
Abstract
Nematode infections are common in both humans and livestock, with major adverse planetary health and economic impacts. Wuchereria bancrofti is a parasitic nematode that causes lymphatic filariasis, a neglected tropical disease that can lead to severe disability and deformity worldwide. For the long-term survival of the bancroftian parasites in the host, a complex immune invasion strategy is involved through immunomodulation. Therefore, immunomodulation can serve as a site of research and innovation for molecular targets. Macrophage migration inhibitory factor (MIF) is a pleiotropic cytokine crucial to the host antimicrobial alarm system and stress response. Interestingly, the nematode parasite W. bancrofti also produces two homologs of MIF (Wba-MIF1 and 2). Using a mass spectrometry-based phosphoproteomics approach, we report new findings on the immunomodulatory effect and signaling mechanism of Wba-MIF2 in macrophage cells. Accordingly, we observed 1201 phosphorylated sites on 467 proteins. Out of the 1201 phosphorylated sites, 1075, 117, and 9 were found on serine (S), threonine (T), and tyrosine (Y) residues, respectively. Our bioinformatics analysis led to identification of major pathways, including spliceosomes, T cell receptor signaling pathway, Th17 differentiation pathway, interleukin-17 signaling pathway, and insulin signaling pathway upon Wba-MIF2 treatment. Wba-MIF2 treatment also enriched CDK4, CDK1, and DNAPK kinases. The comparison of the signaling pathway of Wba-MIF2 with that of human-MIF suggests both share similar signaling pathways. These findings collectively offer new insights into the role and mechanism of Wba-MIF2 as an immunomodulator and inform future diagnostics and drug discovery research for W. bancrofti.
Introduction
Nematode infections occur in huge numbers of humans and livestock, adversely impacting planetary health and the economy. Lymphatic filariasis caused by three nematode parasites, Wuchereria bancrofti, Brugia malayi, and Brugia pahangi, is a major cause of disability throughout the tropical world, significantly impacting the economically weaker sections of the societies. It is a mosquito-borne neglected tropical disease endemic in 72 countries (WHO, 2020). As per the WHO reports, in 2021, in 44 countries, 882.5 million people were living in areas that require at least preventive chemotherapy to stop the transmission of the infection (WHO, 2020).
Millions of people are affected by lymphatic filariasis, which manifests as lymphedema (elephantiasis) and/or hydrocele and results in social stigmatization and reduced productivity (Lenk et al., 2016; Zeldenryk et al., 2011). The Global Program to Eliminate Lymphatic Filariasis (GPELF) was established in 2000 to work toward eliminating lymphatic filariasis worldwide as a public health problem (WHO, 2010).
Lymphedema is an immune-mediated permanent disability, and there is a felt need to identify intervention targets to avert the disability. A characteristic feature of filariasis is that the parasites live for a long time inside their host, facilitated by a complex immune invasion strategy coupled with immunomodulation, which leads to clinical manifestations (Figueiredo et al., 2010). This is mediated by the action of their excretory/secretory molecules, which include proteases and their inhibitors, antioxidant proteins, and counterparts to mammalian cytokines (Harnett, 2014). The immunomodulatory mechanism is driven by the secretion of anti-inflammatory molecules, immune polarization, and induction of the alternative activation of the macrophage, among others.
W. bancrofti lives in the human host for up to 15 years and is reported to excrete/secrete a rapporteur of immunomodulatory molecules (Amdare et al., 2017; Mukherjee et al., 2019). In Western countries, there are alarming increases in autoimmune and allergic diseases (Thorburn et al., 2014; Versini et al., 2015). However, incidences of such diseases are relatively less in the countries endemic to parasitic helminths. Evidence from human studies also shows that the reactivity of the skin allergens and related symptoms were found to be significantly less in the nematode-infected versus noninfected community (Cooper, 2004; Elliott et al., 2000; Falcone and Pritchard, 2005; Maizels, 2005; van Riet et al., 2007).
Apart from the human studies, animal studies also show ample findings of the inverse relationship between nematodes and autoimmune diseases such as diabetes (Saunders et al., 2007) and colitis (Elliott et al., 2000; Ramani et al., 2020). Thus, overall, idea is that helminths might protect against human allergic and autoimmune inflammatory disease through its anti-inflammatory and immunomodulatory armor strategies of the excretory-secretory molecules (Kaushal et al., 1982). One of these includes a human host cytokine homolog, human-migration inhibitory factor (h-MIF) (Moreno and Geary, 2008).
The h-MIF is a pleiotropic cytokine that acts as an integral component of the host antimicrobial alarm system and stress response that promotes the pro-inflammatory functions of immune cells, and it also possesses two enzymatic functions: tautomerase and oxidoreductase activities (Trivedi-Parmar and Jorgensen, 2018). It is released by several cells of the innate and adaptive immune system, as well as nonimmune cells and plays a significant role in immunomodulation and various disease conditions, such as ulcerative colitis, type 1 diabetes (T1D), and during infection (Karabowicz et al., 2022). Therefore, immunomodulation can serve as a site of research and innovation for molecular targets.
Earlier, we identified two homologs of h-MIF from W. bancrofti, that is, Wba-MIF1 and Wba-MIF2, and expression cloned as recombinant molecules. Upon functional characterization, they were also found to possess two enzymatic activities similar to h-MIF (Chauhan et al., 2015; Sharma et al., 2012). In addition, a separate study has noted that filarial worms can offer protection against the development of T1D in non-obese diabetic (NDD) mice through a mechanism involving a shift toward Th2 immune responses (Hubner et al., 2009; Stosic-Grujicic et al., 2008). Our recent study has shown that Wba-MIF2 immunomodulates the Streptozotocin-induced T1D in mouse model (Personal communication; Singh et al., 2024). However, the signaling mechanisms underlying the immunomodulation by Wba-MIF2 need to be understood to substantiate its role.
