
Abstract
A key lesson emerging from COVID-19 is that pandemic proofing planetary health against future ecological crises calls for systems science and preventive medicine innovations. With greater proximity of the human and animal natural habitats in the 21st century, it is also noteworthy that zoonotic infections such as COVID-19 that jump from animals to humans are increasingly plausible in the coming decades. In this context, glycomics technologies and the third alphabet of life, the sugar code, offer veritable prospects to move omics systems science from discovery to diverse applications of relevance to global public health and preventive medicine. In this expert review, we discuss the science of glycomics, its importance in vaccine development, and the recent progress toward discoveries on the sugar code that can help prevent future infectious outbreaks that are looming on the horizon in the 21st century. Glycomics offers veritable prospects to boost planetary health, not to mention the global scientific capacity for vaccine innovation against novel and existing infectious agents.
Introduction
Over the past year, the COVID-19 pandemic has challenged the technical, social, and political determinants of planetary health (Kickbusch et al., 2020; Liu et al., 2020; Mahara et al., 2020; Morhe et al., 2020; Wang et al., 2020b). Moreover, the recent history and increasing frequency of infectious outbreaks, dissolving of the boundaries between human and animal habitats, enormous loss of planetary biodiversity and the climate crises offer converging lines of evidence that the COVID-19 is likely a dry run for future ecological crises. In this expert review, we discuss the science of glycomics, unraveling of the third alphabet of life, the sugar code, and their importance in vaccine development. Glycomics offers veritable prospects to boost planetary health, not to mention the global scientific capacity to innovate therapeutics and diagnostics against novel and existing infectious agents.
COVID-19 Vaccines
COVID-19 is caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) (ICTV, 2020; Yang et al., 2020). Coronaviruses (CoVs) are enveloped single-stranded positive-sense RNA viruses that widely infect vertebrates, including humans and animals, causing respiratory and enteric diseases (Anthony et al., 2017; Heald-Sargent and Gallagher, 2012). Spike glycoprotein (S protein) plays major role in the pathogenesis of CoVs, including specific binding to host cellular receptors and host immune responses. As a result, S protein has been considered a primary target for COVID-19 vaccine development (Du et al., 2009; Liu et al., 2020).
RNA vaccines represent a new approach and were deployed for COVID-19 based on the following mechanisms so as to mimic COVID-19 infection (Chung et al., 2020). After an mRNA vaccine is injected into a person, tiny lipid vesicles containing the mRNA molecules travel through the circulatory system of the organism and merge with a special type of immune cell called antigen presenting cells (APCs) (Linares-Fernández et al., 2020). Inside these cells, the mRNA molecule connects with the ribosome, the protein making machine in the cytoplasm. The ribosome produces a foreign protein, an antigen, which is produced based on the mRNA sequence. This antigenic protein is what will ultimately stimulate the immune system. In the case of a COVID-19 vaccine, it will be the SARS-CoV-2 S protein.
The S protein antigen is presented to the immune system in one of two ways:
In the first pathway, it is broken into fragments that are bound to another protein called major histocompatibility complex (MHC)-1. Once the fragment is bound to the MHC-1 molecules, the complex leaves the endoplasmic reticulum through the secretory pathway to reach the cell surface where it is recognized by a cytotoxic T cell. This enables the cell-mediated immunity against future infections. In the second pathway, the S protein or its fragments are transported to the surface of the cell where it is released and is taken up by other APCs and degraded into fragments that form a complex with an MHC-2 protein. This complex is transported to the cell surface where it is recognized by helper T cells that help B cells make antigen-specific antibodies to neutralize the virus in any future infections. This pathway is called the antibody-mediated immune process (Fig. 1).
