
Abstract
Many cellular functions important for solid tumor initiation and progression are mediated by members of the integrin family, a diverse family of cell attachment receptors. With recent studies emphasizing the role of the tumor microenvironment (TME) in tumor initiation and progression, it is not surprising that considerable attention is being paid to integrins. Several integrin antagonists are under clinical trials, with many demonstrating promising activity in patients with different cancers. A deeper knowledge of the functions of integrins within the TME is still required and might lead to better inhibitors being discovered. Integrin expression is commonly dysregulated in many tumors with integrins playing key roles in signaling as well as promotion of tumor cell invasion and migration. Integrins also play a major role in adhesion of circulating tumor cells to new sites and the resulting formation of secondary tumors. Furthermore, integrins have demonstrated the ability to promoting stem cell-like properties in tumor cells as well as drug resistance. Anti-integrin therapies rely heavily on the doses or concentrations used as these determine whether the drugs act as antagonists or as integrin agonists. This expert review offers the latest synthesis in terms of the current knowledge of integrins functions within the TME and as potential molecular targets for cancer therapeutics innovation.
Introduction
Tumorigenesis involves the transformation of normal cells into cancer cells via mechanisms that include activation of many signaling cascades. Several studies have shown that integrins are directly involved in tumor initiation and progression via influencing cancer cell proliferation, migration, invasion, and survival as well as partaking in cellular signaling (Guo and Giancotti, 2004; Mitra and Schlaepfer, 2006).
Most studies on integrins have focused on their role in cellular migration and invasive behavior. This is mostly due to integrins binding to extracellular matrix (ECM) proteins and aiding in creating traction needed by cells for migration and invasion (see Table 1 for definitions). In addition, several tumor microenvironment (TME) components, including biological molecules such as growth factors and proteases owe their localization to certain regions to integrins (Dzobo et al., 2012, 2014). Thus, integrins can influence cellular proliferation and signaling through “capturing” growth factors and proteases in certain regions of the TME (Assoian and Klein, 2008; Vellon et al., 2005).
Definitions Used in the Article
ECM, extracellular matrix; TME, tumor microenvironment.
In cancer, integrins play a significant role in controlling cellular growth through binding to ECM proteins such as collagens and fibronectin and relay extracellular signals into the cell to affect gene expression (Erdogan et al., 2017; Gopal et al., 2017; Han et al., 2006b). It is important to point out that integrins relay both pro- and antitumorigenic signals, with the balance determining tumor growth and inhibition. Advances in TME biology have revealed that integrins, bound to ECM proteins or not, influence tumor growth via influence on cancer cell stemness as well as chemoresistance (Hamidi et al., 2016; Seguin et al., 2015). Understanding the behavior of integrins in normal tissues and cancer has huge implications for their therapeutic targeting.
Beside integrins found on cancer cells, integrins are also present on cells associated with tumors and play a great role in influencing stromal cell–tumor cell interactions (Attieh et al., 2017; Izumi et al., 2016; Zeltz et al., 2020). These stromal cells range from cancer associated-fibroblasts, -macrophages, -endothelial cells as well as pericytes (Bussard et al., 2016; Dzobo and Dandara, 2020a, 2020b; Franco et al., 2010). Most of these stromal cells are directly linked to processes occurring during tumor development and metastasis such as inflammation, angiogenesis, and desmoplasia (Castells et al., 2012; Senthebane et al., 2017, 2018). Integrins traverse the lipid bilayer and are the link between the extracellular space and the cytoplasm (Bianconi et al., 2016; Hynes, 2002). Integrins relay extracellular cues through receptors for growth factors and cytokines and thus promote cellular signaling. Being present in both cancer cells and stromal cells, integrins influence tumor development as well as metastasis.
New findings show that specific integrins are required by certain growth factors and oncogenes during tumor initiation and growth (Assoian and Klein, 2008; Vellon et al., 2005). This makes it critical to delineate the crosstalk between integrins and growth factors and oncogenes during drug development. To date, several studies both preclinical and clinical trials have been done using integrin inhibitors and these have demonstrated various levels of efficacy at blocking tumor progression (Altei et al., 2020; Zhang et al., 2020). In addition, integrin inhibitors have shown little or no known side effects in some cancer patients (He et al., 2019; Zhang et al., 2019; Zhu et al., 2019). The success demonstrated with integrin inhibitors must be augmented by further research into the role of integrins in different cancers as well as in tumor development and growth.
Literature Search Methodology
Literature search
A search for literature material was conducted on PubMed and MEDLINE databases until February 2021 for published articles with the following keywords: integrins, tumor microenvironment, gene expression, solid tumors, signaling, growth factors, angiogenesis, and metastasis. Language of communication was set to English and full articles were included (Fig. 1).
Selection of studies included in the qualitative synthesis of the review article.
A total of 1421 articles were used and triaged for inclusion/exclusion in the present analyses and review with the aid of the criteria as shown in Figure 1. A final total of 137 articles were included in the present expert review article. The review scope enabled the synthesis of the literature on integrins as key players in cellular processes and signaling and their possible therapeutic targeting using different therapies.
