
Abstract
Newborns and young children rely on innate immunity to protect against infections until the adaptive immune system matures. Immunization helps facilitate protection, but multiple doses are needed to establish sufficient antibody levels and T-cell-facilitated immune memory. Deficient T-cell activation and function among neonates and young children are primarily present in the CD4+ compartment, whereas CD8+ T-cell function is at par with adults. CD4+ T cells in neonates and young children produce low levels of IFNγ, interleukin (IL)-2, IL-13, IL-5, and IL-17. This inherent deficiency in neonatal and young child CD4+ T-cell functionality has been linked to several mechanistic failures: (1) lower sensitivity to T-cell receptor stimulation, (2) increased apoptosis after proliferation, (3) unavailability of antigen for T-cell priming, and (4) inefficient stimulation by relatively immature antigen-presenting cells. In this review, we discuss evidence from infection and vaccination responses that shed light on the various checkpoints possibly involved in delayed maturation of CD4+ T-cell activation and function in newborns and young children.
Introduction
This review will focus on the knowledge regarding deficiencies in the CD4+ T-helper cell arm of the immune system of neonates and young children. Several studies, including ours, using infection models and vaccination have demonstrated reduced, impaired, or different polarization of CD4+ T-cell responses among neonates and young children compared to adults, while function of their CD8+ T-cells remains intact.3–13 The mechanisms responsible for impaired CD4+ T-cell function in newborns and young children involve multiple factors.1,2,8,14–28 (1) Immunologically immature APCs may fail to stimulate T-cells to the same extent as observed in adults2,17,18; (2) maternal antibodies may interfere with normal processing of antigen19,20; (3) lower Toll-like receptor (TLR) expression may impact T-cell activation14,21,22; (4) a predilection to Th2-dominated responses may result in poor T-cell-mediated protection from infections9,23–26; or (5) regulatory T-cells may mediate suppression of CD4+ T-helper cell activation.27,28
Dysfunction in the T-Cell Receptor-Signaling Cascade
Experiments with umbilical cord blood (UCB) T-cells have facilitated most of our understanding of the intrinsic T-cell defects among neonates. Neonatal T-cells exhibit reduced responses to anti-CD3 or anti-CD2 stimulations compared to adult T-cells (Fig. 1, Table 1). 29 This defect associates with an inherent inability to induce protein phosphorylation of signaling molecules downstream from the T-cell receptor (TCR), characterized by impaired activation of phospholipase C-γ, reduced basal levels of Lck, and less phosphorylation of ZAP-70. 30 This dysfunction in the TCR-signaling cascade does not affect neonatal T-cell proliferation. CD4+ CD45+-sorted naïve T-cell populations from UCB as well as adult blood have been shown to proliferate similarly in the presence of interleukin (IL)-2 and a costimulatory signal. 31 However, unlike adult T-cells, UCB CD4+ CD45RA+ T-cells demonstrate extensive apoptosis with bystander increases in CD95L levels compared to adult blood. 31 Thus, after anti-CD3 stimulation, IL-2 produced by UCB CD4+ T cells is insufficient for autocrine signaling to sustain neonatal CD4+ T-cell proliferation. 32
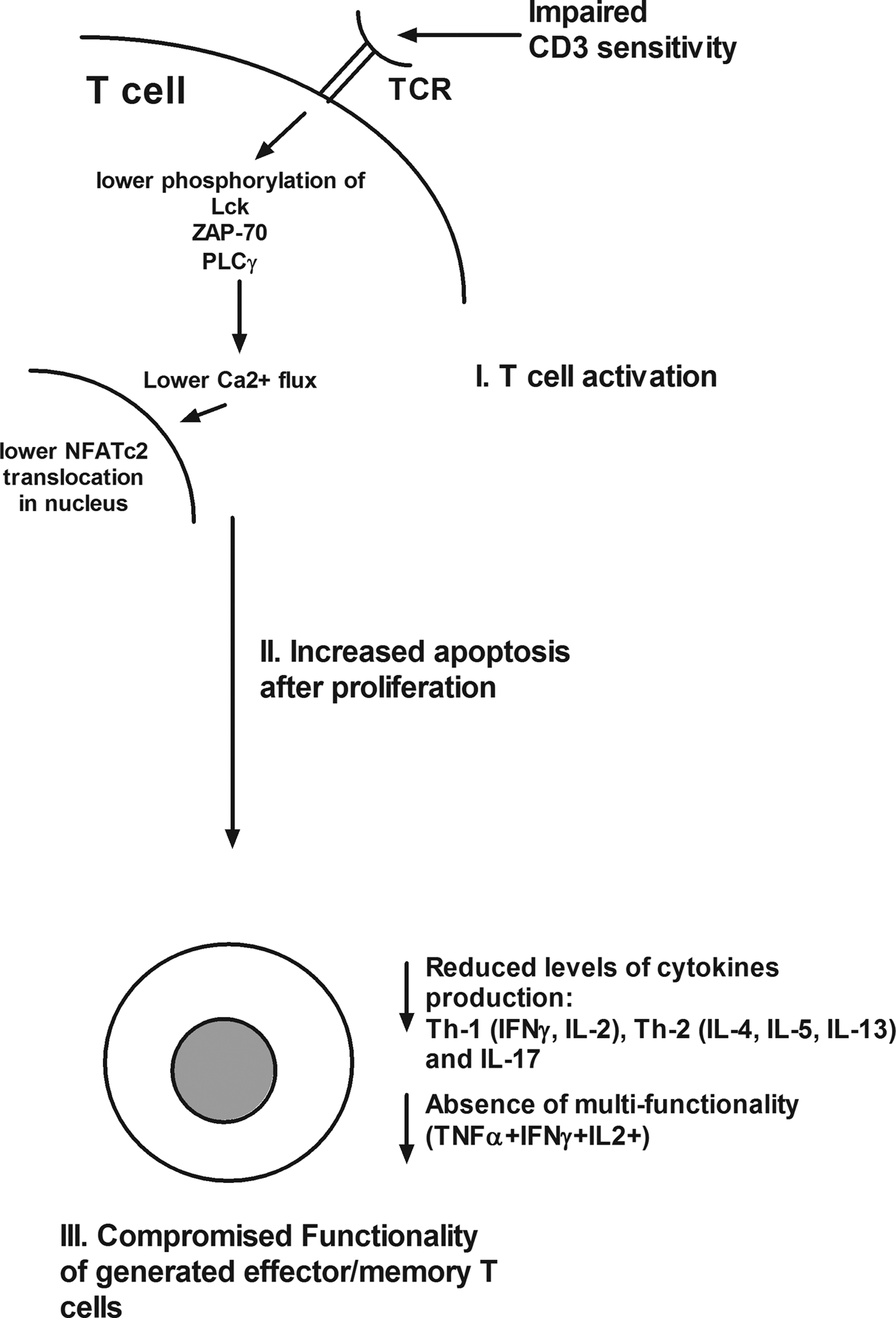
