
Abstract
A global rise in life expectancy comes with an increased burden of serious life-long health issues and the need for useful real-time measures of the aging processes. Studies have shown the value of biochemical signatures of immunoglobulin G (IgG) N-glycosylation as clinically relevant biomarkers to differentiate healthy from accelerated aging. Most human biological processes rely on glycosylation of proteins to regulate their function, but these events appear sensitive to environmental changes, age, and the presence of disease. Specifically, variations in N-glycosylation of IgG can adversely affect inflammatory pathways underpinning unhealthy aging and chronic disease pathogenesis. This expert review highlights the discrepancies between an organism's age in years of life (chronological age) versus age in terms of health status (biological age). The article examines and synthesizes the studies on IgG N-glycan profiles and the third alphabet of life, the sugar code, in relation to their relevance as dynamic indicators of aging, and to differentiate between normal and accelerated aging. The levels of N-glycan structures change with aging, suggesting that monitoring the alterations of serum glycan biosignatures with glycomics might allow real-time studies of human aging in the near future. Glycomics brings in yet another systems science technology platform to strengthen the emerging multiomics studies of aging and aging-related diseases.
Introduction
Aging is a multifactorial interconnected biological process, which manifests as declining physiological functions and increasing susceptibility to chronic inflammation and to aging-related diseases (Kennedy et al., 2014; Lopez-Otin et al., 2013). Studies have suggested that healthier lifestyle such as healthy diet, continued physical exercise, restriction on alcohol drinking, reducing sitting time, and smoking cessation may slow the appearance and/or progression of aging (Blair et al., 1989; Franceschi et al., 2018; Kovell et al., 2020; Reid et al., 2018; Vopat et al., 2014).
With these observations, extensive effort has been made during the past decades to identify biomarkers of aging, which may predict or monitor age-associated physiological decline and disease. The American Federation for Aging Research (AFAR) has summarized nine “hallmarks of aging,” including
epigenetic alternations (e.g., methylation, phosphorylation, ubiquitination, glycosylation),
loss of proteostasis,
deregulated nutrient sensing,
mitochondrial dysfunction,
cellular senescence,
stem cell exhaustion,
altered intercellular communication,
genomic instability, and
telomere attrition (Lopez-Otin et al., 2013).
Among these hallmarks, recent advances in high-throughput technologies to study the sugar code, that is, the third alphabet of life, converged into protein glycosylation as an essential biomarker of aging and aging-related disease (Koga et al., 2011; Kristic et al., 2014; Moreno and Aldea, 2020; Yu et al., 2016). According to the AFAR, “aging biomarkers” need to meet the following criteria (Butler et al., 2004; Johnson, 2006):
it must predict the physical and mental status of a person in their life span; the effects of disease must be excluded; it must be harmless and tested repeatedly in humans; it can be tested in vivo and in vitro.
No such marker has been identified to date that addresses all these dimensions although studies of glycans offer new insights. Thus, in this expert review, we focus on the role of protein glycosylation in aging progress (especially in inflammatory process) and aging-related diseases.
Protein glycosylation is an enzymatic process responsible for the attachment of carbohydrates (glycans) to glycoproteins, predominantly through nitrogen (N) and oxygen (O) linkages. Glycosylation occurs in the endoplasmic reticulum/Golgi compartment of essentially all cells being mediated by the coordinated action of a portfolio of different glycosyltransferase and glycosidase enzymes (Alavi and Axford, 2006; Varki, 2006). The glycan is a significant part of nearly all cellular membrane and extracellular proteins, and of them, more than a half are glycoproteins (Apweiler et al., 1999; Ozdemir et al., 2020). The structure and/or composition of glycan attached to protein backbones plays an important role in nearly all molecular interactions at the cell surface and in the intercellular space during the aging progress (Packer et al., 2008).
