as adhesion, migration, and endocytosis most likely carried out through actin cytoskeletal rearrangement. Besides these, there are certain proteins involved in housekeeping functions such as RNA transport and mRNA surveillance (Supplementary Table S5).
MAdCAM-1-integrin α4β7 phospho-adhesome
Integrin–ligand interactions give rise to intracellular signaling involving adhesion complex proteins, referred to as the adhesome (Zaidel-Bar et al., 2007). The phosphorylated proteins associated with the adhesion complex are termed as phospho-adhesome (Robertson et al., 2015). Since MAdCAM-1 interacts exclusively with integrin α4β7, the resulting signaling process is expected to generate an adhesion complex. Comparison of proteins differentially phosphorylated in response to MAdCAM-1 stimulation with the phospho-adhesome revealed a significant overlap (hypergeometric test; p = 1.44e-19) comprising 32 common proteins (Fig. 3 and Supplementary Table S2). These included both, an intrinsic protein, ITGB7 (adhesion receptor), as well as associated proteins such as ARHGEF2 (GEF), ARHGAP5 (GAP), and ROCK1 (serine/threonine kinase). The differential phosphorylation of adhesome components observed following MAdCAM-1 stimulation confirms the occurrence of early integrin-mediated signaling.

MAdCAM-1 functional interaction network around differentially phosphorylated proteins. Interactions of proteins differentially phosphorylated upon MAdCAM-1 signaling are represented based on STRING database. GO analysis of biological process shows involvement of viral process (red), cell activation (blue), mRNA splicing (yellow), and leukocyte cell–cell adhesion (green) among others. Proteins previously reported to participate in phospho-adhesome formation are termed as MAdCAM-1 adhesome and are indicated in
Phosphoproteomic profile of HIV-1 gp120 signaling
HIV-1 gp120 stimulation led to differential phosphorylation of 181 proteins (Supplementary Table S6). GO analysis revealed differences between MAdCAM-1 and HIV-1 gp120 signaling. Both hypophosphorylated and hyperphosphorylated proteins were similarly enriched for GO terms related to cellular components in case of HIV-1 gp120 signaling (Fig. 4A). These comprised the nucleus and cytoplasm unlike MAdCAM-1 signaling, which led to differential phosphorylation of proteins belonging to organelles, such as endoplasmic reticulum, Golgi, intracellular vesicles, and cell surface.

Phosphoproteomic profile of HIV-1 gp120 signaling.
Molecular functions such as poly (A) RNA binding and chromatin binding, involved in enhancing mRNA stability and translation as well as gene regulation respectively, were found to be enriched (Fig. 4B). This probably indicates early response to viral infection. Kinome map depicts the differential phosphorylation of only typical kinases, such as CaMK2g, CK1g2, ROCK-1, WNK-1, AAK-1, and p38 (Fig. 4C and Supplementary Table S7).
Attempts to identify targets for these kinases using iGPS 1.0 revealed EIF4B to be a substrate for both CAMK2g and CK1g2. Sixteen motifs surrounding serine phosphorylation were identified using Motif X (Supplementary Table S8). These comprised acidophilic, proline-directed, and basophilic motifs, as well as 14-3-3-binding motifs. (Fig. 4D).
The phosphosites of the differentially phosphorylated proteins (Supplementary Table S9) corresponding to these 14-3-3-binding motifs were predicted by all 14-3-3 PRED classifiers to bind to 14-3-3 proteins with high confidence. Interestingly, we found YWHAZ, a 14-3-3 protein, to be hypophosphorylated at S230 in the C-terminal domain, which in turn regulates its binding to other proteins. In view of the potential of nonphosphorylated 14-3-3 proteins to modulate the activity of its binding partners (Dubois et al., 2002; Rommel et al., 1996), it is likely that YWHAZ may play a role in activation/deactivation of the above-predicted 14-3-3-binding proteins upon binding to them.