Shortcomings of neonatal and young child T-cells. Schematic of checkpoints that occur at various stages in neonatal and young child T-cell activation starting with T-cell receptor (TCR)-mediated activation. T-cells have reduced CD3 sensitivity and lower phosphorylation of several TCR-signaling molecules resulting in low Ca2+ flux induction and reduced NFATc2 translocation in the nucleus. After activation, neonates and young child T-cells demonstrate comparable levels of cellular proliferation to adults' T-cells. However, neonatal and young child T-cells are prone to apoptosis postactivation. The limited frequencies of activated T-cells that result in effector/memory generation do not produce optimal levels of cytokines and lack multifunctionality.
STAT4, signal transducers and activators of transcription 4; TCR, T-cell receptor.
Increased Apoptosis of Proliferated Naïve Neonatal CD4+ T Cells
Increased apoptosis of proliferated naïve neonatal CD4+ T-cells could be a contributing factor to their inability in generating a sufficient effector/memory CD4+ T cell pool rather than dependent entirely on APC inefficiency (Fig. 1, Table 1). 33 However, under certain stimulatory circumstances, anti-CD3/anti-CD28 and naïve UCB CD4+ T cells can generate competent effector cells. 34
CD154 (CD40L) Surface Expression on CD4+ T-Cells
CD154 (CD40L) surface expression on CD4+ T-cells and its interaction with CD40 present on B-cells are vital for Ig isotype switching, antibody production to T-cell-dependent antigens, and memory B-cell generation. 35 Neonatal CD4+ T-cells display lower CD154 mRNA levels and reduced surface CD154 expression compared to adult CD4+ T-cells. Consequently, neonates have reduced CD4+ T-cell-mediated B-cell help to pathogens and vaccines, making them more susceptible to respond to infections and more difficult to immunologically prime with vaccines, in parallel to CD154-deficient patients.36–39 Mechanistically, decreased CD154 expression by neonatal CD4+ T-cells after TCR-CD3 engagement results in lower calcium flux induction, a key event for the CD154 gene transcription (Fig. 1, Table 1). Reduced CD154 promoter activity in neonatal T-cells persists when proximal signaling events are bypassed using PMA/ionomycin, suggesting an additional and more distal mechanism for decreased transcription. In contrast, CD154 mRNA stability is similar in neonatal and adult T-cells. 38 Affymatrix oligonucleotide microarrays have been used to compare gene expression of purified CD4+ UCB T-cells to adult CD4+ T-cells after anti-CD3 stimulation. In that study, nuclear factor of activated T-cell (NFAT)-regulated genes exhibited lower expression in UCB CD4+ T-cells, including several transcription proteins of the NFAT pathway viz. C/EBPβ, JunB, and Fosl1. The NFAT family of transcription factors that operate downstream of the Ca2+-signaling cascade contains distinct proteins such as NFATc2, a critical transcription factor necessary for upregulation of multiple cytokines.40,41 As a consequence of diminished NFATc2, key cytokines IFNγ, TNFα, IL-2, IL-4, IL-5, and IL-13, and other factors viz. CD154 and macrophage inflammatory protein 1a (MIP-1a) are reduced in neonatal CD4+ T-cells (Fig. 1, Table 1). 42
Newborns and Young Children Polarize Their CD4+ T-Cell Responses to Th-2
Based on their functional properties, CD4+ T-cells can be categorized as Th-1, producing mainly IFNγ, TNFα, and IL-2 or Th-2, producing IL-4, IL-5, IL-10, and IL-1. 43 Newborns and young children polarize their infection- and vaccine-mediated CD4+ T-cell responses to Th-2.9,26 However, neonatal CD4+ T-cells have shown reduced expression of both Th-1- and Th-2-related transcription factors signal transducers and activators of transcription 4 (STAT4), T-bet, and c-maf when compared with adult CD4+ T-cells poststimulation. 42
Studies of Young Child CD4+ T-Cells
