
Abstract
Regenerative medicine research and testing of new therapeutics for muscle-related human diseases call for a deeper understanding of how human myoblasts gain and maintain quiescence in vitro versus in vivo. The more closely we can experimentally simulate the in vivo environment, the more relevance in vitro research on myoblasts will have. In this context, isolation of satellite cells from muscle tissue causes activation while myoblasts remain activated in culture, thus not simulating quiescence as in their in vivo niche. Cells synchronized for cell cycle present a good starting point for experimental intervention. In the past, myoblast quiescence has been induced using suspension culture (SuCu) and, recently, by knockout serum replacement (KOSR)-supplemented culture media. We assessed the proportion of cells in G0 and molecular regulators after combining the two quiescence-inducing approaches. Quiescence was induced in primary human myoblasts (PHMs) in vitro using KOSR-treatment for 10 days or suspension in viscous media for 2 days (SuCu), or suspension combined with KOSR-treatment for 2 days (blended method, SuCu-KOSR). Quiescence and synchronization were achieved with all three protocols (G0/G1 cell cycle arrest >90% cells). Fold-change of cell cycle controller p21 mRNA for KOSR and SuCu was 3.23 ± 0.30 and 2.86 ± 0.15, respectively. Since this was already a significant change (p < 0.05), no further change was gained with the blended method. But SuCu-KOSR significantly decreased Ki67 (p = 0.0019). Myogenic regulatory factors, Myf5 and MyoD gene expression in PHMs were much more suppressed (p = 0.0004 and p = 0.0034, respectively) in SuCu-KOSR, compared to SuCu alone. In conclusion, a homogenous pool of quiescent primary myoblasts synchronized in the G0 cell cycle phase was achieved with cells from three different donors regardless of the experimental protocol. Myogenic dedifferentiation at the level of Myogenic Regulatory Factors was greater when exposed to the blend of suspension and serum-free culture. We suggest that this blended new protocol can be considered in future biomedical research if differentiation is detected too early during myoblast expansion. This shall also inform new ways to bridge the in vitro and in vivo divides in regenerative medicine research.
Introduction
Bridging the in vitro and in vivo divides continues to be an ongoing methodological challenge as well as prospect to improve medical research and innovation. This is also the case in the field of regenerative medicine and studies of human diseases impacting skeletal muscle tissue. In this context, adult muscle stem cells, also referred to as satellite cells, are located between the basal lamina and the sarcolemma in a reversible state known as quiescence (Mauro, 1961; Mohammad et al., 2019). The cellular quiescent state is marked by a cell cycle arrest induced by enhanced expression of cell cycle inhibitors such as p21 and p27 (Coqueret, 2003; Korgun et al., 2006; Li et al., 2015). Other regulators involved in maintaining the quiescent state include microenvironmental factors such as a low growth factor environment and attachment to the extracellular matrix (ECM) (Atsumi et al., 2011; Fukada, 2011). Evidence strongly suggests that stem cell quiescence is an actively maintained state (Cheung et al., 2012; Liu et al., 2013; Mourikis et al., 2012; Sampath et al., 2018) with several molecular mechanisms at play (Fuchs and Blau, 2020).
Quiescent cells are able to reenter the cell cycle and differentiate when the appropriate stimulus is provided (Cheung and Rando, 2013). Activation of muscle stem cells is achieved via the action of growth factors and physical contact changes with adjacent fibers (Kuang et al., 2008; Smith et al., 2008; Tatsumi et al., 2002; Wozniak et al., 2003) and cytokines, including several other members of the interleukin-6 family (Fuchs and Blau, 2020; Sampath et al., 2018; Steyn et al., 2019). It has been shown that many of these factors promote satellite cells' entry into the cell cycle (Shefer et al., 2006). Once activated, satellite cells can self-renew or differentiate into myogenic cells. Myogenic regulatory factors (MRFs), including Myf5 and MyoD, control satellite cell self-renewal and differentiation, respectively (Gayraud-Morel et al., 2007; Kablar et al., 1997; Megeney et al., 1996; Rudnicki et al., 1992; Rudnicki et al., 1993). The expression of each of these MRFs is associated with a specific stage of satellite activation and myogenic differentiation (Bentzinger et al., 2012; Zammit, 2017).
A small percentage of the satellite cells will return to the quiescent state, replenished via a self-renewal process that is tightly controlled by Myf5 (Fukada, 2011; Gayraud-Morel et al., 2007; Mohammad et al., 2019; Zammit, 2017) among other factors.
Although recent bioinformatic analyses of very large datasets and single-cell analyses have tried to identify factors or proteins associated with satellite cell quiescence (De Micheli et al., 2020; de Morrée et al., 2017; Sampath et al., 2018), validation requires testing identified factors in vitro. Quiescence-reactivation experiments utilizing suspension culture (SuCu) for quiescence induction have tried to elucidate molecular events regulating reversible quiescence (Aloysius et al., 2018; Cheung et al., 2012; Sellathurai et al., 2013). However, many inferences made about the G0-state of primary human myoblasts (PHMs) or satellite cells are based on in vitro studies that are inappropriate as proliferating cells in culture dishes or flasks display different biology to G0-state cells in vivo (Dumont et al., 2015; Forcina et al., 2019; Giordani et al., 2018; Motohashi et al., 2014; Musaro and Carosio, 2017; van Velthoven et al., 2017). It is therefore of paramount importance that such cells be synchronized after harvesting before being used in experiments.
Several protocols have been devised to isolate and expand human myoblasts from muscle biopsies and these methods show different efficacies (Agley et al., 2015; Spinazzola and Gussoni, 2017). Due to the small number of cells isolated, the PHMs require expansion before utilization in experiments or cell therapy. Expansion can be done in vitro using bioreactors and culture dishes in the laboratory, with addition of factors to stimulate proliferation. Given their heterogeneous nature, these cells are likely to enter cell cycle at different times (Relaix, 2006; Relaix and Marcelle, 2009). To overcome this phenomenon, it is critical to induce quiescence in muscle stem cells/PHMs before their use in different applications. Early studies used serum deprivation to synchronize cells before usage in experiments. For example, Ashihara and Baserga achieved cell cycle synchronization via the removal of serum from culture media (Ashihara and Baserga, 1979). Although the removal of serum from myoblast media improved synchronization by inhibiting cell proliferation, it appeared to also promote myogenic differentiation (Lin et al., 2003; Ma et al., 1998).