Different types of glycans can be synthesized with varying enzymes from varying number, order, and type of monosaccharide units. The most abundant monosaccharides that can be found are: fucose, galactose, glucose, mannose, N-acetylgalactosamine, N-acetylglucosamine (GlcNAc), sialic acid, and xylose (Lauc et al., 2014). The extremely complex branched structures of glycan make the analysis of glycosylation significantly more challenging than the analysis of linear DNA and protein sequences. However, with the rapid development of biological techniques, such as hydrophilic-interaction chromatography, high-performance liquid chromatography, capillary electrophoresis, lectin affinity, and mass spectrometry, research on N-glycan of plasma/serum protein and immunoglobulin G (IgG) has experienced an unprecedented advance over recent years (Adamczyk et al., 2012; Wang, 2019; Wang and Ozdemir, 2019).
Protein Glycosylation in Aging Progress
Diverse plasma glycans were shown to be closely related to aging over three decades ago (Parekh et al., 1988) and have also been replicated in recent studies in European (Valerie et al., 2008; Vanhooren et al., 2007, 2009) and Chinese populations (Lu et al., 2011). In a human serum glycoproteins study, the level of the agalactosylated glycans is increased with age, whereas the core alpha-1,6-fucosylated bi-galactosylated biantennary structure decreased, and the similar variation trend has been shown in the Werner syndrome (Vanhooren et al., 2007). These observations suggest that change of N-glycan level can provide a noninvasive surrogate marker of aging (Vanhooren et al., 2007). The levels of N-glycan structures are changed with aging, indicating that examining alterations of serum glycan fingerprint might be suitable for monitoring the progress of aging in real time (Vanhooren et al., 2009).
The changes in plasmatic forms of two glycosyltransferases, beta-1,4-galactosyltransferase 1 (B4GALT1) and beta-galactoside alpha-2,6-sialyltransferase 1 (ST6GAL1), in aging progress also prove the strong association between glycosylation and aging, although the precise mechanism is still unknown (Dall'Olio, 2018). The selective combinations of IgG N-glycan structures associate with biological hallmarks of prechronic disease states when biological age precedes chronological age. The usefulness of IgG N-glycan profiles as indicators of aging can be achieved by profiling IgG N-glycosylation retro- and prospectively in real time to longitudinally capture the influences of genetics, environment, and time on glycome.
Immunoglobulins (Igs) are a group of plasma glycoproteins (IgG, immunoglobulin M, immunoglobulin A, immunoglobulin E, and immunoglobulin D) with antibody activity accounting for ∼20% of plasma proteins (Junqueira and Carneiro, 2003). Among them, IgG is the most abundant (over 80%) Ig and is the main component of antibody formation after a secondary immune response. IgG is considered as a perfect model glycoprotein because of its well-defined glycan structures and functions (Gornik et al., 2012). IgG molecules consist of two identical light chains and heavy chains, which are joined by covalent bonds to form a “Y-shaped” structure (Huber et al., 1976) (Fig. 1). The two domains of IgG that are responsible for their in vivo properties are the Fab (antigen binding fragment) and the Fc (crystallizable fragment) (Schroeder and Cavacini, 2010). A single, N-linked glycosylation conservative site exists at the amino acid 297 (Asn297) in the heavy chain of all IgG subclasses (Fig. 1). On average, 2.8 glycans are linked to each IgG molecular and among them two glycans are located at Asn297 in the Fc region (Arnold et al., 2007).