In this study, we aimed to unravel these mechanisms through proteomics and phosphoproteomic approaches under in vitro conditions using mice macrophage cell line (RAW264.7) as a study model. This type of molecular mechanistic study may help us to establish an integrative approach for understanding the immune invasion strategy utilized by nematode parasites. Also, unraveling the underlying immunomodulatory mechanism(s) of Wba-MIF2 may provide leads for diagnostics and therapeutic innovations for lymphatic filariasis.
Materials and Methods
Wba-MIF2 expression, purification, and endotoxins removal
Wba-MIF2 was purified as reported previously with some modification (Figueiredo et al., 2010). The detailed methodology is provided in Supplementary Information.
Enzymatic activity
Oxidoreductase activity
Oxidoreductase activity of the recombinant Wba-MIF2 was checked by insulin reduction assay, as per the previously reported method (Figueiredo et al., 2010; Harnett, 2014).
Tautomerase activity
Tautomerase activity was checked as reported previously by insulin reduction assay (Chauhan et al., 2015; Paily et al., 2009).
Phosphoproteomics analysis to study the signaling mechanism of Wba-MIF2
Cell culture conditions
RAW 264.7 cells (TIB-71™) were procured from the American Type Culture Collection (ATCC, Manassas, VA). These cells were cultured and maintained in Dulbecco's modified Eagle's medium, containing 10% fetal bovine serum and 1% antibiotic and antimycotic mixture (Thermo Fisher Scientific). The cells were incubated at 37°C in an incubator with humidity and 5% CO2. RAW 264.7 cells were treated with 100 ng/mL of h-MIF or parasitic Wba-MIF2 for different time points.
Protein extraction from cell lines treated with h-MIF and parasitic MIF
The proteins from the treated cells were extracted as described earlier (Karthikkeyan et al., 2020; Najar et al., 2021a). The detailed method is provided in Supplementary Information.
Sample preparation for proteomics
Proteins from each treatment underwent reduction, alkylation, and trypsin digestion as described previously and detailed in the Supplementary Information (Rex et al., 2022b).
Basic pH reverse-phase liquid chromatography-based fractionation
Fractionation was conducted in accordance with the previously described protocol (Najar et al., 2021b; Verma et al., 2017). The tandem mass tagging (TMT) 10plex-labeled dried samples were reconstituted in solvent A for basic pH reverse-phase liquid chromatography (10 mM TEABC) and subjected to chromatographic separation using a Hitachi LaChrom Elite HPLC system. Peptides were separated using a Waters XBridge C18 column (Waters Corporation, Milford, MA; 130Å, 5 μm, 250 × 4.6 mm) attached to the HPLC system. Peptide desalting was accomplished through the C18 Stage Tip method, as previously outlined (Rex et al., 2022a).
Phosphopeptides enrichment using titanium dioxide
Phosphopeptide enrichment was conducted utilizing titanium dioxide (TiO2) beads, following a protocol described in a previously published study (Kalal et al., 2021; Selvan et al., 2014).
Liquid chromatography-tandem mass spectrometry (LC-MS/MS) analysis
Liquid chromatography-tandem mass spectrometry (LC-MS/MS) analysis was conducted employing an Orbitrap Fusion Tribrid mass spectrometer (Thermo Fisher Scientific, Bremen, Germany) in conjunction with an Easy-nLC 1200 nanoflow UHPLC system (Thermo Scientific, Odense, Denmark), as previously outlined (Shaikh et al., 2021). For peptide separation, an EASY-Spray C18 column (75 μm × 15 cm, 100Å) was utilized, maintaining a constant temperature of 40°C. Each sample underwent a 140-min acquisition. Further details are provided in the Supplementary Information.
Database search for protein and peptide identification
Proteome discoverer (PD) version 2.2 (Thermo Fisher Scientific, Bremen, Germany) was used to process the MS raw files. The raw data were searched against the mouse reference proteome database, which also included known contaminant protein sequences (Mouse RefSeq 109 protein database containing 92,449 protein entries with 116 common contaminants). This was achieved through the employment of the MASCOT and SequestHT search algorithms.
The search parameters were set as previously described (Verma et al., 2017): a minimum peptide length of seven amino acids, allowance for up to two missed cleavages with trypsin as the proteolytic enzyme, and maintained precursor and fragment ion mass tolerances of 10 ppm and 0.05 Da, respectively. Static modification was used, which involved the addition of TMT to the peptide N-terminus and lysine residues and carbamidomethylation of cysteine. Furthermore, oxidation of methionine and protein N-terminus acetylation were treated as dynamic modifications.
A false discovery rate was upheld at 1% at both the peptide and peptide-spectrum match (PSM) levels using the percolator node within PD to maintain a high confidence level. In the case of phosphoproteomic raw data sets, a search with the inclusion of a phospho moiety (+79 Da) as a variable post-translational modification on serine (S), threonine (T), and tyrosine (Y) residues was conducted. In this instance, the ptmRS algorithm was applied to determine the probability of phosphorylated site localization. Phosphopeptides with a site localization probability of 75% or greater were considered genuine modified sites and subsequently used for protein identification.
Data analysis
The mass spectrometry data are acquired in technical triplicates. The raw protein and phosphopeptide (phosphoproteomic analysis) abundance values were used for relative quantification analysis. Proteins/phosphopeptides with no abundance values across all channels were excluded from further analysis. Missing values were imputed by zero, and the batch correction was performed to remove the variation between replicates using the “ComBat” algorithm from the Sva package (version 3.44.0). Later, the batch-corrected data were normalized by smooth quantile normalization technique using the qsmooth package (version 1.12.0) (Hicks et al., 2018).
The p-value by Student's t-test and fold change values by taking an average of technical replicates were calculated. Differentially expressed proteins in cells treated with human and Wba-MIF at 5 and 15 min time points with respect to control (5 min) and h-MIF versus Wba-MIF treatment at 5 and 15 min were calculated. In this study, proteins/phosphoproteins with fold change value s ≥ 1.25 and ≤0.8 were considered as upregulated and downregulated. All the statistical analysis were calculated using in-house R scripts (version 4.1).
Availability of mass spectrometry data
The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium through the PRIDE (Perez-Riverol et al., 2022) partner repository with the data set identifier PXD048133.