HIV-1 gp120 responsive phosphoproteins relevant to HIV-1 Infection
To understand the relevance of the HIV-1 gp120 responsive phosphoproteins identified in our dataset, we compared our results with the HIV-1 human protein interaction database (HHPID) (Fu et al., 2009; Pinney et al., 2009; Ptak et al., 2008). HHPID is a manually curated list of over 18,000 interactions involving more than 4000 human proteins reported to interact with HIV-1. We observed a significant overlap (100/181; hypergeometric test, p = 1.04e-22) with HHPID (Supplementary Table S6). Our dataset comprised proteins interacting with all nine primary HIV-1 proteins (Fig. 5).

HIV-1 interacting proteins. Proteins observed to be differentially phosphorylated upon HIV-1 gp120 stimulation in the present study that has been previously reported to interact with HIV-1 proteins in HHPID. HHPID, and HIV-1 Human Interaction Database.
Initial signaling through HIV-1 envelope protein, gp120, is expected to involve actin cytoskeleton rearrangements to facilitate virus entry. Hyperphosphorylation of the cytoskeletal proteins, SOS1, GSN, PXN, WAS, MSN, GIT1, ROCK1, and PPP1R12C, observed in this study, may bring about cytoskeletal rearrangements promoting viral entry. Differential phosphorylation of 10 out of 79 peptides (112 phosphosites) belonging to SRRM2, which modulates host cell alternative splicing machinery during HIV-1 entry and thereby facilitates virus replication and release (23), was also observed (Supplementary Table S10) in the present study.
An overlap of the differentially phosphorylated proteins in our dataset was also observed with previously reported phosphoproteomics-based CXCR4-mediated (31/181; hypergeometric test, p = 4.4e-32) HIV-1 signaling. These results indicate that the identified HIV-1 gp120 responsive proteins (Supplementary Table S6) are involved in HIV-1 infection.
HIV-1 gp120 responsive phosphoproteins associated with integrin signaling
Since we aimed to decipher integrin α4β7-mediated HIV-1 gp120 signaling, we examined the overlap of the HIV-1 gp120 responsive proteins with phospho-adhesome proteins (Robertson et al., 2015), as these constitute the major portion of integrin signaling. A significant overlap (33/181; hypergeometric test, p-value = 3.5e-20) was observed between the two datasets and included intrinsic adhesome proteins like ITGB7 (adhesion receptor), MSN (adaptor), and AGAP2 (GAP), as well as adhesome-associated proteins, such as SOS1 (GEF), ARHGAP5 (GAP), and ROCK1 (serine/threonine kinase). The interactions of these proteins and their targets are termed as the “HIV-1 gp120 adhesome” (Fig. 6 and Supplementary Table S6).