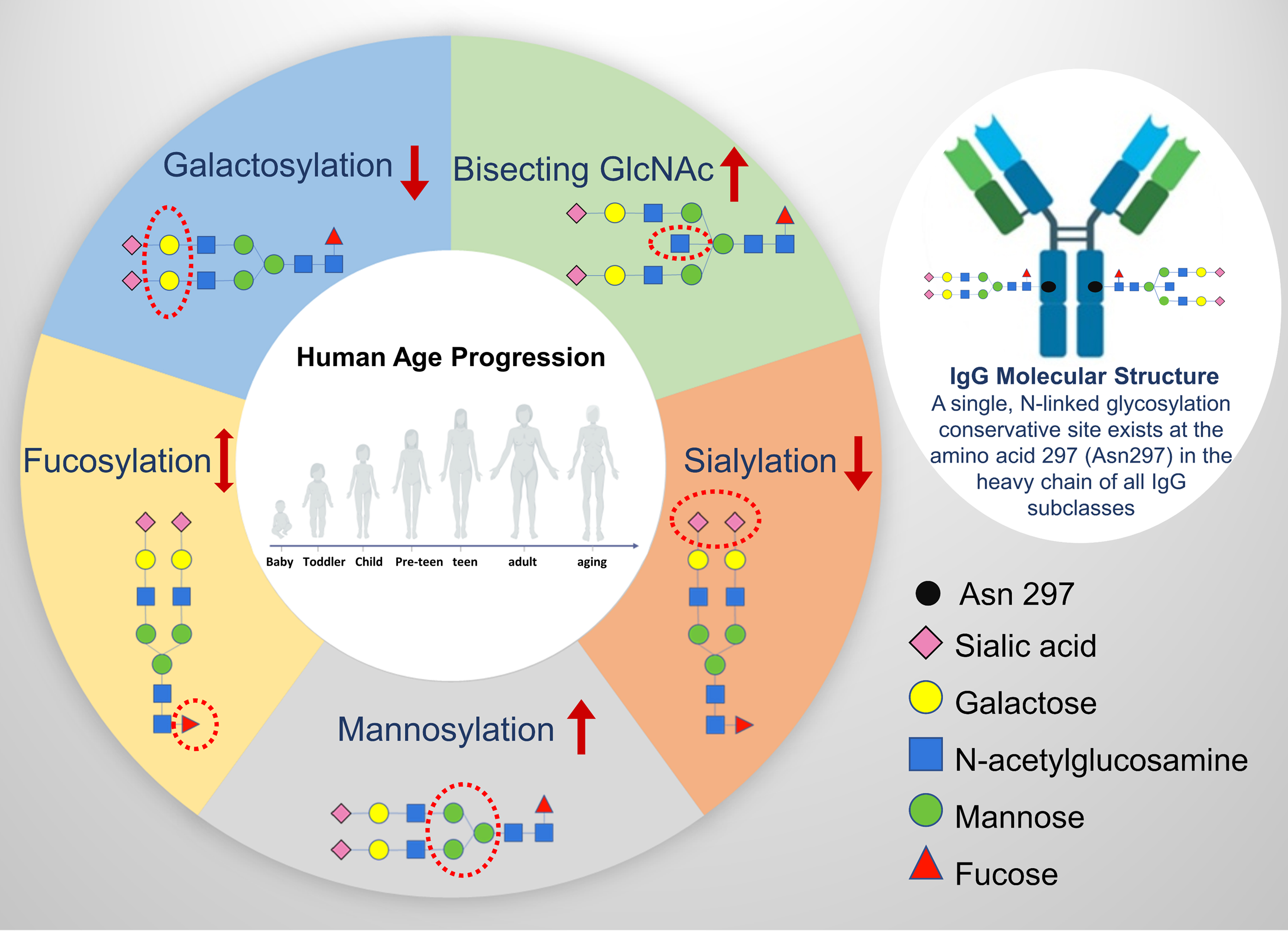
IgG glycosylation in aging progress. The structural schemes of IgG N-glycans are shown in terms of sialic acid (purple rhomb), galactose (yellow circle), N-acetylglucosamine (blue square), mannose (green circle), fucose (red triangle) for visual representation. The red dotted circle highlights the mainly involved glycan domain during the relevant glycosylation progress. The decreasing galactosylation and sialylation while increasing bisecting GlcNAc contents and mannosylation are found in the majority of aging studies. The changes in levels of fucosylation remain contested and need further research. GlcNAc, N-acetylglucosamine; IgG, immunoglobulin G.