To negate the promotion of differentiation by serum deprivation, we recently developed and published a novel protocol that uses knockout serum replacement (KOSR) to induce quiescence in PHMs isolated from human muscle (Gudagudi et al., 2020). A further advantage is that KOSR has been used by others to ensure cellular survival (Ishii et al., 2015; Liu et al., 2016). As mentioned above, one of the specific characteristics of the muscle stem cell microenvironment is that the cells are surrounded by ECM and, therefore, spatially quite static when quiescent (Dinulovic et al., 2017; So and Cheung, 2018). This aspect may be part of the mechanism for successful induction of quiescence using SuCu (Mashinchian et al., 2018; Tierney et al., 2016). Different quiescence induction methods may result in major changes in molecular properties of cells (Liu et al., 2007; Nishikawa et al., 2008). Thus, it is important to evaluate and compare how different induction methods influence the final quiescent cells' stemness, proliferation, and regulators of differentiation. Immortalized cell lines such as C2C12 have been used before, but these cells are genetically abnormal, mainly cultured under in vitro conditions for a long time and this can result in cells deviating from normal function.
The aim of the present study was to determine if a combination of the SuCu method and the KOSR method would render myoblasts with more characteristics of quiescence. Quiescence was assessed via evaluation of expression of genes known to regulate myoblasts and cell cycle analysis. Variability is a major issue when using primary cells. We therefore isolated and expanded cells from three different volunteers, to study both induced quiescence and synchronization of myoblasts from different individuals. Our findings using isolated PHMs show that both the SuCu and KOSR methods are efficient in inducing quiescence in PHMs. In addition, the developed blended method, combining the SuCu and the KOSR methods, displayed better efficiency than either of the individual methods at reducing promyogenic regulatory factors and increasing cell cycle inhibition.
Materials and Methods
Human myoblast isolation
PHMs were isolated from three young healthy male subjects, aged between 18 and 23 years, using the microexplant technique (Smith et al., 2008) and expanded under normal cell culture. Written and informed consent was obtained from volunteers for the use of the biopsy samples and ethical approval was obtained from the Subcommittee C of the Human Research Ethics Committee of Stellenbosch University (Ethics Number: HREC N12/08/051, November 12, 2012) for the protocol used and the study.
In brief, a 5 mm trephine biopsy needle was used with assisted suction on the vastus lateralis of the subjects to allow the harvesting of ∼50–100 mg of skeletal muscle (Gudagudi et al., 2020). Muscle samples were immediately transferred to a cold buffer (1 × phosphate-buffered saline [PBS] solution containing 10% [v/v] penicillin/streptomycin [P/S, P43333; Sigma-Aldrich] and 1% [v/v] gentamicin [15750-060; GIBCO™]). Samples were washed with 1 × PBS, transferred into an entactin-collagen IV-laminin (ECL, 08-110; Merck, USA)-coated six-well tissue culture plates. Proliferation media consisting of Ham's F-10 Nutrient Mixture Medium (N6908; Sigma-Aldrich), supplemented with 20% (v/v) fetal bovine serum (FBS, 10499-044; Life Technologies), 1% (v/v) P/S (P43333; Sigma-Aldrich), and 10 ng/mL recombinant fibroblast growth factor-2 (rh-FGF2, G5071; Promega) were added. Biopsy samples were removed from the culture dish 14 days after transfer and then expanded. Purity of the cells was evaluated by measuring Pax7 and desmin expression.
Myoblast cell culture
PHMs were cultured as described by Steyn et al. (Steyn et al., 2019). PHM's proliferation media consisted of Ham's F-10 Nutrient Mixture Medium (N6908; Sigma-Aldrich), supplemented with 20% (v/v) FBS (10499-044; Life Technologies), 1 (v/v) P/S (P43333; Sigma-Aldrich), and 10 ng/mL fibroblast growth factor-2 (G5071; Promega). Differentiation was induced by culturing cells in Dulbecco's modified Eagle's medium (D5671; Sigma-Aldrich), supplemented with 1% horse serum (26-050-088; Life Technologies) and 1% P/S (15140-122; Life Technologies). PHMs were evaluated for their stemness and myogenic lineage via the measurement of CD34, CD56, Myf5, MyoD, p21, and Ki67 expression levels as previously described (Gudagudi et al., 2020).
Human primary myoblasts' characterization
PHMs were isolated and characterized by evaluation of Pax7 and desmin expression via flow cytometric analysis as well as identification of multinucleated tubes as described before (Steyn et al., 2019). Cells were trypsinized and resuspended in cold PBS at ∼1 × 106 cells/mL. Cells were fixed and permeabilized in a 1:1 Methanol/Acetone solution for 10 min on ice. Washing was done using PBS and resuspension was done in 1% (v/v) bovine serum albumin. In short, cells were incubated in the presence of primary antibodies for 30 min at room temperature and incubated with secondary antibodies before analysis using the BD FACSAria™ Cell Sorter (BD Biosciences, San Jose, CA, USA).
Myogenic differentiation was determined via treatment with differentiation media (Dulbecco's modified Eagle's medium, 1% horse serum). CellTracker with nuclear staining was performed to identify multinucleated tubes. Hoechst (cat # 62249; Thermo Fisher Scientific, MA, USA) 1:200 dilution and CellTracker (cat # C2925; Thermo Fisher Scientific) 1:1000 dilution in proliferation media were used to prepare the staining solution with final volume of 3 mL. Five minutes before the imaging, the staining solution was added to the well inside the hood and incubated at 37°C, and imaging was done using Nikon ECLIPSE E400 microscope. Hoechst nuclei stain is shown in blue and CellTracker stain is shown in green. Terminal differentiation was investigated via measuring myogenin levels.