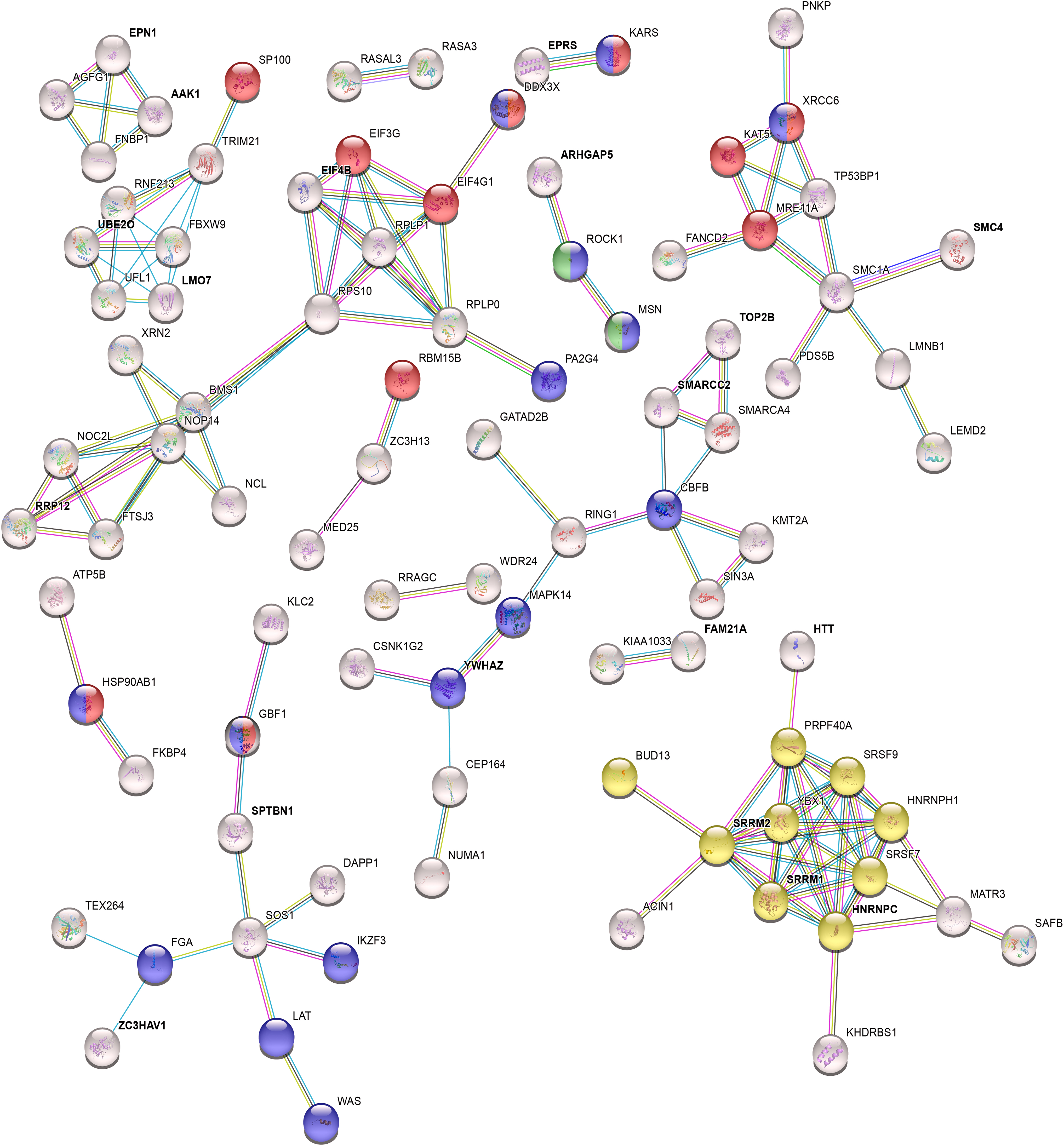
HIV-1 gp120 functional interaction network around differentially phosphorylated proteins. Interactions of proteins differentially phosphorylated upon HIV-1 gp120 signaling are represented based on STRING database. GO analysis of biological process shows involvement of viral process (red), cell activation (blue), mRNA splicing (yellow), and leukocyte cell–cell adhesion (green) among others. Proteins previously reported to participate in phospho-adhesome formation are termed as HIV-1 gp120 adhesome and are indicated in
The differential phosphorylation of integrin β7 at the cytoplasmic tail (T797) constitutes evidence for its involvement in the HIV-1 gp120 signaling. Furthermore, the differential phosphorylation of other adhesome components observed following HIV-1 gp120 stimulation also indicates the involvement of integrin-mediated signaling during early stages of HIV-1 infection.
HIV-1 gp120 signaling through integrin α4β7 results in enrichment of pathways associated with infection
Binding of HIV-1 gp120 to the cell surface results in signaling through multiple receptors. These include the primary receptor CD4 and coreceptors like chemokine receptors as well as attachment receptors, including integrin α4β7 (Arthos et al., 2008). Since both HIV-1 gp120 and MAdCAM-1 have similar integrin α4β7-binding motifs, intracellular signaling independently mediated by these ligands is anticipated to result in phosphorylation changes in proteins having an overlapping presence in both these common processes. We identified 95 such proteins (Supplementary Table S11), which may represent the contribution of integrin α4β7 toward HIV-1 gp120 signaling.
These included differentially phosphorylated proteins, such as PA2G4, CD5, WNK1 SAFB, PDS5B, IKZF3, PHIP, EIF1AD, and CCDC88B, which may regulate cell proliferation, differentiation, and apoptosis in turn supporting HIV-1 replication Out of these, 50 proteins have not been reported so far in the HHPID and need to be explored further for determining their role in HIV-1 infection (Fig. 7). Pathway analysis of all the proteins common between integrin α4β7 signaling through MAdCAM-1 and HIV-1 gp120 (Supplementary Table S5) led to identification of endocytosis pathway among other processes.