Around 30% of IgG and some monoclonal antibodies contain N-linked glycosylation sites in Fab and the hinge region, but they are not conserved (Jefferis, 2012; Wright and Morrison, 1997). The Fc region of glycans generally contains a core heptasaccharide, which is species, tissue, age, and sex specific, and this specificity is conferred by the terminal oligosaccharides (Huhn et al., 2009). Normal structure of IgG N-glycans in the Fc region is complex fucosylated biantennary oligosaccharides, and the main sources of glycan heterogeneity are the galactosylation and/or sialylation in the residues. In the Fab region, glycans are usually sialylated and with a bisecting GlcNAc residue (Wormald et al., 1997). Genetic recombination and/or somatic mutation can affect the process of IgG synthesis and result in the variation of N-linked glycosylation sites in the heavy and light chains of the IgG molecule. cDNA sequential detection shows that 9% of the variable regions of IgG contain potential N-linked glycosylation sites and the majority of the variable regions are located in the heavy chain of IgG (Zhu et al., 2002).
Fine changes in attached glycan composition can dramatically alter the conformation of the Fc domain with significant consequences for effector functions of IgG (Gornik et al., 2012). The addition of terminal sialic acid to this glycan reduces Fcγ receptor (FcγR)-binding and converts IgG antibodies from proinflammatory to anti-inflammatory mediators through the lectin receptor SIGN-R1 or DC-SIGN (Dendritic Cell-Specific Intercellular adhesion molecule-3-Grabbing Nonintegrin), which upregulates surface expression of the inhibitory Fc receptor, FcγRIIb, on inflammatory cells, thereby attenuating autoantibody-initiated inflammation (Anthony and Ravetch, 2010). Studies have identified that decreased levels of bisecting GlcNAc glycoforms of IgG are associated with human longevity in Caucasians (Ruhaak et al., 2010); whereas the level of galactosylation of IgG decreased in aging process in Asian Japanese (Shikata et al., 1998). The aging-associated IgG glycosylation changes are often observed in the chronic inflammatory state (Dall'Olio, 2018), which makes IgG glycosylation an attractive biomarker in monitoring aging-related progress. The main domains of IgG glycan involved in the aging progress are demonstrated in Figure 1.
Protein Glycosylation and Aging-Related Diseases
A large variability of human plasma N-glycans was observed in a population-based study (Knezevic et al., 2009). However, the N-glycan structure are considerably stable within a short time-frame, indicating that changes of N-glycan profiles may the consequences of environmental determinants and/or pathophysiological situations (Olga et al., 2009). A population-based study provided preliminary evidence of an association between the variation of plasma N-glycans and genetic and environmental factors by testing the whole plasma N-glycans (Knezevic et al., 2009; Zaytseva et al., 2020). Other studies with a population focus showed an association between several plasma N-glycan levels (trigalactosylated glycans, biantennary glycans, triantennary glycans, core-fucosylated glycans, monosialylated, disialylated, and trisialylated glycans) and metabolic syndrome-related risk factors (systolic blood pressure [SBP], diastolic blood pressure [DBP], body mass index, fasting plasma glucose, age, and gender) (Lu et al., 2011). Therefore, glycosylation alterations are suggested to be associated with a number of aging-related diseases (Alavi and Axford, 2008).
Malfunction of IgG glycosylation appears to be associated with the incidence and development of many aging-related diseases, such as inflammatory bowel disease (IBD) (Akma et al., 2015), hyperuricemia (Hou et al., 2019), type 2 diabetes mellitus (T2DM) (Liu et al., 2019), hypertension (Wang et al., 2016), rheumatoid arthritis (RA) (Parekh et al., 1985; Sebastian et al., 2016; Troelsen et al., 2012), systemic lupus erythematosus (SLE) (Huang et al., 2003; Sjowall et al., 2015; Vuckovic et al., 2015), Alzheimer's disease (AD) and progressive mild cognitive impairment (Lundström et al., 2014), Parkinson's disease (PD) (Russell et al., 2017), dementia (Zhang et al., 2021), and cancer (Adamczyk et al., 2012; Lan et al., 2016; Wang, 2013).