Results
Wba-MIF2 expression, purification, endotoxins removal and enzyme activity
Recombinant Wba-MIF2 was expressed in Escherichia coli (Rosetta BL21) and purified using an IMAC column. A gradient concentration of imidazole (0–400 mM) was used for the elution of the Wba-MIF2, and 250 nM was found to be optimum. Purified protein Wba-MIF2 was separated on 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis for its purity check, and a single of 17 kDa was observed (Supplementary Fig. S1), indicating the purity. Endotoxins from the purified protein were removed, and when tested, their concentration was less than the permissible limit (0.2 EU/mL).
The recombinant Wba-MIF2 was tested for its oxidoreductase and tautomerase activities, and it was found to possess oxidoreductase activity (Supplementary Fig. S2A), as well as tautomerase activity (Supplementary Fig. S2B), thus indicating that the purified recombinant Wba-MIF2 is functionally active. The oxidoreductase activity of Wba-MIF2 was about twofold compared with DTT + Insulin control at 60 min of reaction. Similarly, the tautomerase activity was about fourfold compared with the control after 5 min of reaction.
Signaling responses mediated by Wba-MIF2 in RAW 264.7 macrophage cells
In this study, we utilized an LC-MS/MS-based phosphoproteomics approach to identify the signaling mechanism involved in Wba-MIF2-mediated signaling in RAW 264.7 macrophage cells. We also compared signaling between h-MIF and Wba-MIF2. The comprehensive workflow utilized in this study is depicted in Figure 1A.

Wba-MIF2-mediated signaling.
The signaling response mediated by Wba-MIF2 was measured in macrophages. To unveil the time-dependent impact of Wba-MIF2 on macrophages, we investigated the changing phosphorylation patterns of established signaling molecules, specifically p-44/42 MAPK (pERK) and pAKT, at five different time intervals after Wba-MIF2 stimulation. Increased phosphorylation of 44/42 MAPK and AKT was observed at 5 min, which decreased at 10 min. However, an increased phosphorylation was observed at 15 and 30 min. The peak phosphorylation of 44/42 MAPK was observed at 5 min in Wb-MIF2 and h-MIF stimulated cells. However, the peak phosphorylation of AKT was observed at 5 min in response to h-MIF and at 30 min in Wba-MIF2. Based on these results, we chose 5 and 15 min for our subsequent experiments (Fig. 1B).
Quantitative investigation of temporal changes in protein phosphorylation in response to Wba-MIF2 stimulation
We conducted a multiplexed quantitative phosphoproteome analysis to gain insights into protein dynamics and examine the signaling responses influenced by Wba-MIF2. In addition, we also compare the signaling response by Wba-MIF2 and h-MIF. RAW 264.7 macrophages were treated with Wba-MIF2 and h-MIF at different time points (0, 5, and 15 min). Upon treatment, cell samples were lysed in a lysis buffer having SDS, protease inhibitors, and phosphatase inhibitors.
Subsequently, the samples underwent trypsin digestion and a C18 purification step. Equal quantities of peptide digests from each time point were labeled using a six-plex TMT labeling technique. After this, phosphopeptides were enriched through TiO2-affinity purification, and the resulting enriched phosphopeptides were subjected to analysis using an Orbitrap Fusion mass spectrometer.
The database searches of the raw files obtained from LC-MS/MS analysis of the RAW 264.7 macrophages treated with Wba-MIF2 and h-MIF protein led to 0.74 million (747,866) MS/MS scans from the total proteomic experiment resulted in the identification of 0.25 million (252,151) PSMs, and 19,505 peptides corresponding to 3545 proteins. Similarly, the 0.67 million (676,918) MS/MS scans from the phosphoproteomics experiment resulted in the discovery of 38,604 PSMs and 3488 peptides correlating to 1184 proteins.
Based on the ptmRS best site localization probability cutoff of ≥75%, we observed 1201 phosphorylated sites on 467 proteins. Out of 1201 phosphorylated sites, 1075, 117, and 9 were found on serine (S), threonine (T), and tyrosine (Y) residues, respectively (Fig. 1C). Furthermore, relative quantification was performed on 3253 proteins from the total proteomic experiment and 1150 phosphopeptides.
Our mass spectrometry phosphoproteomics data revealed that all the phosphopeptides identified across all the conditions mapped to five clusters and are differentially expressed (Fig. 2A). We also identified differentially expressed phosphoproteins upon Wba-MIF2 treatment. Among the 3488 phosphopeptides identified, 54 peptides were significantly hyperphosphorylated (≥1.25-fold), and 15 peptides were significantly hypophosphorylated (≤0.80-fold) corresponding to 54 and 15 proteins, respectively, upon Wba-MIF2 treatment of RAW 264.7 cells.