Induction of cellular quiescence
PHMs were induced to a quiescent state (G0 state) via the use of three methods as outlined in detail below. In brief, cells were cultured in KOSR for 10 days, suspended for 2 days with FBS in the serum or for 2 days in suspension with KOSR. For each method, cells were harvested from proliferation media culture before the intervention started and control cells continued in proliferation media over the particular time period. (1) The KOSR-10 day method involved deprivation of FBS from culture media and the use of KnockOut™ Serum Replacement (KOSR, 10828-010; Life Technologies) to induce quiescence in PHMs as described before (Gudagudi et al., 2020). The final KOSR quiescence media contained Ham's F-10 Nutrient Mixture Medium (N6908; Sigma-Aldrich), supplemented with 20% (v/v) KOSR (10828-010; Life Technologies) and 1% (v/v) P/S (15140-122; Life Technologies). Culture period was 10 days in quiescence-induction media. Media was changed every day for the duration of the experiment to remove proproliferative secretions that might promote proliferation and differentiation. Evaluation of stemness, myogenic lineage, and proliferation was done by measuring p21, Myf5, MyoD, and Ki67 levels; (2)
Induction of quiescence using the SuCu was carried out as described in the literature (Arora et al., 2017; Rumman et al., 2018). In brief, 2% methylcellulose was used to prepare SuCu. Proliferating PHMs were trypsinized and 1 × 106 cells in 500 μL growth media were mixed with FBS (10%, v/v), 2% methylcellulose, penicillin, streptomycin, GlutaMAX, and HEPES (10 mM, pH 7.3), up to final volume of 10 mL. Methylcellulose final concentration was 1.3%. After induction of quiescence for 48 h, cells were obtained by diluting methylcellulose cell suspension with prewarmed PBS at 37 0C. Cell suspension was centrifuged at 1800 g for 30 min at room temperature followed by removal of supernatant three to four times until most of semisolid methyl cellulose was removed. The pellet containing cells was dispersed by gentle pipetting followed by washing of cells with PBS combined with gentle centrifugation. As described before, this treatment of cells will result in optimal viability (Arora et al., 2017). Cells were counted and viability measured via the use of trypan blue staining. The resulting cells were lysed for RNA isolation or used in cell cycle analysis as described elsewhere; (3) The blended method for quiescence induction was similar to the SuCu method as described (2) above, with the only difference being KOSR replacing FBS in the SuCu media. Incubation of PHMs in SuCu media was done for 48 h before cells were harvested as described above.
Cell cycle analysis
Cell cycle analysis was performed as done before (Gudagudi et al., 2020) using the BD Cycletest™ Plus DNA Reagent Kit (340242; BD Biosciences, San Jose, CA, USA). Cells were fixed with 40% methanol, stained with propidium iodide according to manufacturer's specifications, and filtered through a 50 μm nylon mesh. Flow cytometric analysis was done immediately using the BD FACSAria™ Cell Sorter (BD Biosciences) to determine DNA ploidy. On average, 1 × 104 cells were analyzed and proportions of cell cycle fractions, G1, S, and G2 were quantified with ModFit LT 3.0 (Verity Software).
RNA isolation and quantitative real-time reverse transcriptase polymerase chain reaction
Cells were lysed with the TriPure (cat # 11667157001; Sigma-Aldrich (Pty) Ltd, MO, USA) and RNA was isolated according to manufacturer's instructions. RNA quantity and quality were assessed using the Nanodrop spectrophotometer, and cDNA was prepared using 1 μg total RNA. Real-time quantitative polymerase chain reaction (PCR) was performed using the StepOnePlus™ Real-Time PCR system with SYBR Green dye. Samples were made in triplicates for analysis together with positive, negative, and blank controls. Primers included: GAPDH (sense), 5′-TGGTGCTGAGTATGTCGTGGAGT-3′, and (antisense), 3′-AGTCTTCTGAGTGGCAGTGATGG-5′; Myf5 (sense), 5′-AATTTGGGGACGAGTTTGTG-3′, and (antisense), 3′-CATGGTGGTGGACTTCCTCT-5′; MyoD (sense), 5′-TGCACGTCGAGCAATCCAAA-3′ and (antisense), 3′-CCGCTGTAGTCCATCATGCC; Ki67 (sense), 5′-TGACCCTGATGAGAAAGCTCAA-3′ and (antisense), 3′-CCCTGAGCAACACTGTCTTTT-5′; p21 (sense), 5′-TCTTGTACCCTTGTGCCTCG-3′ and (antisense), 3′-ATCTGTCATGCTGGTCTGCC-5′ and Pax7 (sense), 5′-CCCCCGCACGGGATT-3′ and (antisense), 3′-TATCTTGTGGCGGATGTGGTTA-5′. Fold change in gene expression between control and test samples was calculated using the delta delta Ct method (ΔΔct) method. GAPDH was used as a normalizer.
Statistical analysis
Data are presented as mean ± standard error of the mean of three independent experiments. Statistica (Version 13.2; Dell, Inc., 2016) was used for statistical analysis. Two-way repeated measures analysis of variance was used to compare the effects of quiescence versus proliferation. For comparisons of ki67, myf5, and p21 expression in quiescent cells, data were expressed relative to the corresponding proliferation control, and one-way analysis of variance was used to compare the effects of the different quiescence induction methods. These tests were followed by Fischer post hoc testing. p < 0.05 was deemed statistically significant.
Results
Characterization of isolated PHMs
Primary myoblasts isolated from skeletal muscle display higher sarcomere assembly than immortalized cells such as C2C12 when proliferating and differentiating in vitro (Manabe et al., 2016). Thus, isolated primary myoblasts do represent skeletal muscle properties better than immortalized C2C12 cells/myotubes (Manabe and Fujii, 2016). In this study, we utilized isolated myoblasts to evaluate the developed quiescence-inducing protocol (Experimental Scheme, Fig. 1). Characterization of isolated PHMs was achieved through the use of immunofluorescence staining for Pax7 and desmin as well as evaluation of known myoblast regulatory factor gene expression as described in our previous publications (Gudagudi et al., 2020; Steyn et al., 2019). In addition, myogenic differentiation of the PHMs was done via induction of differentiation for 5 days and immunostaining performed for desmin and myogenin (Steyn et al., 2019). Stocks of previously developed PHM clones as described by Steyn and colleagues (Steyn et al., 2019) and Gudagudi and coworkers were used in this study (Gudagudi et al., 2020) (Fig. 1).