Integrin Biological Properties
Integrins are receptors that traverse the lipid bilayer of the cell membranes and their main function is to relay extracellular cues to cells by binding to the ECM and other immunoglobulin molecules (Fig. 2) (Hynes, 2002, 2004; Hynes et al., 2002; Juliano and Varner, 1993). A minimum of 24 different heterodimer combinations from 18 α-subunits and 8 β-subunits are formed. Each combination of α and β subunits specifically bind to certain ECM proteins and immunoglobulin molecules (Hynes, 2002; Juliano and Varner, 1993). The formation of the integrin dimers occur in the endoplasmic reticulum with posttranslational changes occurring in the Golgi apparatus before the inactive integrins are transferred to the cell surface (De Franceschi et al., 2015).

Integrins can have open and closed configurations that influence adhesions to the ECM. The classic outside-in integrin signaling involves integrins binding to the ECM and activation of several intracellular signaling cascades, including the Ras-MEK-ERK signaling. The translocation of phosphorylated ERK1/2 molecules to the nucleus results in expression of genes involved in cellular proliferation, migration, and survival. Figure was adapted from Dzobo et al. (2012). ECM, extracellular matrix.
Different cells express different integrins and this greatly influence adhesion to different surfaces as well as ability to migrate (Hynes, 2002; Senthebane et al., 2018). The recognition and binding of integrins to their respective ligands may be influenced by the specific sequence on the ligand. For example, integrins αv and α5β1 recognize and bind to ligands with the arginine-glycine-aspartate (RGD) sequence (Pytela et al., 1985). Furthermore, integrin α4β1 recognizes and binds to the REDV and EILDV adhesive sequences on several ECM proteins (Aota and Yamada, 1995; Desgrosellier and Cheresh, 2010). Adhesion to the ECM results in integrins forming focal adhesions which are clusters of integrins and signaling molecules and proteins (Burridge and Chrzanowska-Wodnicka, 1996; Critchley, 2000).
Studies done in two-dimensional and three-dimensional report different compositions of focal adhesions for the same cells (Berrier and Yamada, 2007; Lock et al., 2008). Integrins are able to cluster and therefore activate kinases within focal adhesions such as focal adhesion kinases and Src family kinases (Arias-Salgado et al., 2003; Miyamoto et al., 1996; Xue et al., 2010).
In addition, integrins are able to couple the cytoskeleton and the ECM through recruiting several proteins such as α-actin and tensin (Calderwood et al., 2000; Schwartz, 2010). Together with parvin and PINCH, integrins are able to form large platforms that interact with actin and several signaling cascades (Legate et al., 2006; Sakai et al., 2003). A family of transmembrane proteins known as tetraspanins cross the cell membrane four times and has been shown to form complexes through interaction with several proteins, including integrins (Hemler, 2005; Zöller, 2009). Integrins and several of these proteins involved in focal adhesions and other complexes are under investigation as possible target for therapies in many diseases. In addition, oncogene or growth factor-induced signaling may influence integrin-binding affinity, contributing to disease states and progression (Han et al., 2006a; Lee et al., 2009).
Integrin Contribution to Tumorigenesis
Several pieces of evidence have shown that integrins contribute toward cancer growth and progression (Fig. 3). With many cancers starting in epithelial cells, integrins expressed by these cells play key roles during cancer growth and progression. Integrins increase cellular signaling via interactions with FAK, PI3K-Akt, and the MEK-ERK pathways (Dzobo et al., 2012, 2014). In addition, stromal cells, including caner-associated fibroblasts synthesize increased amounts of ECM proteins and these display enhanced interaction with integrins. Several reports show that integrins are also involved in metastasis and the priming and colonization of new sites for secondary tumors to form (Albelda, 1993; Felding-Habermann et al., 2001; White and Muller, 2007). Furthermore, cancer therapy has been shown to induce integrins expression leading to drug resistance (Aoudjit and Vuori, 2012; Liu et al., 2013; Naci et al., 2012).