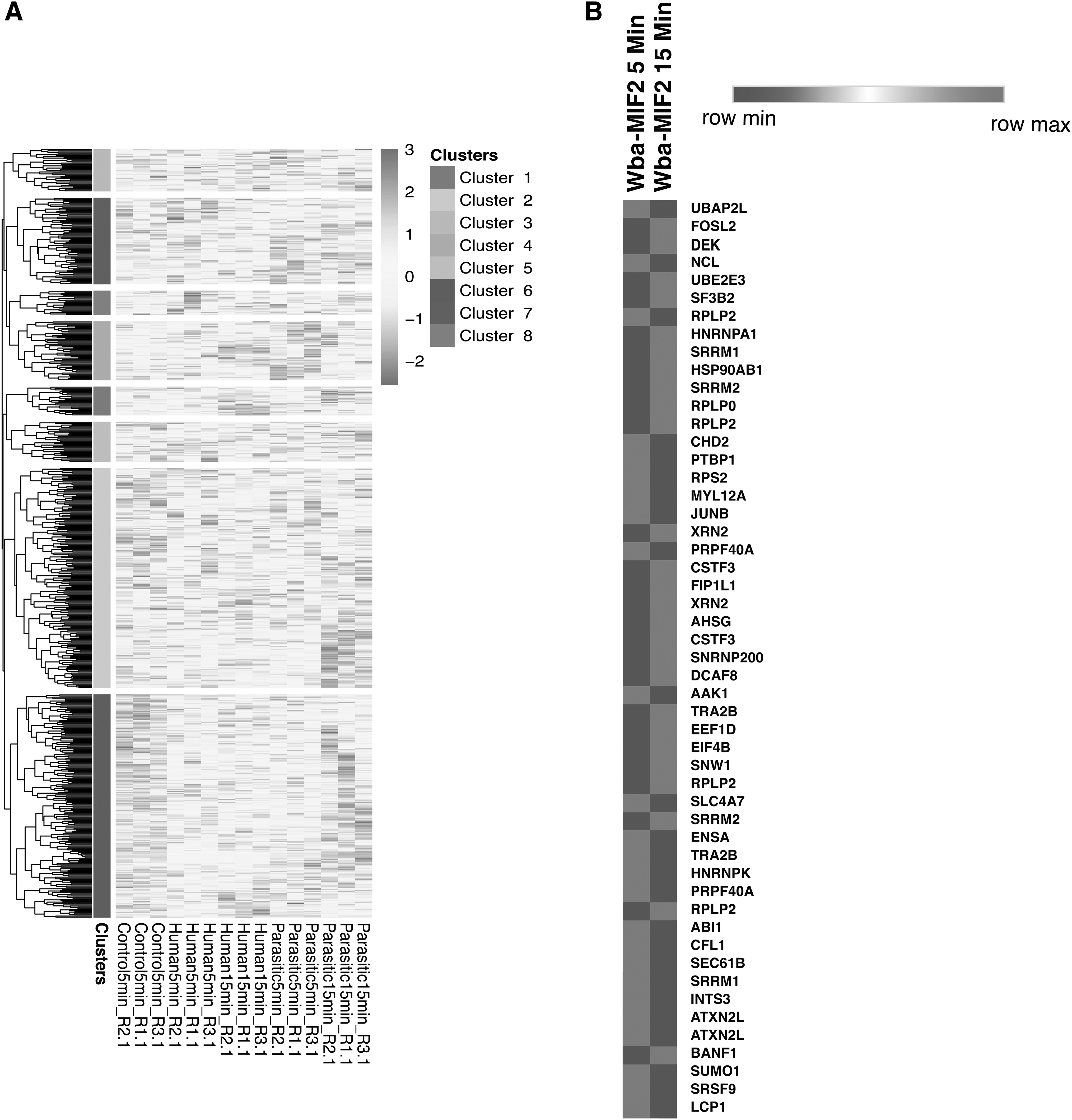
MIF differentially regulates phosphoproteins.
Of these significantly altered proteins, 17 were hyperphosphorylated, and 3 were hypophosphorylated at 5 min. Similarly, 37 proteins were hyperphosphorylated, and 12 were hypophosphorylated at 15 min of Wba-MIF2 induction. Supplementary Table S1 provides detailed information about dysregulated proteins upon Wba-MIF2 and h-MIF. Supplementary Tables S2–S4 contain a complete list of identified and altered phosphopeptides upon Wba-MIF2 treatment and comparison with h-MIF, respectively. The heatmap illustrates the noteworthy changes in phosphoproteins after exposure to Wba-MIF2 treatment (Fig. 2B).
Functional and subcellular classification of phosphoproteins identified as targets of Wba-MIF2
Phosphoproteins have a role in the structural and functional characteristics of cells and tissues and the coordination of several metabolic and signaling pathways. We performed functional annotation of identified phosphoproteins using the PANTHER tool to categorize phosphoproteins based on their biological processes, molecular functions, and subcellular localization to understand better the functionality of the identified dysregulated phosphoproteins in response to Wba-MIF2 in macrophages.
The subcellular localization analysis illustrated in Supplementary Figure S3 and Supplementary Table S6 revealed that a majority of the phosphoproteins belonged to the cytosol (GO:0005829) (60%), extracellular exosome (GO:0070062) (34%), nucleoplasm (GO:0005654) (39%), cytoplasm (GO:0005737) (49%), membrane (GO:0016020) (34%), nucleus (GO:0005634), (46%), endoplasmic reticulum (GO:0005783) (10%) and ribosome (GO:0005840) (5%).
The analysis of the biological process in Supplementary Figure S3 and Supplementary Table S5 revealed a majority of the phosphoproteins to be involved in the cytoplasmic translation (GO:0002181) (6%), translation (GO:0006412) (8%), mRNA splicing (GO:0000398) (10%), and RNA processing (GO:0006364) (7%). Apart from these, they are also involved in the tricarboxylic acid cycle (GO:0006099), intracellular protein transport (GO:0006886), protein transport (GO:0015031), cytoskeleton organization (GO:0007010), etc.
The molecular functional analysis in Supplementary Figure 3 and Supplementary Table S7 pointed out that the majority of phosphoproteins belonged to protein binding (GO:0005515) (89%), RNA binding (GO:0003723) (34%), ATP-binding (GO:0005524) (14%), protein binding (GO:0042802) (14%), DNA binding (GO:0003677) (9%), cadherin binding (GO:0045296) (8%), ATPase activity (GO:0016887) (6%), and protein homodimerization activity (GO:0042803) (6%) among others.
Wba-MIF2 is associated with vital cellular functions.