Contribution of integrin α4β7 toward HIV-1 gp120 signaling. Schematic representation of various stages of intracellular HIV-1 infection depicting functions of the predicted list of proteins common to HIV-1 gp120 signaling and MAdCAM-1 signaling identified in this study. This is indicative of the contribution of integrin α4β7-mediated HIV-1 gp120 signaling to HIV-1 infection.
Although internalization of the virus following membrane fusion is an accepted mode of HIV-1 infection, its uptake by endocytosis in CD4 and non-CD4 T cells is also well documented. Accordingly, our dataset shows differential phosphorylation of proteins associated with clathrin-mediated endocytotic process, such as AAK1, FNBP1, MFF, and SMAP2, whereas WDR44 and KIAA1033 are involved in endosome recycling. We also saw differential phosphorylation of proteins associated with actin cytoskeleton rearrangement, such as RCSD1, SNTB1, PHIP, ROCK-1, and ARHGAP5.
Following integration of viral DNA, a series of transcription, splicing, and translation cycles produce viral proteins along with parallel replication of RNA that is assembled together in virions. In line with this, we also found differential phosphorylation of proteins, such as BUD13, SRRM1, ACIN1, and SRRM2, participating in splicing; proteins involved in transcription, such as MED25, GATAD2B, NLC, CTR9, SAFB, TOP2B, SMARCC2, UBE2O, PAPOLG, MAPK14, and IKZF3; and translation-associated proteins, such as EIF1AD, RPLP1, DDX3X, and SLBP.
Discussion
We report, for the first time, mass spectrometry-based analysis of MAdCAM-1 signaling. Our results suggest that combinatorial approaches for targeting both HIV-1 gp120 and MAdCAM-1 signaling may be necessary for efficient control of HIV-1 infection as well as novel innovation strategies in HIV therapeutics. These findings can be placed to a broader integrative biology context as follows.
Viruses use integrins as attachment or entry receptors and promote pathogenesis by modulating multiple essential cellular processes, including intracellular signaling, growth, migration, survival, and apoptosis (Stewart and Nemerow, 2007). HIV-1 is a versatile virus using multiple receptors, including integrins to enhance its infectivity (Ballana et al., 2009, 2011; Cicala et al., 2011). It preferentially targets a subset of metabolically active integrin α4β7high CCR5high CD4+ T cells leading to productive infection (Cicala et al., 2009). Higher expression of integrin α4β7 is associated with HIV-1 acquisition and disease progression (Sivro et al., 2018). In view of this, antibody-mediated blocking of integrin α4β7 could be regarded as a potential therapeutic option for control of HIV infection (Byrareddy et al., 2014).
However, details of integrin α4β7-mediated HIV-1 gp120 signaling in CD4+ T cells such as the proteins involved in this pathway as well as the associated cellular processes have not been investigated. Similarly, MAdCAM-1 signaling has also not been studied in detail despite evidence for its role in promoting HIV-1 replication (Nawaz et al., 2018). The presence of multiple HIV-1 receptors on CD4+ T cells makes it challenging to delineate the contribution of an individual receptor. We, therefore, determined the overlap of phosphoproteomic signature of HIV-1 gp120 signaling with that corresponding to the ligand of integrin α4β7 to decipher its role in HIV-1 gp120 signaling.
Compared with a previous report on MAdCAM-1 signaling, which identified only paxillin and p105Cas-L (Murata et al., 2002), this study has identified several other proteins with wide ranging functions. Many of these proteins are part of the phospho-adhesome, which is considered as a hallmark of integrin signaling. Interestingly, we observed differential phosphorylation of proteins involved in MHC class I-associated antigen presentation, which is also used for displaying viral peptides, a function that has not been previously attributed to MAdCAM-1 signaling.
Additionally, processes linked to mRNA quality control, such as RNA transport, RNA splicing were also observed. Altered antigen presentation along with T cell proliferation and activation, may be linked to enhanced HIV-1 replication reported to occur following MAdCAM-1 costimulation (Nawaz et al., 2018).