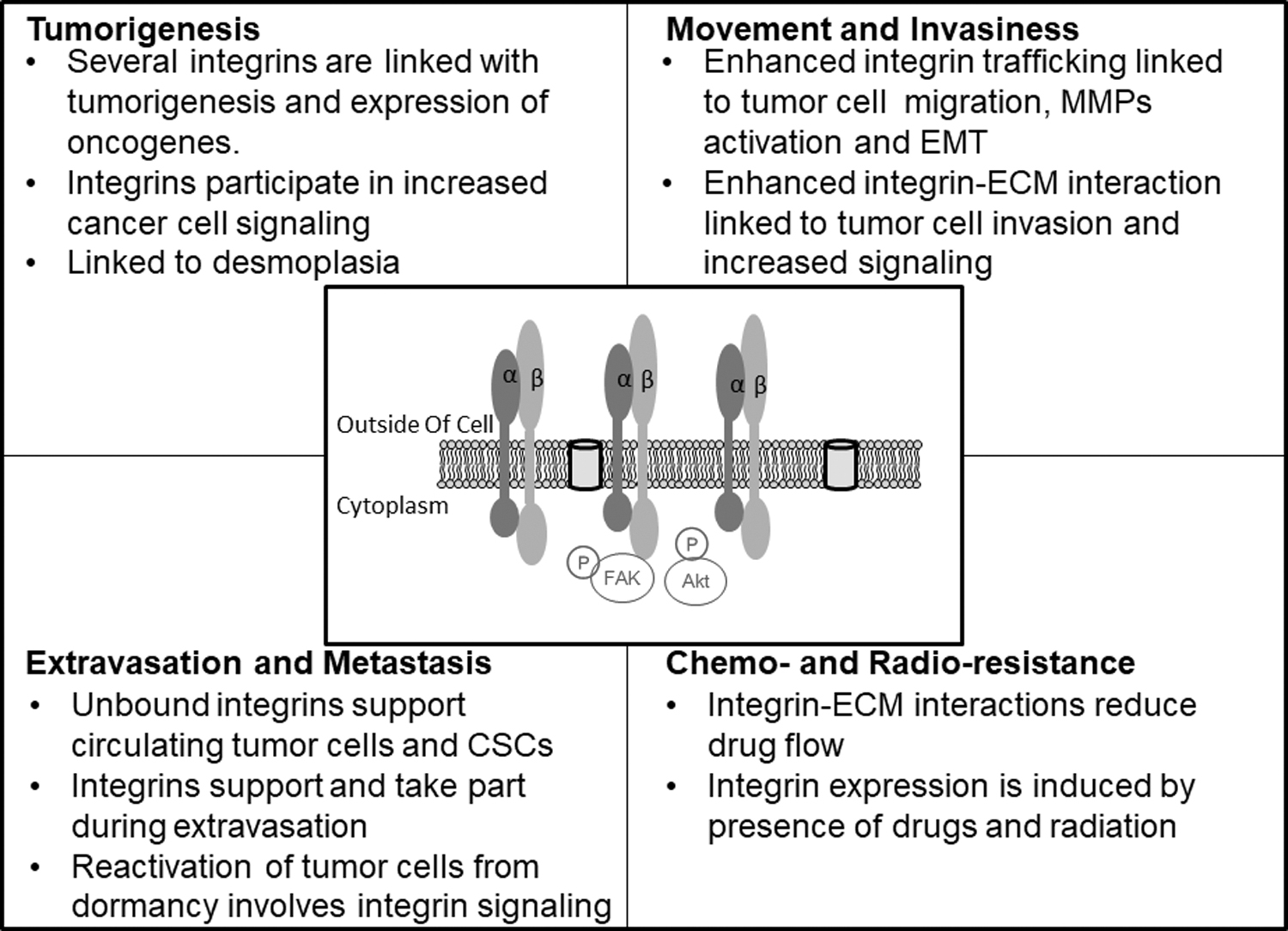
The effect of integrins on different stages of tumor progression. Integrins are involved at every stage of tumor progression starting with tumor initiation to tumor cell migration and invasion, extravasation and metastasis, and finally the development of therapy resistance.
The Significance of Integrins in Tumorigenesis
Examples of integrins expressed by epithelial cells include α2β1, α6β4, and α3β1 and may display dysregulated expression in tumors. While data strongly show these integrins role in cellular attachment to the ECM and the basement membrane, recent data also showed their participation in cellular proliferation, migration, and metastatic behaviors (Aoudjit and Vuori, 2012; Lamb et al., 2011; Petitclerc et al., 1999). Integrins expression has also been shown to change when epithelial cells transform to cancer cells. For example, αvβ6 and αvβ3 integrins are known to be highly upregulated in tumors such as ovarian and colon compared to normal cells (Agrez et al., 1999; Ahmed et al., 2002a, 2002b; Thomas et al., 2001). In addition, these integrins are also associated with increased metastasis and tumor cell proliferation in ovarian cancer (Landen et al., 2008; Slack-Davis et al., 2009).
Several reports also show that some integrins decrease in expression in some tumors. For example, α2β1 integrin expression is decreased in breast tumors (Zutter et al., 1990, 1993, 1995). Decreased integrin expression in cancer cells has been linked with invasion and metastatic behavior (Kaur et al., 2009; Meyer et al., 2004; Thomas et al., 1997). Several studies have linked integrin expression in tumors to patient survival and disease progression, with integrins αvβ3, α6β4, and αvβ6 among the most studied. As new data are generated, the picture become clear on which integrins are important in different tumors. McCabe et al. (2007) demonstrated that αvβ3 integrin is associated with metastatic behavior of prostate cancer cells to the bone matrix.