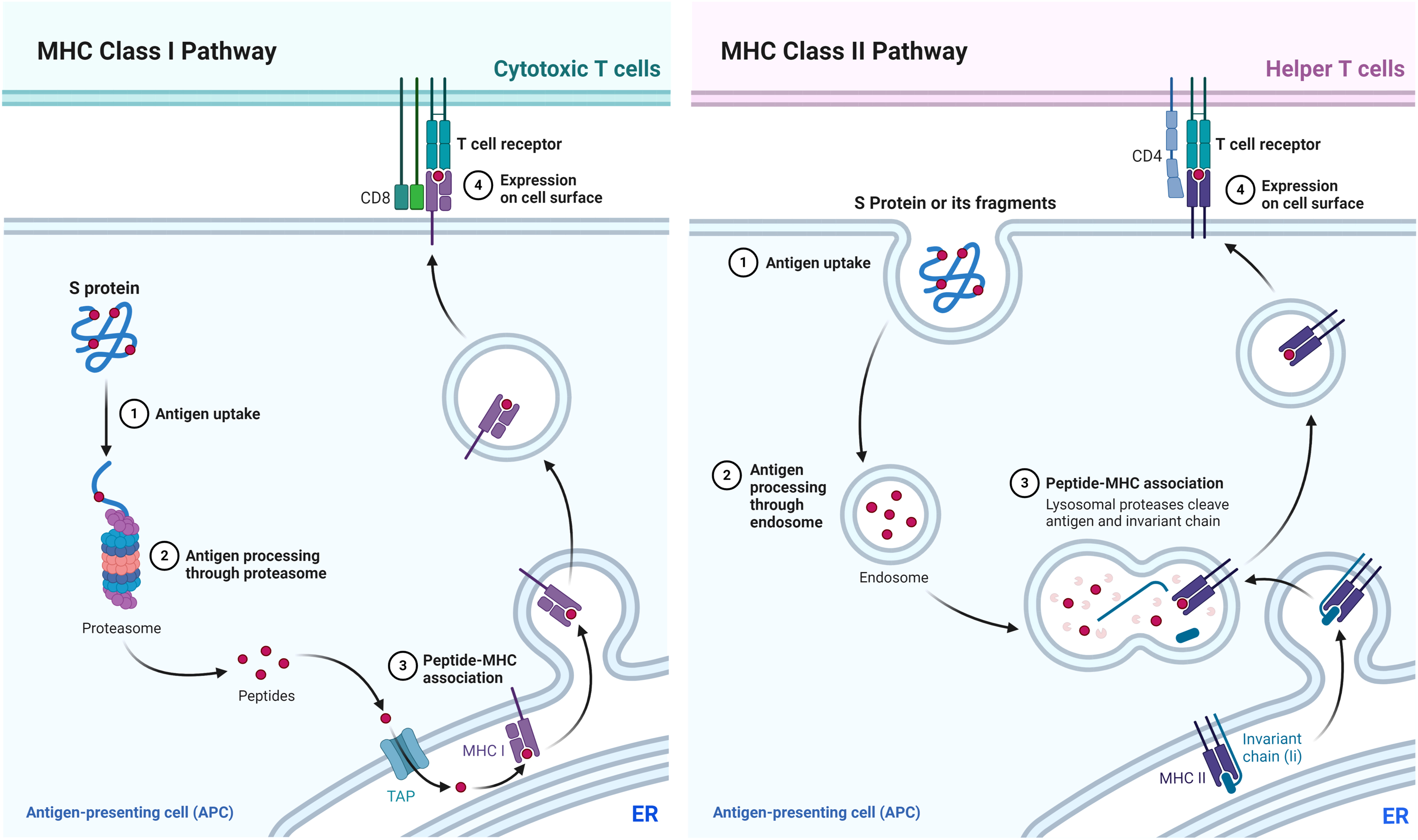
MHC class I and MHC class II pathways. APC, antigen presenting cell; ER, endoplasmic reticulum; MHC, major histocompatibility complex; TAP, transporter associated with antigen processing. Figures created with BioRender.com.
One of the advantages of the mRNA vaccine is that it stimulates both types of immunity, that is, cell-mediated immunity and antibody-mediated immunity. Also, no viral particle is present during this process, which makes mRNA vaccines considerably safer than conventional attenuated or inactivated vaccines (Table 1).