Integrin signaling influences cellular processes, such as adhesion, migration, actin cytoskeleton rearrangement, and cell proliferation. The kinases involved in these processes were also observed to be differentially phosphorylated upon MAdCAM-1 stimulation. Moreover, we identified components of endocytosis to be differentially phosphorylated. These proteins may be largely involved in integrin trafficking for recycling to the cell surface. Thus, in addition to the integrin-associated functions, data indicate the involvement of MAdCAM-1-mediated integrin α4β7 signaling in nonclassical functions such as antigen presentation and mRNA quality control, which in the context of HIV-1 infection may contribute to presentation of HIV-1 antigens on the surface of CD4+T cells and thereby promote immune activation and pathogenesis.
HIV-1 interaction with T cells is extensively studied and quantitative phosphoproteomics-based HIV-1 signaling has also been previously investigated (Wiredja et al., 2018; Wojcechowskyj et al., 2013). However, the analysis of integrin α4β7high CD4+ T cells in this study led to the delineation of the contribution of integrin α4β7 in HIV-1 signaling, unlike the previous studies. These studies involve the use of the whole virus, while we have used CCR5 tropic recombinant HIV-1 gp120 for stimulating CD4+ T cells that were cultured ex vivo to maximize expression of integrin α4β7 before HIV-1 gp120 treatment. The use of primary CD4+ T cells subjected to culture conditions resulted in practical limitations, which led to the small sample size of two biological replicates.
Furthermore, despite retinoic acid treatment to increase expression of α4β7, uniform expression of the integrin could not be achieved in the primary CD4+ T cells. Owing to this, it is likely that the biological replicates showed variability in magnitude of the phosphorylation changes. Hence, to obtain meaningful interpretation of the data, we considered proteins to be differentially phosphorylated even if one the replicates exhibited fold changes above or below the cutoff, which was arbitrarily selected. However, in spite of these limitations as well as the changes in duration of HIV-1 gp120 stimulation, differentially phosphorylated proteins observed in our study showed significant overlap with those reported previously (Fu et al., 2009; Pinney et al., 2009; Ptak et al., 2008; Robertson et al., 2015; Wiredja et al., 2018; Wojcechowskyj et al., 2013).
Proteins associated with actin cytoskeleton rearrangement that can facilitate viral entry, were observed in our dataset. Many of the differentially phosphorylated proteins have been previously reported to interact with different HIV-1 proteins. This suggests that, the detected signaling events influence cellular pathogenesis beyond mere viral entry. Early HIV-1 signaling has been reported to influence splicing events occurring later during the infection cycle. In concurrence with this, differential phosphorylation of the splicing machinery-associated proteins, SRRM1 and SRRM2, were noted in this study, although at sites different than the ones reported previously.
Since integrin signaling is characterized by formation of adhesome, the HIV-1 gp120 phospho-adhesome represented in this study is indicative of the intersection of integrin and HIV-1 gp120 signaling. HIV-1 gp120-associated integrin signaling has not been reported previously, hence it was decided to characterize the contribution of integrin α4β7 to HIV-1 gp120 signaling. To achieve this, differentially phosphorylated proteins common to both HIV-1 gp120 and MAdCAM-1 signaling were further analyzed.
Pathway analysis suggests the possibility of integrin α4β7 involvement in virus internalization through clathrin-mediated endocytosis, which has been demonstrated to be an important mode of HIV-1 entry (Miyauchi et al., 2009). This is also supported by the dependence of HIV-1 entry (Daecke et al., 2005) on dynamin, a GTPase essential for pinching off endocytic vesicles during clathrin-mediated endocytosis. Integrin α4β7 has been shown to be incorporated in HIV-1 virions, thereby facilitating infection and gut homing (Guzzo et al., 2017).
It is likely that some of the virions following binding to integrin α4β7 and internalization by endocytosis may be recycled to the cell surface and released along with bound integrin α4β7, thereby supporting its incorporation by virions. As suggested earlier (Permanyer et al., 2010) endosomal entry of HIV-1 may be a mechanism for viral escape from the immune system and HIV-1 entry inhibitors. This finding provides an insight into the mechanisms involved in attenuation of HIV-1 infection upon antibody-mediated blocking of integrin α4β7 (Byrareddy et al., 2014; Cicala et al., 2009).