Quiescence induction in PHMs using KOSR or SuCu
Quiescence was induced in isolated PHMs as shown in Figure 1 above. To validate induction of cellular quiescence using the 10 day KOSR protocol, control PHMs actively proliferating over the same 10 days were harvested for comparison via flow cytometry. Cell cycle analysis indicated that a large proportion (>90%) of cells were arrested in G0/G1 state in PHMs cultured in KOSR media for 10 days compared to that seen in the proliferating cells, while a significant decrease was noted in the S phase (Fig. 2A–E). All three PHM (KH1, KH3, and S6.3) clones used in the study showed that when PHMs were induced into quiescence, there was an increase in cells in G0/G1 phase in quiescent cells than in proliferating cells (Fig. 2A–E). Concurrently, the number of cells in S-phase decreased when cells were cultivated in KOSR media compared to normal proliferating media (Fig. 2A–E). Quiescence induction using the SuCu method yielded similar results to the use of KOSR media (Fig. 3A–E). Culturing PHMs in suspension resulted in significant increase in cells in the G0/G1 phase (>90% on average) compared to PHMs remaining unsuspended in proliferating media (Fig. 3A–E). This pattern was similar for all three PHM clones used in the study (Fig. 3A–E). Concurrently, the number of cells in the S-phase decreased significantly as cells enter quiescence (Fig. 3A–E).

Effect of proliferation media and KOSR media on PHMs (KH1, KH3, and S6.3) cell cycle progression at 10 days.
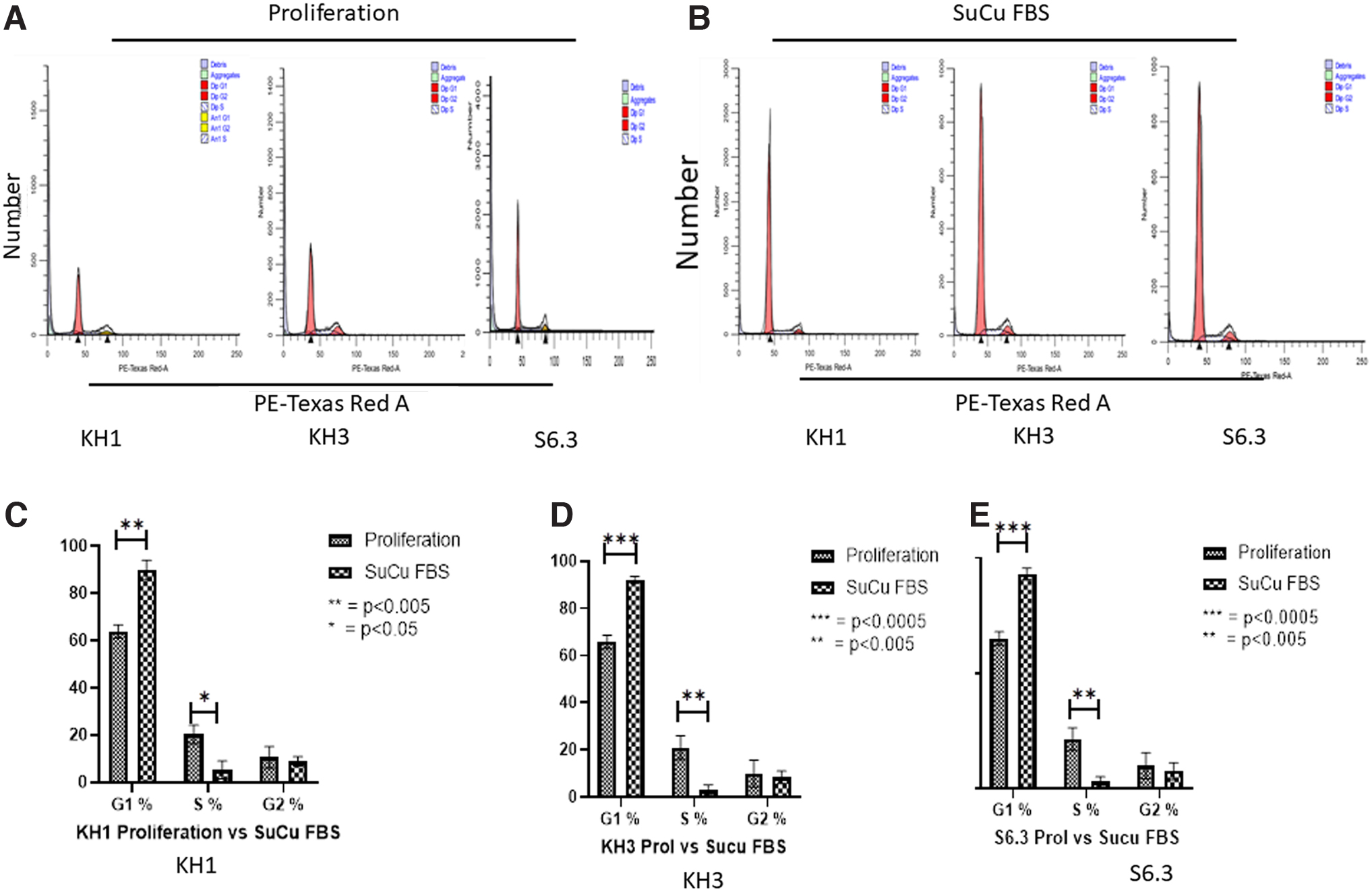
Effect of proliferation media and SuCu on PHMs cell cycle progression at 10 days.