A study found that the levels of glycosylated IgG are significantly higher in diabetic patients compared with normal controls, and such modification in vivo was associated with IgG functional alteration (Kaneshige, 1987). The glycation levels of IgG were increased in poorly controlled T2DM patients when compared with healthy subjects and also well controlled diabetic patients (Lapolla et al., 2000). In a multiethnic study (China, Croatia, and Scotland), five glycans (IgG with digalactosylated glycans) significantly differed in participants with prehypertension or hypertension compared with those with normal blood pressure, whereas additional 17 glycan traits only significantly differed in participants with hypertension compared with those of normal blood pressure (Wang et al., 2016). These glycans were also significant correlated with SBP or DBP (Wang et al., 2016). IgG hypogalactosylation correlated with inflammation-associated molecules, such as interleukin-6 (IL-6), and C-reactive protein in RA patients (Troelsen et al., 2012).
Different distributions of asparagine-linked biantennary complex-type oligosaccharide structures of IgG were found between normal individuals and patients with RA (Troelsen et al., 2012) and patients with osteoarthritis (Parekh et al., 1985). A study on RA demonstrated that the agalactosylated IgG glycan due to its exposure of the mannose-rich core glycan structure acquires the capacity to bind to mannose-binding lectin, providing an explanation for this enhanced proinflammatory activity in vivo (Malhotra et al., 1995). The IgG differences between the normal individuals and the patients indicate that glycosylation is involved in the pathogenesis of the diseases (Parekh et al., 1985). In addition, disease activity of SLE patients correlated with higher exposure of Aleuria aurantia lectin-reactive fucosyl residues, an immobilized IgG complex (Sjowall et al., 2015). In another SLE study, the significant changes include decreased galactosylation and sialylation of IgG, as well as decreased core fucose but increased bisecting GlcNAc of IgG, indicating that changes in IgG glycosylation may be applied as a dynamic biomarker to the monitoring of SLE progress (Vuckovic et al., 2015).
A lower abundance of complex galactosylated and sialylated IgG glycan structures was found in AD patients (Lundström et al., 2014). Furthermore, IgG glycan profiling from AD patients with and normal controls suggested that there may be a reduced capacity for the IgG to inhibit Fcγ-RIIIa binding, which would allow an increased ability for the IgG to cause antibody-dependent cell cytotoxicity (ADCC) and a possible state of low-grade inflammation in individuals with PD (Russell et al., 2017). In addition to the above diseases, glycosylation of whole plasma/serum protein is also found to be a potential biomarker for liver, pancreatic, prostate, ovarian, breast, lung, and stomach cancers (Adamczyk et al., 2012; Wang, 2013). In most of these cancers, the fucosylation and sialylation expressions are found to be significantly modified (Adamczyk et al., 2012; Wang, 2013).
These studies suggest that the individual variation in N-glycosylation of IgG contributes to the pathogenesis of aging and aging-related disease, presumably through its effect on pro- and/or anti-inflammatory, ADCC, and complement-dependent cytotoxicity (CDC) pathways. The addition of terminal sialic acid to glycan reduces FcγRs converts IgG antibodies from proinflammatory to anti-inflammatory mediators through the lectin receptor SIGN-R1 or DC-SIGN, which upregulates surface expression of the inhibitory Fc receptor FcγRIIb, on inflammatory cells, thereby attenuating autoantibody-initiated inflammation (Anthony and Ravetch, 2010). As the pervasive feature of aging tissues and nearly all age-related diseases are featured by chronic inflammation (Franceschi and Campisi, 2014), the glycan profile could be a reliable biomarker in aging progress and even in the pathogenesis of aging-related diseases.