Current Severe Acute Respiratory Syndrome Coronavirus 2 Vaccine Candidate Features
Figures created with BioRender.com. Data current as of February 2021.
SARS-CoV-2, severe acute respiratory syndrome coronavirus 2.
There are >66 N-linked glycosylation sites in SARS-CoV-2, 54 of them are conserved glycosylation sites between SARS-CoV-2 and SARS-CoV (Kumar et al., 2020). In more detail, there are 22 canonical N-glycosylation sites in SARS-CoV-2 S protein. Eighteen of them are conserved with SARS-CoV S protein (Zhang et al., 2020b). The highly glycosylated modification in SARS-CoV-2 S protein allows the virus to have multiple mutations, which is one of the main obstacles for vaccine development (Vigerust and Shepherd, 2007). Glycan shields also “hide” immunogenic peptides, leading to the low immunogenicity of the S protein, thereby evading neutralizing antibodies that prevent virus adhesion to host cellular surface receptors (Walls et al., 2016; Zhang et al., 2020b; Wrapp et al., 2020).
By the end of 2020, the SARS-CoV-2 vaccines reported to be at various stages of development (Koirala et al., 2020) are divided into two major groups: (1) vaccines prepared from virus replicating in cell lines, such as live attenuated viruses and inactivated viruses; (2) vaccines composed of nucleic acid (DNA and RNA) and viral vectors or nanoparticles that deliver the gene or mRNA of SARS-CoV-2 S protein.
Both groups possess their own advantages. A great advantage is that they can be engineered for increasing their immunogenicity for Group 1 vaccines. Whereas the ability to produce vaccines in short time is the major advantage of DNA and RNA vaccines in Group 2 (Table 1). Since the immunizing antigens of Group 2 vaccines are produced within the vaccinated individuals, engineering of the protein component of the vaccine is not feasible (Koirala et al., 2020).
Glycoengineering the glycan shield on viral vaccines for its conversion from protein masking glycan into glycan that effectively targets vaccines to APCs will markedly increase the immunogenicity of viral vaccines (Galili, 2020a). This effective targeting for the extensive uptake of viral vaccines by APCs is achieved by harnessing the natural anti-galactose (Gal) antibody, which is abundant in all humans who are not severely immunocompromised. The α-1,3-galactose glycan (α-Gal) epitopes of vaccine/CoVs is the ligand for the human natural anti-Gal antibody, which constitutes ∼1% of human immunoglobulins. Glycoengineering of glycan shields will present α-Gal epitopes for the human natural anti-Gal antibody, thereby amplifying the vaccine efficiency (Chen, 2020; Galili, 2020a).
Methods for SARS-CoV-2 Vaccine Glycoengineering
Glycans at the N-linked glycosylation site on S protein are synthesized by the relevant cellular enzymes in host cells (Galili and Swanson, 1991; Galili et al., 1996; Galili, 2020a; Repik et al., 1994). Previous studies show that enveloped viruses replicating in human cells lack α-Gal epitopes, because nucleotide mutations inactivated the enzyme α1,3 galactosyltransferase (α1,3GT) in human cells (Galili and Swanson, 1991; Galili et al., 1996; Galili, 2020a; Repik et al., 1994). However, because of the stimulation by gastrointestinal bacteria carrying galactose, anti-Gal antibodies are abundant in humans, constituting ∼1% of serum immunoglobulins (Hamadeh et al., 1995; Hamanova et al., 2015). Human anti-Gal antibodies help the immune system to eliminate many kinds of zoonotic viruses carrying galactose synthesized by the source animals (Galili and Swanson, 1991; Galili et al., 1996; Galili, 2020b; Repik et al., 1994). Although anti-Gal antibodies are abundant in human bodies, their activity varies among individuals (Buonomano et al., 1999; Galili, 2020a). Meanwhile, immunization with antigens carrying galactose could result in rapid elevation of anti-Gal antibodies titer, which is beneficial to mediate APCs to destroy zoonotic viruses, including SARS-CoV-2 (Breiman et al., 2020; Galili, 2020a). Zoonotic SARS-CoV-2 S protein carries α-Gal, which can bind to anti-Gal antibodies in humans. Glycoengineering on SARS-CoV-2 vaccines to present more α-Gal epitopes will increase anti-Gal-mediated amplification of vaccine immunogenicity. Some of the reference methods for glycoengineering SARS-CoV-2 S protein vaccine to present more α-Gal epitopes are listed as follows.