In addition, the expression of αvβ3 and αvβ6 is linked to decreased cervical cancer patients survival (Gruber et al., 2005; Hazelbag et al., 2007). Integrin α5β1 expression is associated with metastasis and decreased cancer patient survival in melanoma and lung carcinoma (Adachi et al., 2000; Hieken et al., 1999; Nip et al., 1992). The expression of αvβ3 is linked to pancreatic cancer cells metastasizing to the lymph node (Hosotani et al., 2002).
Integrin Promotion of Tumor Cell Survival
Integrins have been shown to act in both a pro- and antitumorigenic manner, depending on signals within the TME. Tumor cell-expressed integrins promote cell survival via involvement in proliferation, invasion, and metastasis among several processes. In the case of invasion, integrins binding to different ECM proteins generate the needed traction for movement and invasive behavior. Formation of focal adhesions and the involvement of matrix metalloproteinases (MMPs) at the leading edge allow cells to invade new territories and tissues (Das et al., 2017). Several reports, however, point to the involvement of some integrins in preventing tumor cell migration. For example, Kren et al. (2007) demonstrated that the absence of integrin β1 increased cancer cell metastatic behavior.
Integrins can promote tumor cell survival through participation in several signaling mechanisms, while integrins have also been shown to play in a role in the process of apoptosis. How integrins behave depends on whether they are free or bound to their ligands. A critical balance is maintained through integrins interacting with ligands and this helps to maintain cell and organs integrity. For example, the binding of integrins to ligands can activate survival pathways such as MEK-ERK, nuclear factor-κβ and PI3K-Akt pathways (Aoudjit and Vuori, 2001; Courter et al., 2005; Dzobo et al., 2012, 2014; Scatena et al., 1998). Interaction of integrins and the ECM upregulates BCL-2 expression in cancer cells (Matter and Ruoslahti, 2001; Uhm et al., 1999).
The interaction between integrins and growth factor receptors also play a role in preventing apoptosis in cells (Alavi et al., 2003; Hood et al., 2003). Deletion of integrins genes or inhibition of integrin-mediated signaling has been shown to activate a compensatory upregulation of vascular endothelial growth factor receptor-2 (VEGFR2) expression, complicating the targeting of integrins in pathological conditions such as cancer (Reynolds et al., 2002, 2004; Weis et al., 2007). Several data show that integrin function in tumors may be context dependent, with data showing that integrins such as αvβ3 is protumorigenic but can inhibit tumor growth in cancers such as melanoma and glioblastoma (Danen et al., 1996; Kanamori et al., 2004; Petitclerc et al., 1999; Varner et al., 1995).
A subpopulation of cells known as cancer stem cells (CSCs) that is thought to give rise to all tumor cells present in tumors (Dzobo, 2020; Dzobo and Dandara, 2020a, 2020b; Dzobo et al., 2016). Several pieces of evidence point to integrins playing a part in tumorigenesis and as markers of CSCs (Hamidi et al., 2016; Nieberler et al., 2017). For example, Asselin-Labat et al. (2007) demonstrated that stem cell-like cells within the mammary ductal epithelium express αvβ3 integrin and it can be used a marker of these cells. Vaillant et al. (2008) revealed that β3 integrin and WNT1 were both markers of mammary CSCs. Luo et al. (2009) demonstrated that removal of integrin-associated focal adhesion kinase signaling reduced the number of CSCs in mammary tumors in mice. Samanna et al. (2006) showed that αvβ3 expression was linked to tumorigenicity and MMP-2 activity in melanoma cells. Specifically, the authors showed that integrins influence the expression of CSCs markers such as CD44 (Samanna et al., 2006).
Integrins Contribution Toward Tumor–Host Cell Interactions
Integrins are also involved in the response of normal cells to the presence of tumor cells. Reports indicate that most cells, including fibroblasts, mesenchymal stem cells, pericytes, immune cells, and endothelial cells, utilize integrins in cellular processes such as migration, angiogenesis, and desmoplasia. The targeting of integrins on stromal cells may aid in limiting the “support” given to tumor cells by normal cells.
Integrins are involved in every stage of tumorigenesis from initiation to metastasis (Avraamides et al., 2008). Several pieces of evidence points to “leaky” blood vessels in tumors which causes impaired blood flow and drug delivery (McDonald and Baluk, 2002). In addition, leaky blood vessels are also known to promote fibrosis in tumors (Brown et al., 1989). Brooks et al. (1994a) showed that integrin αvβ3 was upregulated in chick chorioallantoic membrane and associated to angiogenesis.
It has been speculated that integrins may enable the interaction between ECM proteins and angiogenic-associated cells such as endothelial cells within the TME (Eliceiri and Cheresh, 1999; Stupack and Cheresh, 2002). Remodeling of the ECM has also been linked to increased integrin binding to degraded ECM proteins such as collagen (Davis, 1992). Several reports have demonstrated that integrins are important for cellular processes such as migration and angiogenesis (Avraamides et al., 2008). Various compounds or small molecules in addition to genetic manipulation have confirmed the involvement of integrins, including α2β1, α6β1, and α6β4 (Avraamides et al., 2008; Dzobo et al., 2012, 2014).