Another important role that glycosylation plays in IgG is modulation of ADCC: the addition of fucose residues in the core of the glycan (core-fucose) modifies the conformation of the Fc domain in a way to significantly reduce its affinity for FcγRIIIa; therefore core-fucosylated IgGs are less efficient in activating ADCC (Anthony and Ravetch, 2010; Iida et al., 2006). Most of IgGs (over 95%) are core fucosylated, which can prevent them from wrong elicitation of ADCC (Scanlan et al., 2008). Decreased IgG Fc sialylation could reduce its CDC activity through C1q binding, which could lead to increase in activation of the lectin-initiated complement pathway. These changes of the IgG N-glycan may make them possible converting from anti-inflammatory to proinflammatory effects upon antigenic challenge (Zhang et al., 2021). The main proposed roles of IgG N-glycan involved in the pathogenesis of aging progress and aging-related disease are demonstrated in Figure 2.
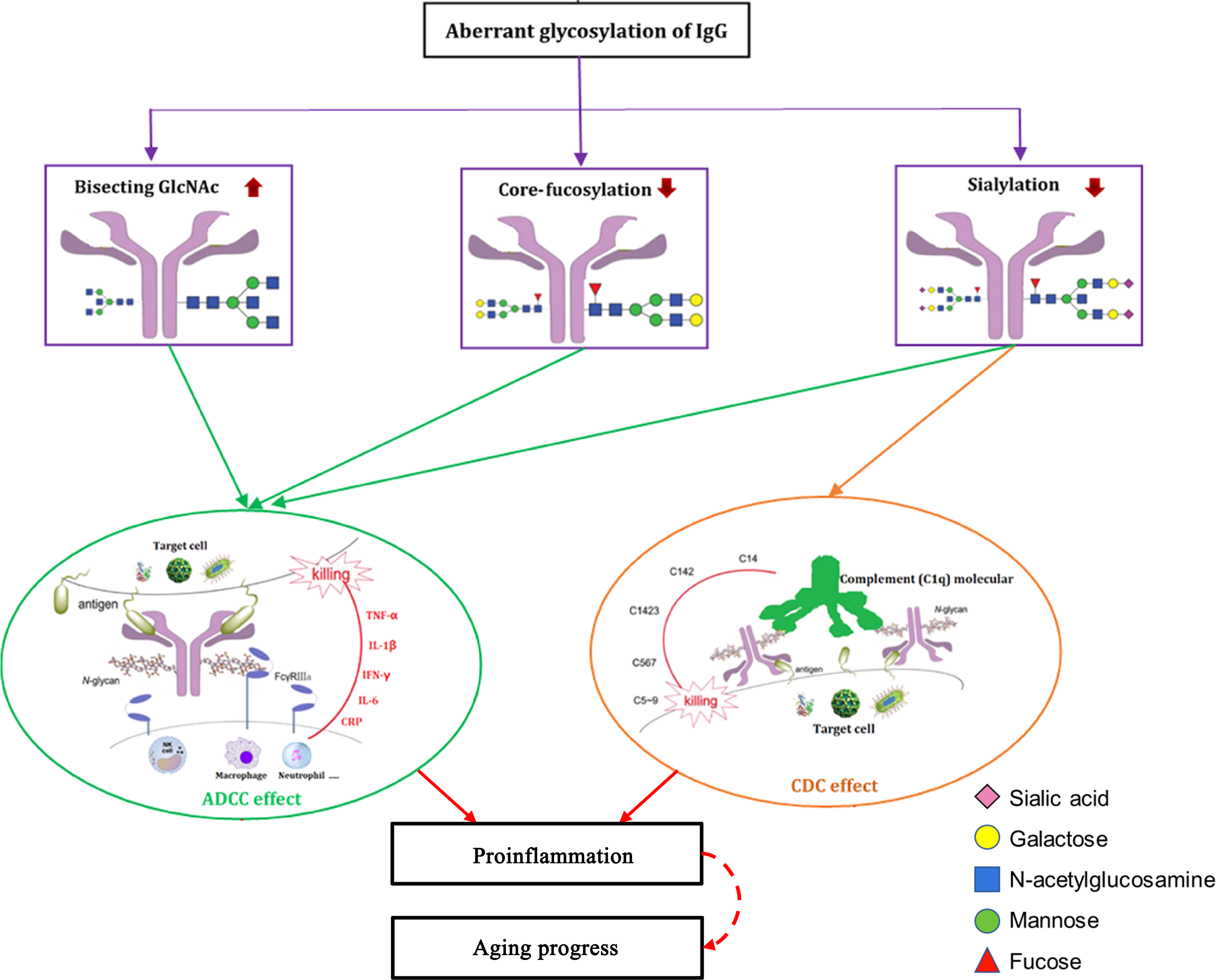
The proposed roles of IgG N-glycans in pathogenesis of aging progress and aging-related diseases. The individual variations in N-glycosylation of IgG contribute to pathogenesis of aging and aging-related diseases, presumably through their effects on pro- and/or anti-inflammatory, ADCC, and CDC pathways. The decreasing sialylation and core fucosylation as well as increasing the bisecting GlcNAc contents within the Fc domain of IgG could activate its ADCC by reducing inhibition to ligate Fcγ-RIIIa on NK cells, macrophage, and neutrophil, which could release proinflammatory factors (such as IL-1β, IL-6, CRP, TNF-α, and IFN-γ). Decreases in IgG Fc sialylation could reduce its CDC activity through C1q binding, which could then lead to increased activation of the lectin-initiated complement pathway. These changes of the IgG N-glycan can translate to a host of effects from anti-inflammatory to proinflammatory, upon antigenic challenge. Figure is reproduced from Zhang et al. (2021) with permission. ADCC, antibody-dependent cell cytotoxicity; CDC, complement-dependent cytotoxicity; CRP, C-reactive protein; FcγR, Fcγ receptor; IFN, interferon; IL, interleukin; NK, natural killer; TNF-α, tumor necrosis factor-α.
Outlook