PHMs molecular characteristics following culture in KOSR or SuCu
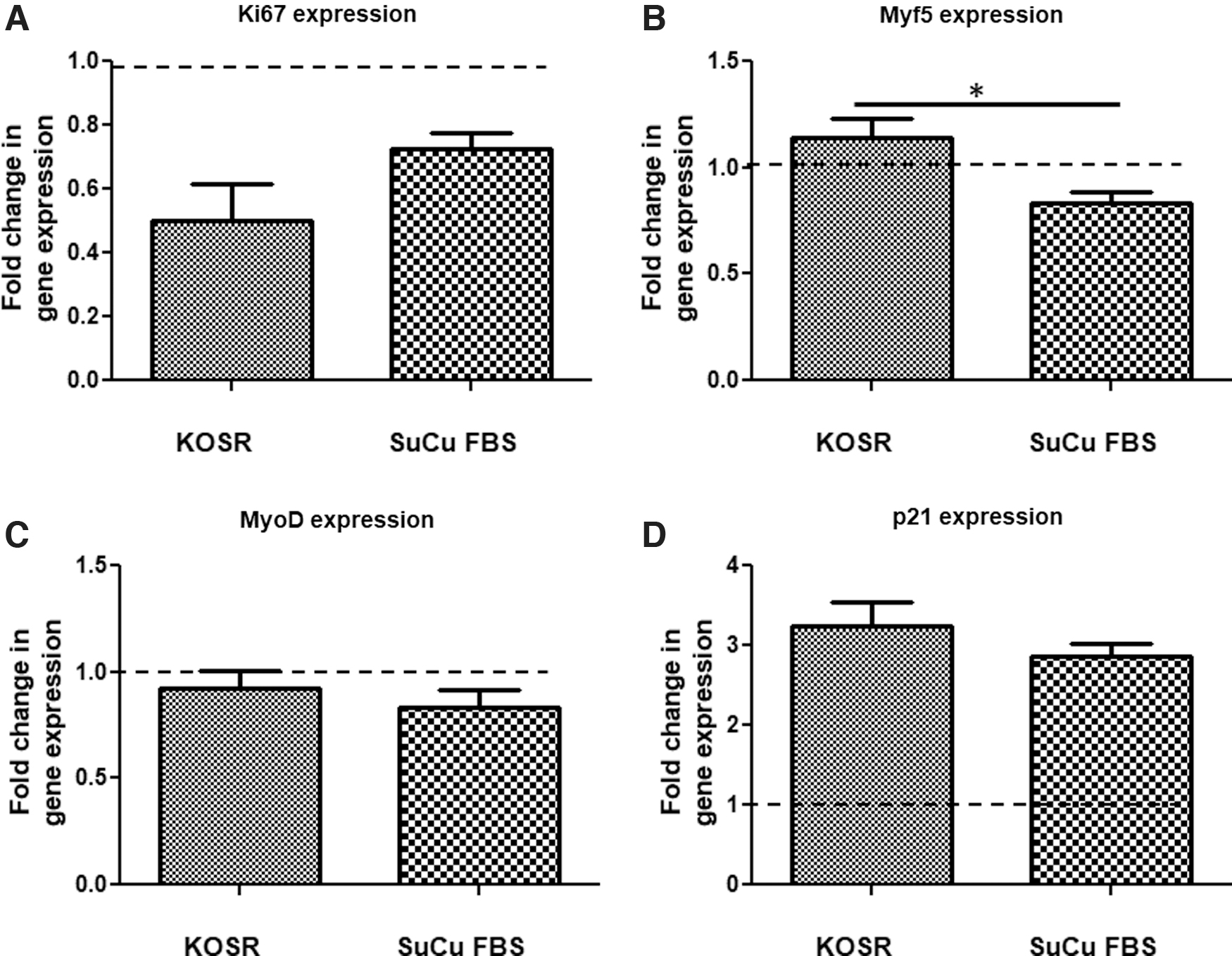
Separate experiments were then performed to evaluate the differences in PHMs responses to quiescence induction using KOSR media and SuCu. To do this, PHMs were cultured in KOSR media for 10 days and in SuCu for 2 days and harvested for several analyses. Since our data showed that all PHM clones used (KH1, KH3, and S6.3 clones) generally behaved in the same trend, when induced into quiescence, we then continued experiments using only S6.3 clone. Gene expression analysis through the use of quantitative real-time reverse transcriptase polymerase chain reaction (RT-qPCR) was performed to evaluate differences between PHMs exposed to either KOSR or SuCu conditions. The genes analyzed for this study are Ki67, Myf5, MyoD, and p21. There were no differences in the expression of Ki67, MyoD, and p21 (Fig. 4A, C, D). While Ki67 and MyoD gene expression was reduced, with KOSR method p21 showed a 3.23-fold increase and the suspension method showed a 2.86-fold increase, Myf5 expression was, however, higher in cells cultured in KOSR media compared to SuCu (Fig. 4B).
Cell cycle and molecular characteristic of PHMs exposed to the blended protocol for quiescence induction through combining SuCu and the use of KOSR-supplemented media
As shown above, the use of either SuCu- or KOSR-supplemented media was able to induce quiescence in PHMs as demonstrated by increased cells in G0/G1 phase and decrease in corresponding S-phase. Under normal culture conditions, SuCu is performed with media supplemented with FBS. We then hypothesized that a combination of the above-mentioned methods might result in cells with a better G0/G1 phenotype. Thus, for the third experiment, we induced quiescence in PHMs in vitro via culture of PHMs in suspension in normal proliferating media supplemented with FBS as control, and culture of PHMs in suspension combined with the use of KOSR-supplemented media also for 2 days as the final experimental intervention. This was followed by molecular and functional characterization of the resultant cells. Induction of quiescence (G0/G1 state) was confirmed by cell cycle analysis using flow cytometry and gene expression analysis using RT-PCR. The expression of Ki67, Myf5, MyoD, and p21 were quantified in PHMs treated with SuCu supplemented with either FBS or KOSR (Fig. 5A–D). The expression of Ki67, Myf5, and MyoD were significantly lower in PHMs cultured in suspension in media supplemented with KOSR than in media supplemented with FBS (Fig. 5A–C). However, there was no statistical difference in p21 gene expression, was an exception showing no difference in fold change, for cells cultured in suspension with FBS or KOSR (Fig. 5D).