Synthesis of α-Gal epitopes by recombinant α1,3GT
The glycotransferase, α1,3GT, links the galactose to the glycans of disialylated glycoproteins to form α-Gal epitopes (Fig. 2). The formation of multiple a-gal epitopes by rα1,3GT was also observed on human tumor cells (Chen et al., 2001; Furukawa et al., 2017; Galili et al., 2003), on influenza virus (Abdel-Motal et al., 2007; Chen et al., 2001), and on gp120 of human immunodeficiency virus (Abdel-Motal et al., 2006, 2010; Colomb et al., 2019). In these studies, α-Gal epitopes were synthesized and shown to be effectively targeted by anti-Gal to autologous dendritic cells.
Glycoengineering of α-Gal epitopes on S protein of SARS-CoV-2. Left chain: glycans of the complex type on viral envelope glycoproteins are synthesized on asparagine (N) in amino acid sequences of asparagine-X-serine/threonine (N-X-S/T-). Many of these carbohydrate chains of the S protein are capped by SA. Center chain: SA is removed from the glycan chain by sialidase to expose the penultimate Galβ1-4GlcNAc-R, called N-acetyllactosamine (LacNAc). Right chain: Incubation of inactivated virus or soluble S protein carrying the disialylated glycan, with recombinant α1,3galactosyltransferase (rα1,3GT) leads to synthesis of α-Gal epitopes on the glycans of vaccines. These epitopes would bind the human natural anti-Gal antibody at the vaccination site and generate immune complexes which are targeted for the extensive uptake by human APCs. Figures created with BioRender.com. α-Gal, α-1,3-galactose glycan; SA, sialic acid.
Intracellular synthesis of α-Gal epitopes on SARS-CoV-2
Synthesis of α-Gal epitopes on SARS-CoV-2 could be achieved within cells where the virus is replicating. Studies suggest that some of the glycans on SARS-CoV-2 propagated in cells containing active α1,3GT will be synthesized with α-Gal epitopes (Galili and Swanson, 1991; Galili et al., 1996). Researchers indeed find that transfection with the α1,3GT gene enables cells lacking α-Gal epitopes start synthesizing this epitope on cell surface glycans (Rossi et al., 2005). In view of vaccine development experience for influenza virus, the efficacy of targeting vaccines to APCs by anti-Gal might be directly proportional to the number of α-Gal epitopes on inactivated SARS-CoV-2 or on S protein subunit vaccines (Ambrozaitis et al., 2009; Szymczakiewicz-Multanowska et al., 2009). Maximizing α-Gal epitope synthesis on a virus such as SARS-CoV-2 replicating in a host cell line can be achieved by elevating the activity of α1,3GT, decreasing sialyltransferase activity, or both. In addition, an alternative method that enables introduction of multiple copies of the α 1,3GT gene in host cells for SARS-CoV-2 is transduction of such cells with replication defective adenovirus containing the α1,3GT gene (AdaGT) (Deriy et al., 2002), before infection of the cells with replicating SARS-CoV-2.