Aging is one of the most important phenotypes in medicine and ecology. Aging biomarkers warrants more in-depth research. New post-genomic technologies such as glycomics offer prospects such as real-time or longitudinal analyses of aging. Current glycomics evidence shows that aging is closely correlated with IgG Fc monogalactosylation. IgG glycosylation has also been shown to change with both chronological and biological age of an individual. The decrease in levels of digalactosylation and increase in bisecting GlcNAc levels were observed in population-based aging studies. Age-related decline in IgG galactosylation is associated with increases in systemic inflammation in older people, which can be explained by higher levels of proinflammatory cytokines, such as TNF-α and IL-6, found in older individuals. Accordingly, decreased galactosylation also enhances the proinflammatory potential of IgG. The decrease in IgG galactosylation has been observed in numerous inflammatory and autoimmune diseases, such as IBD, RA, and SLE. Population-specific IgG N-glycosylation evidence suggests that persons in developing countries may be exposed to environmental factors that can cause low-grade chronic inflammation and an apparent increase in biological age (Tambuk et al., 2020). Although we still do not fully know the exact mechanisms underlying the age-related changes in whole plasma/serum glycan and IgG glycan profiles, the cellular and humoral immunity pathways identified thus far suggest various enzyme activities involved in glycosylation processes through modulation of the balance between pro- and anti-inflammation, and that this fine balance contributes to the process of both chronological and biological aging.
This expert review underscores the marked potential of glycan monitoring as an approach to better understand and unravel the biomarkers of aging progress, and aging-related diseases. While new digital technologies such as three-dimensional facial-image analysis (Xia et al., 2020) and epigenetic approaches (Saul and Kosinsky, 2021) offer comprehensive studies of temporal and spatial changes in aging-related phenotypes, glycomics brings in yet another systems science technology platform to strengthen the emerging multiomics studies of aging and age-related diseases.
Footnotes
Author Disclosure Statement
The authors declares that no conflicting financial interests exist.
Funding Information
This work was partially supported by Australia–China International Collaborative Grant (NHMRC APP1112767-NSFC 81561128020) and an Edith Cowan University SRF-Seigo Co., Ltd., Grant 2020–2022.