Together with growth factor receptors, integrins can relay signals from growth factors to influence angiogenesis in tumors (Brooks et al., 1994a, 1994b; Friedlander et al., 1995). In elaborate experiments, Friedlander et al. (1995) demonstrated that protein factors, basic fibroblast growth factor, and tumor necrosis factor-α require the integrin αvβ3 to induce angiogenesis in corneal. In addition, the same authors also showed that vascular endothelial growth factor and transforming growth factor (TGF)-α induced angiogenesis through interaction with integrin αvβ5 (Friedlander et al., 1995). Most importantly, integrins can bring together signals from both the ECM and several growth factors to influence cellular processes.
Several cells have been shown to play a role in angiogenesis and these include vascular smooth muscle cells and pericytes. Blood vessel maturation, for example, requires pericytes and smooth muscle cells (Goede et al., 1998; Klein et al., 2011; Korff et al., 2001; Raza et al., 2010). Most blood vessels found within the TME are abnormal and leaky, leading to less oxygen reaching all cells.
For example, interaction between pericytes and endothelial cells is via integrin α4β1 and vascular cell adhesion molecule-1 (Garmy-Susini et al., 2005). This may lead to blood vessel formation and stability (Garmy-Susini et al., 2005). The branching of blood vessels within tumors also requires pericytes involvement through integrin binding (Fukumura and Jain, 2007b). Thus, a combination of therapeutic targeting tumor cells as well as therapeutics targeting angiogenesis and its associated cells to bring about blood vessel normalization may provide a synergistic and durable effect in cancer treatment (Fukumura and Jain, 2007a; Jain et al., 2007a; Tong et al., 2004). Indeed, it has been suggested that treatment with antiangiogenic therapeutics first before cytotoxic drugs may be better at treating cancer (Tong et al., 2004). Thus today, vascular normalization is part of cancer therapies in many settings and clinics.
One of the observed hallmarks of desmoplasia in tumors is the synthesis and deposition of large amount of ECM proteins such as collagens. Increased collagen within the TME means increased integrin signaling allowing promotion of cancer cell survival and resistance to therapy (Conti et al., 2008). Integrins on both cancer cells and stromal cells contribute toward cancer cell growth via activation of signaling that promote cancer cell growth. For example, Zhu et al. (2007) demonstrated that in nonsmall cell lung carcinoma, several integrins, including α11, are necessary for cancer cell growth. Integrin α11 was instrumental in inducing insulin-like growth factor 2 release in human nonsmall cell lung cancer cells (Zhu et al., 2007). Integrins inhibitors must therefore target both cancer cells and stromal cells for effective and durable cancer treatment.
Several pieces of evidence have shown that bone marrow-derived cells from the bone marrow are found within solid tumors (Adams et al., 2014; Ghossein et al., 1999; Senthebane et al., 2017). Once in the TME, these bone marrow-derived cells can exhibit contrasting effects on cancer cells with some cells having antitumorigenic effects, while others have tumor-promoting effects.
For example, infiltrating macrophages expressing integrin αvβ3 are able to have antitumorigenic effects (Taverna et al., 2004). In addition, bone marrow-derived cells with no functional integrin β3 are not found within tumor sites and cannot therefore promote angiogenesis (Feng et al., 2008). Jin et al. (2006a) demonstrated that cells attracted to tumors, including endothelial cells, require integrin α4β1. Integrin α4β1 is needed for the binding of bone marrow-derived cells to the endothelium within TME (Jin et al., 2006b). In terms of targeting integrins needed for the homing of bone marrow-derived cells to tumors, a balance must be maintained between antitumorigenic and protumorigenic effects.
Tumor metastasis has been associated with the interaction between cancer cells and platelets. For example, Mammadova-Bach et al. (2016) demonstrated that integrin α6β1 on platelets promotes metastasis via binding to lung cancer cell-derived ADAM9.
In addition, Gay and Felding-Habermann (2011) showed that platelets can alter tumor cell characteristics and promote metastasis. Furthermore, fibrinogen can behave as a connection between platelets and cancer cells via attaching to integrins on both cells and this allows cancer cells to be associated with blood vessels (Bakewell et al., 2003; Felding-Habermann et al., 1996). Metastasis to the bone marrow and the lungs is also made possible via interaction between cancer cells and platelets (Felding-Habermann et al., 1996; Jain et al., 2007b). Trikha et al. (2002) demonstrated that the combined inhibition of integrins on both cancer cells and platelets enhances the antitumorigenic of the inhibitors compared to inhibition of integrins on tumor cells.
Integrin Crosstalk with Other Receptors
Cooperation between integrins and oncogenes in promoting tumor growth has been noted. Guo et al. (2006) demonstrated that integrin α6β4 cooperates with ERBB2 and enhances tumor initiation and invasion in breast cancer. There is also an association between integrin β1 and polyoma middle T oncoproteins in driving breast cancer (White et al., 2004). Some lung tumors require the involvement of integrin α1 for the initiation and growth of the tumors (Macias-Perez et al., 2008). The Src oncogene activity has been shown to be enhanced via its interaction with integrin α4β3 (Huveneers et al., 2007). These several pieces of evidence demonstrate that integrin signaling is involved in enhancing the protumorigenic effect of many oncogenes.