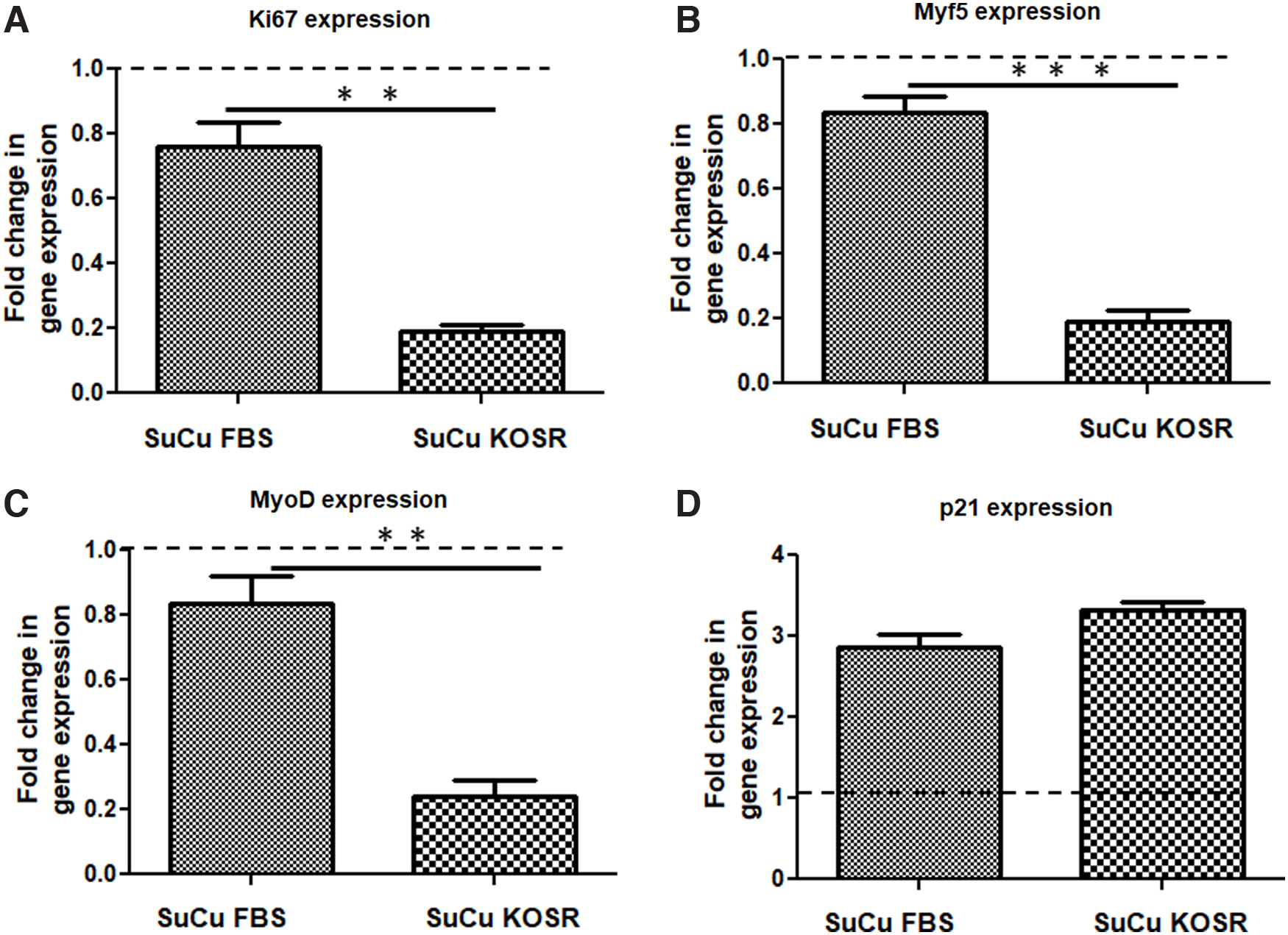
Expression of transcription factors influencing proliferation, including myogenic regulatory factors in PHMs induced into in vitro quiescence with SuCu supplemented with FBS (SuCu FBS) or KOSR (SuCu KOSR). Following induction of quiescence using 48 h treatment for both groups, cells were harvested and gene expression quantified from isolated RNA.
Our data show that a combination of both culture in suspension and use of KOSR produce PHMs with characteristics of quiescence more than the use of individual methods based on expression of MRFs and reduction of proliferative abilities. Finally, our cell cycle analysis substantiated changes in gene expression with flow cytometric analysis showing increase in cells in G0/G1 phase when cultured in suspension combined with the use of KOSR compared to cells cultured in suspension in media supplemented with FBS (Fig. 6A–C). Concurrently, cells in S-phase decreased significantly when PHMs were cultured in suspension in media supplemented with KOSR compared to SuCu in media supplemented with FBS (Fig. 6A–C). Our data show that a combination of both culture in suspension and use of KOSR produced PHMs with characteristics of quiescence more evident than with the use of individual methods based on increased cells in G0/G1 (cell cycle arrest) and less in S-phase.

Effect of SuCu with proliferation media (FBS)
Reactivation of quiescent PHMs
It is important to demonstrate the utility of any new developed protocol and in this case to show that once PHMs are induced into quiescence, they can be reactivated to reenter the cell cycle (Fig. 7). Our data show that cells in the S-phase of cell cycle decreased from 2.5% to 2% during quiescence induction and increased significantly to 5.4% upon reactivation (Fig. 8A–B). These data mean that PHMs were induced to become quiescent and lost quiescence upon culture in normal media. Cell cycle data confirm the reversibility of the G0-linked changes in cell cycle and demonstrate the utility of the combined method of quiescence induction.