Pathway enrichment analysis
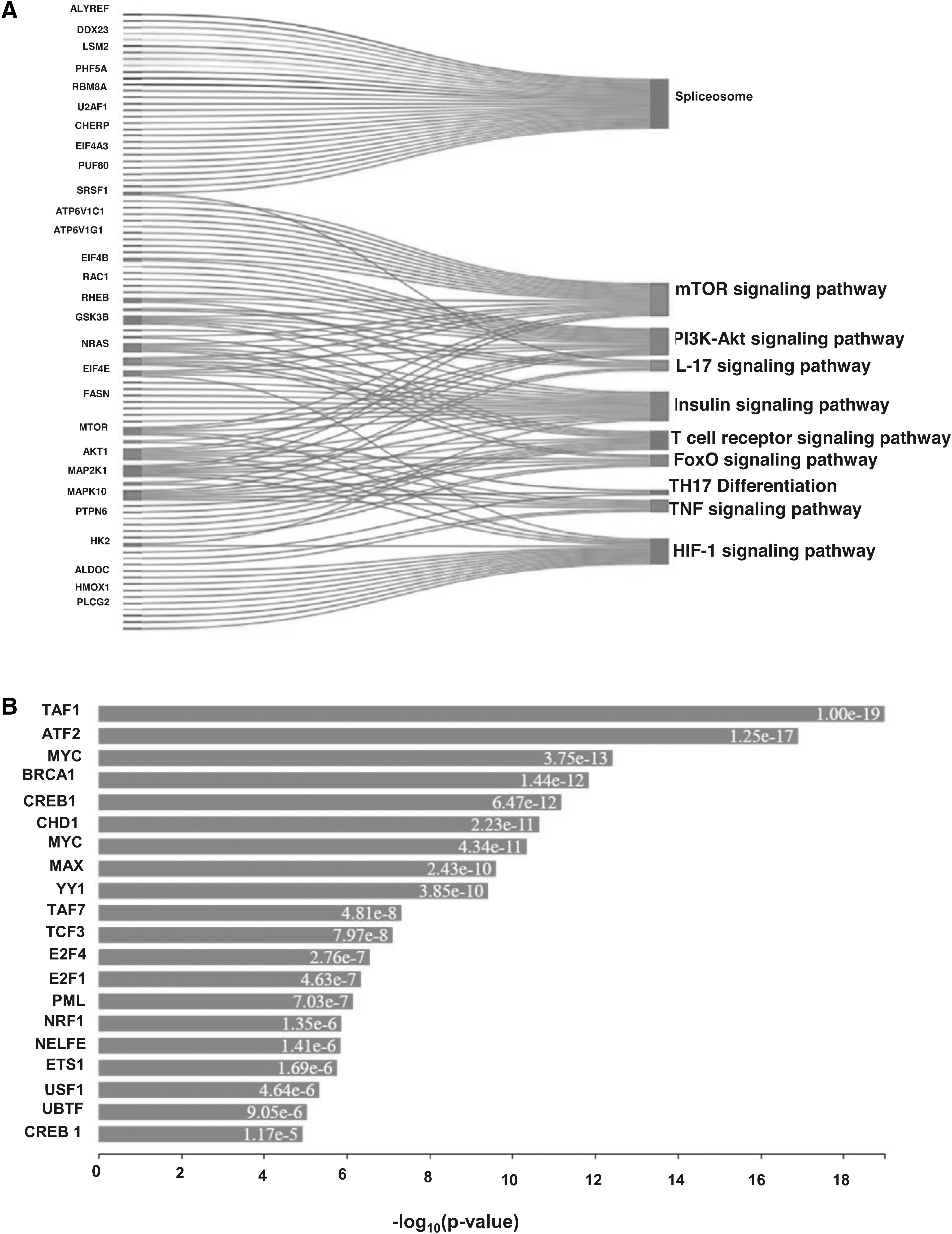
Protein dynamics is a key factor in regulating several cellular pathways. We conducted pathway enrichment analysis using both David and Panther tools. We identified several significant pathways (p < 0.05) (Fig. 3A and Supplementary Table S8). We have shortlisted essential pathways such as spliceosome (Supplementary Fig. S5), mRNA surveillance pathway, interleukin (IL)-17 signaling pathway, vascular endothelial growth factor signaling pathway, sphingolipid signaling pathway, insulin signaling pathway, PI3K-Akt signaling pathway, and mTOR signaling pathway. The proteins involved in these pathways are given in Supplementary Table S8.
Transcription factor enrichment
Several proteins called transcription factors are involved in the process of transcription of DNA. Numerous phosphoproteins serve as transcription factors. We enriched transcription factors using the online eXpression2Kinases (X2K; http://amp.pharm.mssm.edu/X2K/) tool to gain more insight into Wba-MIF2-mediated signaling. Our analysis leads to the enrichment of several transcription factors, a few of them are TAF1, ATF2, MYC, BRCA1, CREB1, CHD1, MYC, MAX, YY1, TAF7, TCF3, E2F4, E2F1, PML, NRF1, and NELFE (Fig. 3B). A detailed list of transcription factors enriched in the study is provided in Supplementary Table S9.
Kinase enrichment analysis
In addition to the enrichment of transcription factors, we conducted kinase enrichment analysis using X2Kweb, an online tool (Clarke et al., 2018) for identifying dysregulated kinases after Wb-MIF2 treatment. X2Kweb employs computational methods to predict the involvement of upstream cell signaling pathways. Our analysis revealed an enrichment of CDK5, CDC2, DNAPK, and CDK2 as downstream kinases in the Wb-MIF2-induced signaling cascade. These findings suggest that the signaling cascade mediated by Wba-MIF2 in macrophages may involve CDK5, CDC2, DNAPK, and CDK2, subsequently engaging MYC, TCF3, MAX, ATF2, TAF7, YY1, CREB1, BRCA1, and TAF1 transcription factors (Fig. 4A and Supplementary Table S10).