Tumor development and growth studies have shown that integrins play crucial roles through interactions with growth factors as well as cytokines. The involvement of integrins in adhesion, migration, and invasion of tumor cells is well documented, however, integrins have been shown to play more roles, including enhancing angiogenesis and signaling. Interaction between integrins and growth factors and cytokines is not always protumorigenic, with several pieces of data showing that integrins interactions can be antitumorigenic (Shimizu et al., 2003). The interaction of integrins and growth factors is likely to influence signaling pathways involving kinases to enhance tumor cell survival and invasiveness (Assoian and Schwartz, 2001; Tai et al., 2003). Growth factors can also influence integrin internalization and activity and vice versa (De et al., 2005; Ignotz and Massagué, 1987; Ning et al., 2005, 2007; Wang et al., 1999, 2009).
The cooperation between integrins and growth factors and their receptors has been noted for its role in tumor development, growth, and eventual metastasis in several cancers. Involvement of growth factors, including the epidermal growth factor (EGF) receptor family can be through increased expression or activation leading to oncogenic signaling in tumor cells.
For example, enhanced expression of ERBB2, also known as v-erb-b2 avian erythroblastic leukemia viral oncogene homolog 2, and its receptors is observed in several cancers, including breast and nasopharyngeal carcinoma (Joensuu et al., 2003; Roychowdhury et al., 1996; Shen et al., 2014; Tsutsui et al., 2002). Cooperation between integrins and the EGF receptor family has been reported in breast cancer. Guo et al. (2006) demonstrated that integrin α6β4 cooperates with ErbB2 signaling in breast cancer to promote tumor development and growth. This cooperation also promoted invasion of tumor cells (Guo et al., 2006). Furthermore, the authors showed that this cooperation causes the activation of the signal transducer and activator of transcription 3 signaling, a signaling pathway known to cause increased cellular proliferation (Guo et al., 2006).
The targeting of both α6β4 integrin and the ERBB2 may produce a synergistic therapeutic effect (Guo et al., 2006). The interaction between integrins, growth factors, and ECM proteins such as collagen and glycoproteins such as vitronectin can promote cancer cell migration and metastasis (Brooks et al., 1997; Klemke et al., 1994; Ricono et al., 2009). Integrin αvβ5, for example, interacts with EGF and vitronectin to promote tumor cell migration (Brooks et al., 1997; Klemke et al., 1994; Ricono et al., 2009). Pouliot et al. (2001) demonstrated that EGF cooperates with α3β1 and the ECM protein laminin, for example, to promote migration of human colon carcinoma cells. Another important study demonstrated that integrins α1β1 and α2β1 interact with EGF to promote the invasive behavior of hepatocarcinoma cells in a fibrotic microenvironment (Yang et al., 2003).
The cooperation between integrins and the hepatocyte growth factor (HGF) receptor, mesenchymal epithelial transition factor (c-Met), has been implicated in tumor initiation and invasion. Bertotti et al. (2005) demonstrated that integrin β4 cooperates with HGF receptor, MET, to promote fibroblasts transformation and the eventual involvement of such transformed fibroblasts in tumorigenesis. In another study, the same authors also showed that through induction of integrin β4 phosphorylation, MET signaling upregulates anchorage-independent growth (Bertotti et al., 2006). Reports show that integrin αvβ5 can regulate the expression of genes induced by HGF and involved in cell invasion (Crouch et al., 2004). Overall, HGF-MET signaling cascade involvement in tumorigenesis is dependent on cooperation with integrins.
Another growth factor shown to be involved in promoting tumorigenesis is TGF-β. For example, TGF-β has been shown to play a key in epithelial-mesenchymal transition (EMT) leading to increased tumor cell movement and invasiveness (Oft et al., 1996; Vo et al., 2013; Welch et al., 1990). Integrins such as αvβ6 and αvβ8 play a key role in the activation of TGF-β signaling through binding to the RGD sequence of the latency-associated peptide of TGF-β1 (Annes et al., 2002; Lu et al., 2002; Ludbrook et al., 2003; Munger et al., 1998).
Marsh et al. (2008) demonstrated that enhanced integrin αvβ6 expression together with increased TGF-β activation leads to aggressive disease in basal cell carcinoma. Immunohistochemical studies and transcriptional activation experiments confirmed that αvβ6 activates the TGF-β signaling cascade in vivo and EMT leading to tumor growth (Bates et al., 2005; Van Aarsen et al., 2008). TGF-β signaling together with integrin αvβ3 activates EMT process in epithelial cells (Galliher and Schiemann, 2006). Our study demonstrated that both tumor cells and stromal cells secrete TGF-β, and it is plausible that the crosstalk between TGF-β and integrins play a key role in tumor growth and progression (Senthebane et al., 2017).