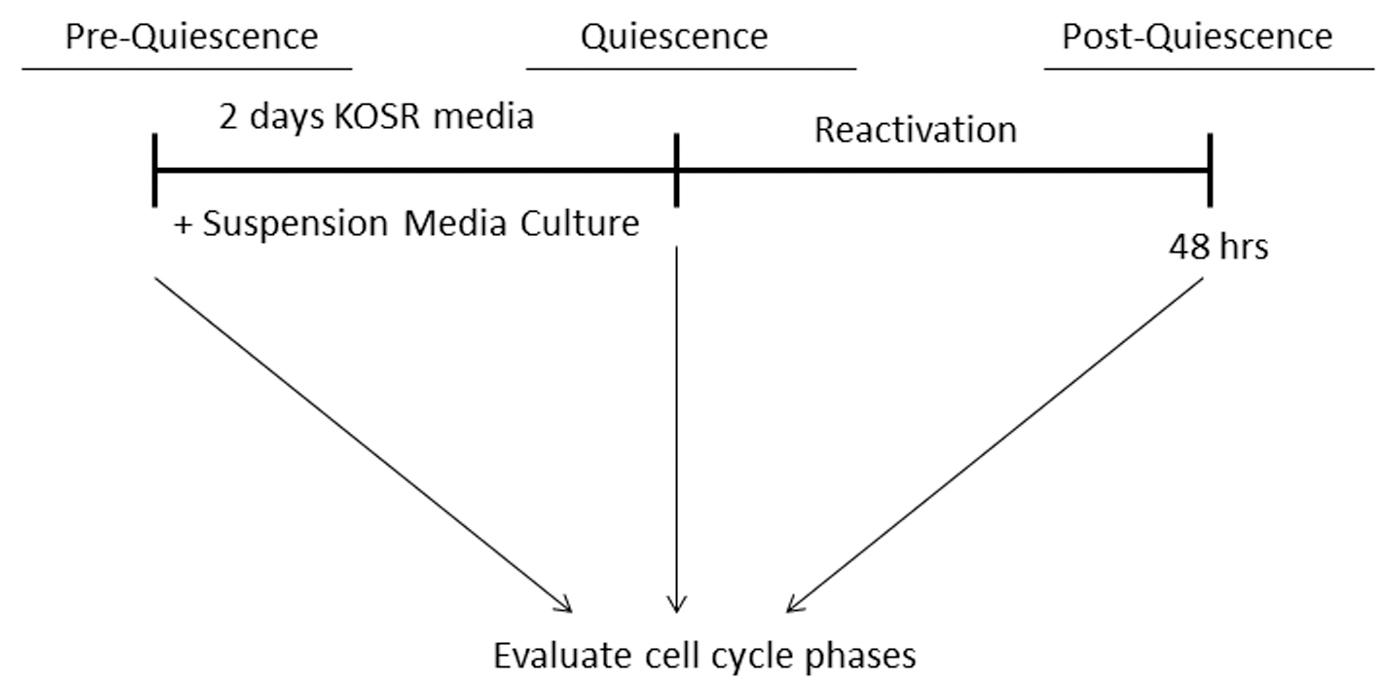
Schematic description of reactivation of quiescent PHMs to reenter the cell cycle.

Reactivation of quiescent PHMs allows cells to reenter the cell cycle and proliferate.
Discussion
Many studies have utilized immortalized cell lines such as C2C12 in their investigations of muscle cell properties and function. While these cells have yielded great amount of information, they can be genetically abnormal due to culture under in vitro conditions for a long time and therefore deviate slightly from normal function. The use of asynchronized cells in in vitro experiments can result in large standard deviations, a complication which may influence interpretation of data and hence critical aspects of control relevant to cells in vivo might be missed. Muscle stem cells' in vivo niche components include multiple protein factors. In vitro culture of cells requires the use of mitogen-controlling media such as FBS for the cells to survive and proliferate. The contents of the FBS can have different and conflicting effects on cells (Fang et al., 2017; Karnieli et al., 2017; van der Valk et al., 2018). Thus, cells cannot be synchronized through the use of FBS. These are problems that motivated the current study using PHMs.
Muscle stem cells exist in the quiescent state or G0 state in vivo (Chakkalakal et al., 2012; Fukada et al., 2007), although only a subset (Abou-Khalil et al., 2010; Baghdadi and Tajbakhsh, 2018; Buckingham, 2007). An explicit and detailed analysis of such cells in vitro is made difficult by the activation of primary cells during isolation and growth. Nonetheless, the quiescent state, characterized by a reversible cell cycle arrest (G0 state), can be induced in vitro through removing certain conditions needed by proliferating cells (Brown et al., 2017; Dumont et al., 2015; Gudagudi et al., 2020; Rumman et al., 2018; van Velthoven et al., 2017). One approach utilizes the depletion of mitogenic growth factors in the culture media (Gudagudi et al., 2020; Pardee, 1974), while another approach deprives cells of anchorage to substrate, rather suspending them in a viscous medium made up of methylcellulose (Kwon et al., 2003; Libby and O'Brien, 1983; Montagnier and Macpherson, 1964). In this study, we took advantage of both approaches individually and then simultaneously to induce cellular quiescence: SuCu and the deprivation of FBS through the use of KOSR. In addition, we probed some of the molecular mechanisms characterizing the observed changes in proportion of cells in G0 state.
We validated the use of either KOSR-supplemented media or SuCu to generate greater than 90% G0/G1 state for PHMs in culture. In particular, both methods resulted in approximately three-fold increase in mRNA of the cell cycle inhibitor, p21. While culture media with FBS is normally used in SuCu, we next modified the culture media used during SuCu by replacing FBS with KOSR. This “blended” quiescence-inducing method/protocol resulted in cells with an improved quiescence state as shown by significant reduction in the marker of proliferation, Ki67. Indeed, with the blended protocol, the Ki67 expression was decreased substantially to only 0.19-fold of control whereas suspension with FBS reduced Ki67 expression much less, only to a level of ∼0.8-fold compared to control. Finally, the promyogenic state, characterized by the myogenic regulatory factor profile, changed more with the blended protocol. In particular, both MyoD and Myf5 mRNA were reduced as low as 0.24-fold and 0.19-fold, respectively. These data provide evidence of myogenic dedifferentiation.
Myoblasts and other stem cells are known to self-renew by returning to quiescence, a process that is important for the maintenance of regenerative capacity (Cho et al., 2019; Fleming et al., 2008; Hernández-Hernández et al., 2017; Rattis et al., 2004). However, irreversible cell cycle arrest is also known as senescence (Vande Berg and Robson, 2003). Senescent cells are not responsive to growth factors (Vande Berg and Robson, 2003) and require specific gene manipulation to regain a young phenotype (Cho et al., 2019; Rumman et al., 2018). It is therefore important to consider whether experimentally induced cell cycle arrest maintains potential for reversal and reentry into cell cycle. Our previously described quiescence-inducing method involving the use of KOSR, altered the proportion of cells in the active phases of the cell cycle when compared to culture with FBS, without effect on subsequent differentiation (Gudagudi et al., 2020). Although differentiation of myoblasts is promoted by expression of the myogenic regulatory factors (Asfour et al., 2018; Hernández-Hernández et al., 2017), Myf5 plays a role in reducing the number of myoblasts going through proliferation cycles (Lindon et al., 1998; Ustanina et al., 2007). Here, KOSR exposed PHMs had slightly elevated Myf5, but in SuCu Myf5, mRNA was less than in proliferation. This appears to indicate an affinity toward a return to the parent myogenic lineage in 10 day KOSR compared to SuCu.