Synthesis of recombinant S proteinα-Gal in glycoengineered yeasts and bacteria
Yeasts are considered an effective expression system for production of recombinant proteins. However, production of therapeutic glycoproteins for injection into humans is limited by the ability of yeasts to synthesize N-linked glycans that are only of the high mannose type. Therapeutic glycoproteins with high mannose N-linked glycans have short half-life in humans. To overcome this limitation, yeasts have been glycoengineered to synthesized humanized glycans that resemble the complex structure of the left glycan in Figure 2 by introducing the corresponding glycosyltransferase genes into an yeast expression system (Choi et al., 2003; Wildt and Gerngross, 2005). It is possible that introducing the α1,3GT gene (GGTA1) into the yeast will result in synthesis of glycans capped with α-Gal epitopes on recombinant S protein and on other recombinant glycoproteins produced in this expression system. In addition, Bacteria such as Escherichia coli may also serve as expression system for production of recombinant S proteinα-Gal (Fisher et al., 2011).
Cellular Ecology: Interactions of Glycans Between Hosts and Viruses
Besides forming shields covering SARS-CoV-2 S protein, glycans are also a nexus for interactions between host cells and different viruses (Flint et al., 2008). Glycans possess the potential to induce immune tolerance, which raising the conflict in evolutionary competitions between human and viruses. CoVs exercise the use of glycan shielding and conformational masking of S protein to circumvent the host immune cells (Kaur and Gupta, 2020). Hosts also use glycans to defend themselves against pathogens. After virus invasion, the proliferation of the virus and disease progression depend on the interaction between the virus and the host cell receptor. On the one hand, for viral pathology, the host cell surface glycoproteins can promote viral infection. For example, the sialic acid component on the surface of the host cell could be easily recognized by rotavirus and influenza virus (Connor et al., 1994). On the other hand, the modification of host protein glycosylation can sometimes inhibit viral infection. A viral restriction factor, bone marrow stromal cell antigen 2 (BST-2), is an interferon-induced glycosylated protein, which is mainly located in the cell membrane. BST-2 glycosylation process protects the host cells from viral infections by blocking the viruses in cell multivesicular body (Lv et al., 2015). In addition, most animal epithelia secrete a heavily O-glycosylated glycoprotein, mucin (Cohen et al., 2013). This defensive glycoprotein resists proteolytic degradation and form hydrated gels to generate a physical barrier against invading microbes (Cohen et al., 2013). Mucins also serve as decoys, mimicking the cell surface receptor structure, protecting host cells from recognition by viruses. Correspondingly, many viruses evolve to express sialidases to bypass sialic acid-rich mucin decoys. Glycans mediate aspects of ecology that promote evolution rapidly, their roles in ecology and evolution are pivotal (Springer and Gagneux, 2016).
Glycomics Integration with Multiomics Platforms to Prevent Future Crisis
Glycomics in a context of multiomics helps triangulate biological data across the cellular biology hierarchies from genes to glycans as well as offering both structural and functional context in cell biology (Ly et al., 2010). Based on glycomics, new scholarships including systems glycobiology and personalized glycomedicine that collectively aspire to explain the role of glycans in intra- and interindividual variations in disease susceptibility and response to health interventions such as vaccines (Kunej, 2019). Systems glycobiology is a subfield of systems biology, established on combining multiple glycomics data sets with multiomics data by glycoinformatics (Bennun et al., 2016). Systems glycobiology is a holistic approach to disclose complex glycosylation process and relations among its components.
Glycosylation plays an important role in normal physiology as well as pathological development (Varki, 2017). Different glycosylation patterns could distinguish between health status and diseases, functioning as stratification biomarkers for personalized medicine (Reily et al., 2019). The integration of glycomics with multiomics biomarkers provides novel prospects for disease diagnosis and prevention innovation. For example, multivariate models consisting of different omics data (proteomic, metabolomic, genomic, epigenomic, and glycomic) from previous studies have been shown to improve accuracy for risk stratification of dementia, metabolic syndrome, type 2 diabetes mellitus, cardiovascular diseases, and aging (Adua et al., 2018, 2019; Ge et al., 2018, 2019; Liu et al., 2018, 2019; Murphy et al., 2018; Wang et al., 2016, 2019; Yu et al., 2016; Zhang et al., 2020a, 2021; Zhao et al., 2018).