Integrins cooperation with growth factor occurs between tumor cells and stromal cells as well. For example, endothelial cell migration and proliferation require integrin and growth factor interactions during angiogenesis. Several reports have linked specific integrins to specific growth factors during angiogenesis (Brooks et al., 1994a, 1994b; Lehmann et al., 1994; Varner, 1997). Fibroblast growth factor receptor is known to interact with integrin αvβ3 leading to the phosphorylation of serine residues in Raf (Bao and Strömblad, 2004; Hood et al., 2003). The resulting complex known as MAP3K5 complex causes inhibition of the apoptotic pathway, thus protects endothelial cells from apoptosis (Mahabeleshwar et al., 2007).
In addition, VEGFR2 has been shown to interact with integrin αvβ3 and αvβ5 to inhibit apoptosis by regulating the Ras-ERK pathway (Hood et al., 2003). VEGFR2 also interacts with integrin αv in endothelial cells to increase angiogenesis (Byzova et al., 2000; Mahabeleshwar et al., 2007; Reynolds et al., 2002). Increased tumor cell proliferation is observed when activated αvβ3 causes secretion of VEGF (De et al., 2005). Nikolopoulos et al. (2004) demonstrated that integrin β4 cooperates with fibroblasts growth factor to induce angiogenesis and increased tumor growth. CXCR4 is expressed by both tumor and stromal cells and its increased expression has been associated with increased expression of several integrins (Cardones et al., 2003; Engl et al., 2006; Sun et al., 2007).
Targeting Integrins in Cancer Therapy
Integrins are appealing therapeutic targets for cancer therapy due to their increased expression in several cancers and cooperation with growth factors during tumor development and metastasis. Furthermore, integrins have been shown to play a key role in the development of drug resistance. Our work and that of others have specifically shown the involvement of integrins and their ligands in cancer cell drug resistance (Hamidi et al., 2016; Seguin et al., 2014; Senthebane et al., 2017, 2018; Sethi et al., 1999). Several in vitro and animal studies have shown that inhibition of integrin function through the use of antagonists is effective at blocking tumor development and metastasis. Integrin function blockage can affect both tumor cells and stromal cells (Avraamides et al., 2008; Curley et al., 1999; Scarborough et al., 1993; Shimaoka and Springer, 2003). For example, cilengitide is currently under Phase 3 clinical trial for the treatment of glioblastoma.
Data from clinicaltrials.gov show that cilengitide is effective at treating patients with prostate and glioblastoma. A monoclonal antibody, LM609 or its humanized version etaracizumab, demonstrated antiangiogenic activity in many studies, including breast cancer studies (Brooks et al., 1995), but a clinical trial study was terminated as reported on the clinicaltrials.gov website.
Mulgrew et al. (2006) demonstrated that blocking αvβ3 with the monoclonal antibody Abegrin can result in inhibition of tumor growth. Integrin αvβ3 is one of the major integrins expressed by osteoclasts, and its inhibition through the use of monoclonal antibody Vitaxin® blocks bone metastasis (Gramoun et al., 2007). Vitaxin also demonstrated efficacy in patients with renal cancer and other solid tumors (Delbaldo et al., 2008; McNeel et al., 2005). Trikha et al. (2004) and Chen et al. (2008) demonstrated that the monoclonal antibody CNTO95 has antiangiogenic effects and can block cell signaling and invasion in breast cancer cells, respectively. The amount of integrin antagonists used in studies is very important as some studies demonstrated that certain concentrations of doses of antagonists can act as an integrin agonist (Aznavoorian et al., 1990; Legler et al., 2001; Odekon et al., 1991).
Most antiangiogenic drugs increase tumor perfusion, which is linked to better drug delivery in tumors (Ebos et al., 2009; Pàez-Ribes et al., 2009). Thus, several studies have demonstrated that antiangiogenic therapy targeting integrins works well when combined with chemotherapy. In addition, integrin targeting affects not only tumor cells but also several stromal cells within the TME. Integrin β1 via interaction with several signaling cascades, including the Akt and the FAK pathways, has been shown to play key roles in development of lung cancer and head and neck drug resistance (Eke et al., 2015; Kanda et al., 2013). Hirata et al. (2015) demonstrated that enhanced integrin β1 signaling via interaction with FAK was associated with resistance to the highly selective inhibitor of B-Raf, PLX4720.
Vascularized tumors, including glioblastomas, demonstrate increased expression of integrins such as integrin αvβ3 and β8 (Giovanna and Kaye, 2007; Gladson, 1996; Tchaicha et al., 2011). In most cases, these tumors are aggressive and exhibit low patient survival making the development of integrin antagonists necessary to increase patient survival (Giatromanolaki et al., 2004; MacDonald et al., 2001; Yamada et al., 2006).