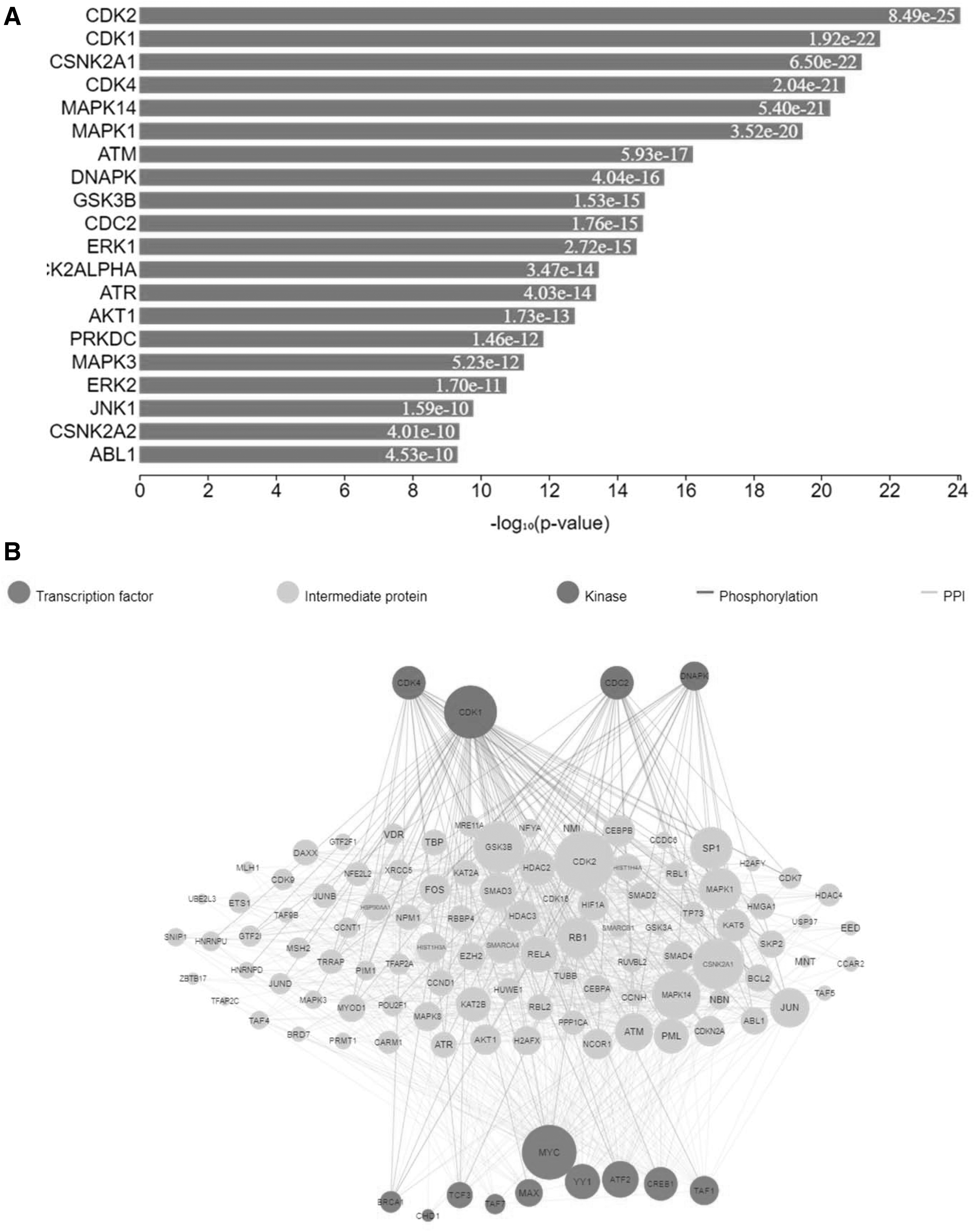
As depicted in Figure 4A, kinases are enriched among the identified phosphoproteins in macrophages exposed to Wba-MIF2. In Figure 4B, the expression kinases analysis of phosphorylated proteins presents a network illustrating the enriched kinases, protein–protein interactions, and transcription factors within the phosphorylated data set of macrophages treated with Wba-MIF2.
Comparison between h-MIF and W. bancrofti MIF (Wba-MIF2)
As a pleiotropic protein, macrophage migration inhibitory factor (MIF) contributes to inflammatory and immunological reactions. MIF was initially identified as a lymphokine involved in tumoricidal action, delayed hypersensitivity, and several macrophage-related activities, such as phagocytosis and dissemination. In this study, we compared signaling mediated by Wb-MIF2 and h-MIF to see the efficiency of modulating immunological response compared with h-MIF. We have compared the phosphorylation status of the proteins dysregulated by h-MIF and Wba-MIF2.
We also compared differentially expressed proteins from treatment with Wba-MIF2 and h-MIF. Our results show that h-MIF and Wba-MIF2 regulate the exact signaling mechanism; however, Wba-MIF2 showed some advantages over h-MIF. Supplementary Table S3 shows the number of differentially expressed proteins in each comparison. Supplementary Figure S4 represents the heat map corresponding to the expression of proteins and phosphoproteins in various conditions.
Discussion
Nematode infections are common and require novel molecular targets for diagnostics and therapeutics innovation. Lymphatic filariasis stands out as a major contributor to disability in tropical regions. This study sought to unpack immune modulation during nematode infection as a site of potential therapeutics innovation using temporal quantitative phosphoproteomics profiling of the macrophage MIF2.
Systemic and local inflammatory and immunological responses are greatly influenced by the Macrophage MIF protein. MIF has been implicated in the pathogenesis of various autoimmune diseases, including T1D (Saunders et al., 2007), multiple sclerosis, autoimmune hepatitis, and rheumatoid arthritis (Hjæresen et al., 2022). The pleiotropic cytokine (h-MIF) has two enzymatic activities, tautomerase, and oxidoreductase, in addition to its cytokine properties (Kudrin et al., 2006).
Importantly, MIF is also found in several parasites, including W. bancrofti, the major lymphatic filarial parasite (Chauhan et al., 2015; Sharma et al., 2012). Likewise, parasitic MIF (Wba-MIF2) has two enzymatic functions (Chauhan et al., 2015). There is currently sufficient data to conclude that h-MIF is crucial in autoimmune conditions such as diabetes mellitus, ulcerative colitis, arthritis, and immunomodulation (Bilsborrow et al., 2019; Calandra and Roger, 2003; Hao et al., 2013; Sanchez-Zamora and Rodriguez-Sosa, 2014).
Since parasite MIF homolog (Wba-MIF2) also possesses enzymatic functions, it also plays a significant role in immunomodulation (Chauhan et al., 2015; Hoerauf et al., 2005; Karabowicz et al., 2022; Mukherjee et al., 2019; Sharma et al., 2012); however, its molecular mechanism is less explored. Therefore, we conducted this study to delineate the signaling mechanism involved in the immunomodulation by the Wba-MIF2 and compare it with that of the h-MIF.