When we treat COVID-19 as a signal from the nature, it might serve as a warning that single strategy (such as imaging diagnosis and clinical medication) against contagious diseases could not functionate individually. A systematic map of all possible connections between bioscience multiomics techniques and other discipline advantages such as the future social policy optimization will enable a superior global pandemic control system. For the past year, global scientific research and development in response to the pandemic has been unprecedented in terms with the academic publication and vaccine development. Besides science as the considerable victory, another remarkable success would be if the world can collaborate in global solidarity to ensure equitable access to the vaccines, therapeutic methods and materials, and diagnostics required to terminate the pandemic (Ihekweazu, 2021). The SARS-CoV-2 is rapidly mutating (Wang et al., 2020a). There have been three variant strains emerging that possess greatly increased viral transmissibility (ECDPC, 2021). More cases have been reported in Southern Africa (ACSS, 2021). The low-income countries may again be forced into strict lockdowns in response to the emerging cases, which would lead to crucial disruption to education and economics. The inequality in access to vaccines would exacerbate the challenges. Therefore, there is urgent requirement for rapid new investments in science and development to remain ahead of the natural mutation that selects the variant viral strains. At this juncture, countries are obliged to work together to overcome the adversity. Each individual country could not expect to fight against the pandemic threat in a vacuum, either end a global crisis at national borders. For COVID-19 and other contagious diseases, the vaccines, therapeutics, and diagnosis need to be considered as a common public benefit and not only as business prospects. Solidarity and collaboration are our best approaches to control the acute phase of COVID-19 pandemic (Ihekweazu, 2021; Springer, 2020).
Outlook
With the emergence of the novel SARS-CoV-2, planetary health needs ongoing and future action in preventive medicine and for pandemic proofing against future ecological crises. A major hope in large planetary scale infectious disease crises is the new candidate vaccines and potential antivirals. The transmembrane spike (S) glycoprotein of CoVs, which facilitates the virus's entry into the host cells, is covered with N-linked glycan shields. S glycoprotein is known as the main target of antibodies having neutralizing potency and is also considered as an attractive target for therapeutic or vaccine development. Glycoengineering the vaccine glycan shields to present α-Gal epitopes will increase the binding with human anti-Gal antibody, therefore amplifying the SARS-CoV-2 vaccine immunogenicity. The amplification is achieved by targeting anti-Gal/SARS-CoV-2α-Gal or S proteinα-Gal for extensive uptake by APCs through immunoglobulin Fc/Fcγ receptors interaction. This amplified immune response would contribute to considerably higher titers of neutralizing antibodies and increased T cell response against cells infected by SARS-CoV-2. Glycoengineering of inactivated SARS-CoV-2 or S protein vaccines to present multiple α-epitopes is feasible by use of sialidases, recombinant α1,3GT, and by propagating the virus in cell lines that contain high α1,3GT activity. In addition, yeast or bacterial expression system also enables production of recombinant S protein with multiple α-Gal epitopes.
The interactions between host and different viruses involve the participation of glycans. Glycans can respond to demands that change on physiological, developmental, ecological, and evolutionary timescales. Unlike protein whose formation follows the transcription and translation of genes (i.e., the central dogma), glycans are formed without requiring a template and their synthesis involve multiple enzymes that add or subtract monosaccharide units to proteins. Thereby the glycosylation process, the sugar code, is associated with the concept of paracentral dogma (Ma et al., 2018; Özdemir et al., 2020; Wang, 2019). As a result, glycans are often recruited to enact dynamic traits. Glycans are tractable molecules whose genetic and environmental determinants can be recognized and whose ecological relevance can be understood. Quantifying and predicting the synthesis, regulation and function of glycans will allow researchers to study aspects of ecology and evolution that are not accessible to studies of proteins and genetic materials alone.
Footnotes
Acknowledgments
The authors thank Australia–China International Collaborative Grant.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
Funding Information
The authors thank Australia–China International Collaborative Grant (NHMRC APP1112767-NSFC 81561128020). X.Q.W. was supported by China Scholarship Council (201608230108).