Irrespective of the mode of HIV-1 entry, either through membrane fusion or endocytosis, proteins differentially phosphorylated through integrin α4β7 can contribute to the various stages of HIV-1 cellular pathogenesis. The results of this study also support the molecular mimicry of MAdCAM-1 by HIV-1 gp120 (Arthos et al., 2008) not just at the level of binding, but also at the level of intracellular signaling. Besides this, many proteins identified in the present study have not yet been reported in HHPID. This may be on account of the lack of studies focused on integrin α4β7 signaling in CD4+ T cells during HIV-1 infection. These proteins may be involved in novel interactions that need to be confirmed by experimental approaches.
Additionally, the findings of our study related to the identification of proteins involved in MAdCAM-1 signaling and their association with HIV-1 gp120-mediated integrin α4β7 signaling as well as with viral entry need to be experimentally validated. The results of the present study thus indicate the occurrence of processes, such as endocytosis, actin cytoskeleton rearrangement, integration of viral DNA, transcription, splicing, and translation of HIV-1 proteins downstream to HIV-gp120-integrin α4β7 signaling, which could be additionally targeted to enable efficient control of HIV-1 infection. An example of this could be the exploration of endocytosis inhibitors which may supplement current therapeutic strategies as well as circumvent the hurdle posed by resistance to drugs, including those being investigated for inhibition of HIV-1 fusion.
These findings also suggest that the impact of potential therapeutic interventions aimed at blocking integrin α4β7, on some of these physiologically important processes, need to be determined. Finally, the results of the current study also support the growing evidence regarding a role for both HIV-1 gp120 and MAdCAM-1-mediated integrin α4β7 signaling in facilitating HIV-1 infection.
Conclusions
Integrin α4β7 is an important CD4 independent receptor of HIV-1, which is also regarded as a promising therapeutic target for prevention and control of infection. The present study cataloged proteins associated with both MAdCAM-1 and HIV-1 gp120 signaling in primary CD4+ T cells by examining phosphoproteomic signatures. Besides exhibiting a considerable overlap with proteins previously reported to be important for phosphoadhesome formation and HIV-1 infection, the study also identifies several HIV-1 gp120 responsive proteins that have so far not been associated with infection.
Pathway analysis of the proteins representing the intersection of MAdCAM-1 and HIV-1 gp120 signaling indicate their potential involvement in processes such as viral entry and replication. Validation of these findings is important for not only gaining additional insights into HIV-1 pathogenesis but also for expanding the therapeutic potential of integrin α4β7-targeted approaches in control of HIV-1 infection.
Footnotes
Author Contributions
N.K. performed the experiments, analyzed the data, and prepared the figures; P.K. in planning and supervision of the proteomics experiment; B.D. in data analysis; V.M.B. conceived the study, designed the experiments, and interpreted the data; V.M.B. and N.K. wrote the article. All authors contributed to revisions for important intellectual content, read, and approved the final article.
Acknowledgments
The authors thank the Director, Indian Council of Medical Research–National Institute for Research in Reproductive Health (ICMR–NIRRH) and Dr. K.V.R. Reddy for the support provided throughout the course of this study. They also thank Dr. Lalita Savardekar and the staff of the ICMR–NIRRH bone health clinic and women's health clinic, Naigaon, Mumbai, India, for help in screening and recruiting the participants for the study.
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
The work was supported by the Indian Council of Medical Research (ICMR) (5/7/852/12-RCH to V.M.B.); Department of Biotechnology, Government of India (BT/PR4876/MED/108/7/2012 to V.M.B.) collaborative grant for Human Immunodeficiency Virus-Acquired Immunodeficiency Disease Syndrome and Microbicides; Indian Council of Medical Research–National Institute for Research in Reproductive Health (intramural grant to V.M.B.), and the University Grants Commission and Junior Research Fellowship [21/12/2014 (ii) EU-V to N.K.].
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.