Our findings collectively indicate that Wba-MIF2 can activate signaling cascades, such as MEK-ERK and AKT signaling pathways, in a time-dependent manner. Based on this observation, we have selected 5- and 15-min time points for further experiments.
Wba-MIF2 showed a vigorous indication of phosphorylated proteins across these two time points. We observed hyperphosphorylation in several proteins, including ubiquitin-conjugating enzyme E2 E3 (UBE2E3), which is involved in various cellular activities, including DNA binding, cytoskeletal motor activity (Berndsen and Wolberger, 2014), and immune system process (Loureiro and Ploegh, 2006; Singh et al., 2023).
SF3B2 was hyperphosphorylated at 15 min at serine 430, 434, and 435. Generally, this protein serves as an anti-inflammatory and brings about immunomodulation in neurons (Jeong et al., 2023). Our recent studies have shown that Wba-MIF2 could modulate the host immune system in the T1DM mice model from a pro-inflammatory to an anti-inflammatory milieu (Personal communication; Singh et al., 2024). After 5 weeks of wb-MIF2 treatment, a significant (p < 0.05) downregulation of pro-inflammatory cytokines, IL-1β IL-2, IL-6, tumor necrosis factor (TNF)-α, and interferon (IFN)-γ and a significant (p < 0.05) upregulation of anti-inflammatory cytokines, IL-4, and IL-10 were observed in the diabetic animals, as compared with vehicle-treated groups.
The pattern of expression of genes of pro-inflammatory and anti-inflammatory cytokines in splenocytes reflected the levels of respective cytokines in the blood of animals. IL-4 and IL-10 gene expression levels were significantly increased in spleen tissue, but IL-1β, IL-2, IL-6, IFN-γ, and TNF-α gene expression levels were markedly decreased due to Wba-MIF2 treatment (Personal communication; Singh et al., 2024).
RPLP2 is crucial for the elongation phase of protein synthesis. Increasing or reducing RPLP2 expression can change the pace of protein translation, affecting how quickly cells proliferate (Chen et al., 2002). Our mass spectrometry-based phosphoproteomics data revealed that Wba-MIF2 promotes the phosphorylation of RPLP2 at serine 303/306. It may be possible that these phosphorylation sites of RPLP2 may induce Wba-MIF2-mediated cell proliferation.
PTBP1 mediates several biological activities in some cell types, including immune cell activation, neuronal cell development, and differentiation. To control the expression of CD40 ligand (CD40L) and IL-2, PTBP1 regulates the cluster of differentiation 4 T cell activation by activating the nuclear factor-B and phospholipase C-1 (PLC-1)/extracellular signal-regulated kinase 1/2 (ERK1/2) pathways (La Porta et al., 2016). Our results identified hyperphosphorylation of PTBP1 at serine 141 and 142. It suggests that this protein may be involved in Wba-MIF2-mediated immunomodulation.
Our gene ontology and pathway enrichment results showed that most enriched phosphoproteins are involved in cytoplasmic translation, mRNA splicing, and RNA processing. The molecular functional enrichment analysis indicates that most phosphoproteins have protein binding, RNA binding, and DNA binding properties. Our pathway enrichment results showed enrichment of several key pathways, including spliceosomes, T cell receptor signaling pathway, Th17 differentiation pathway, IL-17 signaling pathway, and insulin signaling pathway. These observations show that Wb-MIF2 activates signaling pathways through modulating RNA-binding proteins and spliceosome machinery and may bring immunomodulation (Fig. 5).
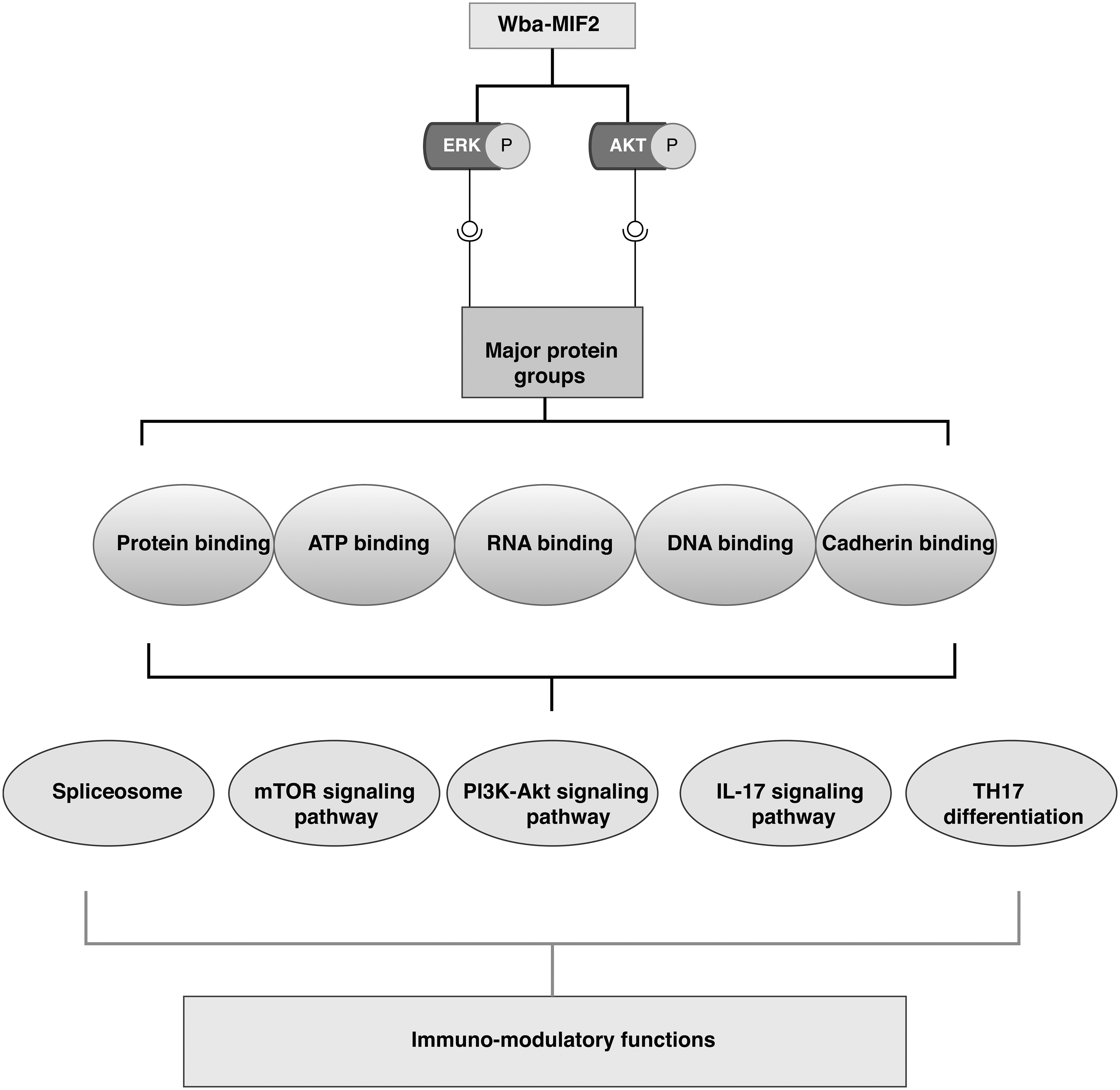
Wba-MIF2 contributes to immunomodulation by activating various signaling molecules, such as ERK and AKT. These signaling molecules are categorized into different protein groups, including those involved in protein binding, ATP binding, RNA binding, DNA binding, and cadherin binding. The phosphorylated proteins are likely associated with multiple pathways, encompassing spliceosome, mTOR signaling, PI3K-AKT, IL17, and TH17 differentiation pathways. Changes in these pathways may underlie the immunomodulatory functions of Wba-MIF 2