SuCu has been used successfully to induce G0 state in cells even when grown in culture media containing FBS (Mitra et al., 2018; Rumman et al., 2018). Mitra and colleagues described the induction of quiescence in fibroblasts through the use of SuCu as well as further manipulations of proliferating media and quiescent media to evaluate cellular regulators of quiescence (Mitra et al., 2018). Gos and coworkers demonstrated that cellular quiescence can be induced by contact inhibition in C3H10T1/2 cells (Gos et al., 2005). An important study by Eliazer and coworkers demonstrated that Wnt signaling through the involvement of Wnt4 regulates the mechanical properties of quiescent muscle stem cells (Eliazer et al., 2019). The preparation of viscous media containing methylcellulose is a relatively simple technique. However, other studies have shown that SuCu alone might not effectively inhibit proliferation and cells can only be kept in SuCu for a limited time due to exhaustion of nutrients (Amit et al., 2011; Bligny and Leguay, 1987). Here, our data show that KOSR can be used for a prolonged period and reduces proliferation and myogenic activity as effectively as SuCu. Quiescence was achieved in a synchronous manner since very high proportion of PHMs was in G0/G1 phase.
The G0 state is universal as the same characteristics are observed in cells from different animals and humans. Several reports have shown that cell cycle regulatory proteins are involved in mechanisms that induce quiescence in adult cells (Dumont et al., 2015; Mohammad et al., 2019; Pietras et al., 2011). For example, decreased RNA levels and decreased expression of cyclins is reported in quiescent mouse fibroblasts as well as human myoblasts (Sellathurai et al., 2013; Subramaniam et al., 2013). Our data show that p21 mRNA had an exceptionally high fold-change, clearly demonstrating that the major mechanism by which the SuCu and the KOSR methods induced quiescence and achieved synchronous quiescence in PHMs in vitro was by inhibition of cell cycle. Previously, it was shown that p21 is crucial in maintaining quiescence in hematopoietic stem cells and neural stem cells (Cheng et al., 2000; Kippin et al., 2005). The lowest expression of proproliferative markers and myogenic regulatory factors was in the blended protocol SuCu-KOSR. This supports our hypothesis that combining the SuCu and the KOSR methods would result in a much improved quiescence profile. Given the prominence of Myf5 and p21 in this study, further investigation of molecular signatures related to these regulators would aid in identification and manipulation of the quiescent state.
Relevance and Application
The ability to gain and maintain quiescence of PHMs in vitro has implications for our understanding of in vivo muscle stem cell biology and the utility of muscle stem cells for muscle treatments. The more closely we can simulate the in vivo environment, the more relevance in vitro research will have.
Because the immortalized C2C12 cell line is genetically abnormal and deviates slightly from normal function and because myoblasts in culture are typically asynchronized, interpretation of data and hence critical aspects of relevance to cells in vivo might be missed. Quiescence induction in PHMs results in homogenously synchronized cells.
Although muscle stem cells' in vivo niche components include multiple protein factors, media such as FBS is very high in mitogens. FBS components and their concentrations can have different and conflicting effects on cells. Despite a lack of mitogens, KOSR can be used for at least 10 days without affecting subsequent myoblast function upon replacement of mitogens. KOSR does provide basic nutrients for the myoblasts to survive.
Since p21 was the factor most associated with cell cycle arrest in G0 phase, the in vitro model we developed could be used for testing factors that might manipulate p21, which could be valuable from a potential therapeutic standpoint.
Slowly dividing myoblasts maintain self-renewal abilities in vitro while rapidly proliferating cells tend to proceed to myogenic differentiation. Such early differentiation is a risk for prolonged expansion. The blend of SuCu with KOSR could be administered if differentiation is detected too early during expansion.
Quiescent muscle stem cells are able to tolerate genetic stresses that can prevent mutations during DNA replication. A better understanding of the mechanisms controlling maintenance of quiescence may lead to strategies that could overcome regenerative inefficiencies ascribed to genetic stress, as observed in several muscle disease states and aging (Bengal and Odeh, 2019; Sampath et al., 2018).
Limitations of the Study
Other studies investigated differences between male and female donors of muscle cells and concluded that there may be slight differences in growth between these cells (Neal et al., 2012). To avoid the impact of gender and age differences on our study, we utilized cells from male donors only. It would be relevant for further studies to investigate the utility of our quiescence induction approach in cells from males and females as well as in donors of different ages.
One important future evaluation is how the observed quiescence phenotype of PHMs closely resembles that of satellite cells in vivo. This will require the use of in vivo muscle models such as animals or human studies. It is possible that gene expression and presence of specific surface markers for the G0/G1 state may be identified in PHMs and then may be used during isolation of stem cells from muscles to obtain a pure cell population. Another important issue to investigate in future is whether PHMs induced into quiescence have greater myogenic potential once restimulated, than when PHMs are stimulated in varying phases of the cell cycle. Our study does not investigate issues that may be important, including the need to recapitulate the 3D in vivo environment, including the ECM.
Conclusions
Either contact inhibition through the use of SuCu, or FBS deprivation via the use of KOSR media are efficient to induce quiescence in PHMs. Importantly, combining SuCu and KOSR media enhances the proportion of PHMs in the G0 phase of cell cycle to very high levels, alongside many-fold increase in cell cycle inhibition. Typical characteristics of proliferating, myogenic PHMs were reduced to a much greater extent using the blended protocol, but the remaining presence of Myf5 transcripts indicates that the myogenic profile is not completely removed. Our study is important for future studies evaluating and characterizing in vivo biology of PHMs and may be useful in improving the control of PHMs or satellite cells for therapeutic trials and use in regenerative medicine.
We suggest this blended new protocol can be considered in future biomedical research focused on skeletal muscle if differentiation is detected too early during the myoblast expansion. This shall also inform new ways to bridge the in vitro and in vivo divides in regenerative medicine research.
Footnotes
Author Contributions
K.B.G. and K.H.M. contributed to the study conception and design. Material preparation, data collection, and analysis were performed by K.B.G. The first draft of the article was written by K.B.G. and K.H.M. and both authors commented on all subsequent versions of the article. Both authors read and approved the final article.
Acknowledgments
The authors thank Mrs. Lize Engelbrecht and Ms. Rozanne Adams from the Central Analytical Facility at Stellenbosch University for their technical assistance with imaging and flow cytometry.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
Funding Information
This study was funded by the National Research Foundation: South African Research Chairs Initiative (SARChI). Grant number: SARCI150212114075.