It has been shown that TME is a crucial determinant of therapy effectiveness, thus targeting angiogenesis and stromal integrins is a viable option (Dzobo, 2020; Dzobo and Dandara, 2020a, 2020b; Giatromanolaki et al., 2004; Giovanna and Kaye, 2007; Senthebane et al., 2017, 2018). For example, esophageal carcinoma abundantly express ECM proteins, some of which are ligands for several integrins and thus can influence cancer cell survival and migration (Senthebane et al., 2018). Thus, it is possible to target integrins such as α2β1 as the ECM ligands, including fibronectin and collagen, are abundantly expressed in esophageal cancer (Dzobo et al., 2012, 2014; Senthebane et al., 2017, 2018). Several clinical trials have been performed or under way to determine the effectiveness of integrin targeting in different cancers (MacDonald et al., 2008; Nabors et al., 2007; Reardon et al., 2008).
Park et al. (2006) demonstrated that an integrin β1 inhibitory, AIIB2, was able to reduce breast cancer survival through activation of apoptosis. In yet another study, Bhaskar et al. (2008) showed that chimeric integrin α5β1 antibody, Volociximab, displayed antiangiogenic activity and inhibited tumor growth. Khalili et al. (2006) also showed that peptide inhibitor of integrin α5β1, ATN-161, was able to inhibit cancer growth as well as metastasis. The same peptide inhibitor, ATN-161, together with commonly used drug fluorouracil, was able to inhibit colon metastasis to the liver (Stoeltzing et al., 2003). A small molecule integrin αvβ3 inhibitor S247 was able to inhibit breast cancer cell metastasis to the bone (Harms et al., 2004). The same effect was demonstrated by another integrin αvβ3 inhibitor PSK1404 (Zhao et al., 2007). S137 and S247 are RGD peptide mimetics and have been shown to inhibit metastasis (Shannon et al., 2004). 6.3G9 is an integrin αvβ6 antibody that has been shown to have anticancer activity in pharyngeal carcinoma cells (Van Aarsen et al., 2008).
One major challenge associated with targeting integrins and its associated cellular processes such as angiogenesis is the lack of validated biomarkers. Several markers such as serum levels of fibroblast growth factor and vascular endothelial growth factor have been suggested to indicate the efficacy of anti-integrin therapy. However, few or no reports have shown the efficacy of such biomarkers in the clinical setting. Recent works and reports reveal new data on the involvement of integrins in several aspects of tumor growth from migration, invasion, and drug resistance (Baltes et al., 2020; Ramovs et al., 2019; Turaga et al., 2021; Yousefi et al., 2021). Current investigations involving coupling integrin antagonists and radionuclides aim to identify newly formed blood vessels in tumors (Cai et al., 2006). In addition, newly formed vessels can also be identified via the use of ultrasound with microbubbles as shown for αv-integrins by Leong-Poi et al. (2003). Angiogenesis in tumors can also be detected via the use of 18F-labeled PEGylated RGD peptide (Chen et al., 2004).
The above noted examples and others demonstrate the potential use of labeled integrins antagonists as both treatment options and as diagnostic tools in cancer therapy. Furthermore, therapeutics targeting integrins can also be used to deliver drugs and proapoptotic peptides to tumor cells. For example, Hood et al. (2003) targeted integrin αvβ3 with a nanoparticle while delivering mutant RAF1 gene to blood vessels within the tumor, leading to tumor shrinkage. Nanoparticles can also be coupled to drugs such as doxorubicin and be able to target integrins and tumor vasculature (Murphy et al., 2008).
Various reports have shown that there is potential therapeutic value in targeting integrins in cancer, with promising data coming from trials of many compounds and inhibitors in different cancers. Based on data from clinicaltrials.gov, several integrin inhibitors have either been completed with minimal benefit to patients or terminated due to lack of efficacy. For example, Abituzumab, a monoclonal antibody against αν integrin intended to prevent integrins and ECM interactions, failed in clinical trials. Part of the reason for this is the ability of tumor cells to signal via integrin-independent mechanisms. Finally, nanoparticles have been used to target integrins in cancer. Nanoparticles can be modified with ligands for integrins to be used to treat cancers. Various nanoparticles can be attached to peptides that target integrins as well as being loaded with nucleotides. Cells displaying high levels of integrins such as cancer cells can take up these nanoparticles.
Conclusions
While most cells within the human body express integrins, those expressed by tumor cells and stromal cells within the TME are involved in several processes along the tumorigenic path. Importantly, integrins have been shown to partake in promoting tumor cell proliferation, survival, migration, as well as metastasis. Recent studies have demonstrated the role played by both bound and unbound integrins with both integrins playing a role in promoting tumor cell survival and metastasis. These newly discovered properties of integrins are important in drug discovery and integrin targeting. Integrins also cooperates with growth factors in promoting tumorigenesis and formation of secondary tumors. Furthermore, integrin investigations must be coupled to ECM studies as both play a role in tumor cell response to therapy and development of therapy resistance.
Footnotes
Author Disclosure Statement
The author declares there are no conflicting financial interests.
Funding Information
No funding was received for this article.