We further carried out several analyses to gain deeper insights into the Wba-MIF2-mediated signaling mechanism in macrophages. Our transcription factor enrichment analysis identified several downstream transcription factors involved in gene transcription in various cellular events. CREB1 has diverse cellular functions, including proliferation, survival, and differentiation of the cells. Immediate early genes transcriptional induction is one of the crucial functions of CREB as a transcription factor and master regulator of anti-inflammatory immune responses. These genes contain nuclear receptors, cytokines, chemokines, growth, and transcription factors (Wen et al., 2010).
Another transcription factor that is enriched in our study is MYC. MYC controls a wide range of genes in both innate and adaptive immune cells, regulating their activation, proliferation, polarization, and subsequent functional events that these cells generate (Gnanaprakasam and Wang, 2017). ATF2 is another transcription factor that, upon activation, stimulates the transcription of several genes involved in immune responses and immunomodulation. It regulates the expression of cell adhesion molecules, pro-inflammatory cytokines, and chemokines (Yu et al., 2014).
Our analysis of Wba-MIF2-mediated dysregulated phosphoproteins identified several upstream kinases, including CDK4, CDK1, and DNAPK. DNA-dependent protein kinase catalytic subunit (DNAPK or PRKDC) is a kinase that is a component of the DNA-dependent protein kinase (DNA-PK) complex, which is essential for V(D)J recombination and the repair of DNA double-strand breaks. It also promotes central T cell tolerance (Mathieu et al., 2015).
CDK1 and CDK4 are involved in developing innate immune system cells and maintaining balanced innate immunological activity. Type I IFN (IFN−) secretion was suppressed in monocytes, thus modulating the immune system. Macrophage differentiation and activity depend on CDKs (Laphanuwat and Jirawatnotai, 2019). We also compared the immunomodulatory properties of Wba-MIF2 with h-MIF, as h-MIF is a well-established molecule that brings about immunomodulation (Calandra and Roger, 2003).
Immunomodulation is a complex process and mechanistic studies can lend evidence for new targets and therapeutics innovation. Both MIF bring about immunomodulation by the same molecular mechanism and involvement of the same signaling pathways. However, further studies are required to understand how the role of a parasite MIF and h-MIF differs in immunomodulation.
Conclusions
This study's findings based on global phosphoproteomics support the idea that unpacking immunomodulation attendant to W. bancrofti can serve as a site of research and innovation for molecular targets. We identified 467 phosphorylated proteins upon Wba-MIF2 treatment of macrophages RAW 264.7 cells. Our bioinformatics analysis shed light on the majority of these phosphoproteins involved in RNA binding and spliceosome machinery and involved in immunomodulation.
We further compared data obtained from Wba-MIF2 and h-MIF and concluded that both act through the same mechanism. The present data from in vitro studies and our recent data on the effect of Wba-MIF2 in T1D animal models (Personal communication; Singh et al., 2024) show that Wba-MIF2 plays a role in immunomodulation. These data inform future diagnostics and drug discovery research for W. bancrofti and call for further research for validation and confirmation of Wba-MIF2 as an important immunomodulator.
Footnotes
Acknowledgments
We thank the Indian Council of Medical Research for the ad hoc project (No. 2015-1226-BMS) and the Government of Karnataka for funding Yenepoya (Deemed to be University) under the Biotechnology Skill Enhancement Programme (BiSEP). The authors thank the Director ICMR-NITM for encouragement and support. A.K. is a recipient of a senior research fellowship from the University Grants Commission (UGC), Government of India.
Authors' Contributions
S.L.H, P.K.M., and T.S.K.P. conceptualized the study, designed the workflow, and edited the article. P.K.M. and I.S. prepared the illustrations and wrote the drafts of the article. A.K. C.N.K., I.S., and P.K.M. carried out experiments, obtained data, and analyzed proteomic data. C.N.K. carried out bioinformatics analysis. All authors made a significant intellectual contribution on the article drafts, read, edited, and approved the final version of the article.
Authors Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
Indian Council of Medical Research Grant no: 2015-1226-BMS.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.