There are very few studies of young child CD4+ T-cells because of the difficulty in obtaining sufficient quantities of cells for study. Our group recently found that frequencies of CD4+ T-cells responding to several vaccine candidate proteins expressed by Streptococcus pneumonia and Haemophilus influenzae are very low among infants and young children below 30 months of age. The CD4+ T-cells detected mainly produced IL-2 and very little or no IFNγ [Sharma SK, et al., (submitted)]. When we investigated Th-1 responses to DTaP vaccination, we found the CD4+ T-cells significantly lacked multifunctionality (TNFα+IFNγ+IL-2+). 11 Th-2 responses (IL-4) are also reduced in response to DTaP vaccination of infants. 11 In infants and young children, vaccine antigen- and CMV-specific CD4+ T-cells fail to produce or have reduced IFNγ production.8,11,44,45 This impairment is not solely explained by a reduction in the aforementioned NFATc2 levels, as NFATc2-deficient neonatal mice produce IFNγ to normal levels suggesting some alternate deficiencies in neonatal CD4+ T-cells. 46 Reduced percentages of IFNγ-producing CD4+ T cells among preterm infants are observed after polio vaccination. 6
Maturation of IFNγ production by CD4+ T cells comes with age. After the neonatal period, IFNγ-producing T cells increase over time in response to different stimuli.47,48 An age related increase in the frequency of CD4+ T-cells producing IFNγ can be seen in response to Staphylococcus enterotoxin B between age 2 months and 11 years. 49 Antigen-specific expression of CD69, an early activation marker that initiates Ca2+ flux, signal transduction, and cytokine production in T-cells, is similarly expressed on activated CD4+ T cells among infants and adults.11,49 Despite this similarity in CD69 expression, pathways to achieve full IFNγ effector function of CD4+ T cells do not mature until the age of ∼10 years. 49 Nevertheless, in some cases among older children (median age 5 years), antigen-specific CD4+ T-cell IFNγ function develops that can reduce S. pneumoniae nasopharyngeal colonization.10,50 In addition, certain antigens derived from Mycobacterium spp. have the ability to induce efficient IFNγ production among neonates.7,51 Further, in young children prone to repeated otitis media infections, we observed that nasopharyngeal colonization with Spn or nontypeable H. influenzae (NTHi) produces lower frequencies of IFNγ-producing CD4+ T-cells compared to those who are not prone to otitis, they may display a more neonatal-like immune phenotype. 13 Besides these intrinsic defects in the T-cells, upstream signals provided by newborns and infants' APCs are also a contributing factor responsible for defective T-cell response generation among this group.
Impaired Contribution of APCs in Activation of CD4+ T-Cells
APCs serve as the interface between innate and adaptive immunity by facilitating T-cell activation and responses. 52 Neonates and infants have less-robust T-cell responses to viral or bacterial infections and to vaccination, because they have immature APCs.2,18 The extent to which innate immune responses to microbial stimuli differ between neonates and infants compared to older children and adults are open questions.17,53 Microbial stimuli and vaccines trigger innate immunity by stimulating TLRs on the surface of APCs (dendritic cells, monocytes, and macrophages).54,55 APCs direct the subsequent adaptive immune response after integrating information from TLRs. TLR-mediated cytokine responses in UCB are distinct from those of adults. 21 In response to TLR ligand stimulation, neonatal CD4+ T-cells produce less IL-1α, IL-1β, TNFα, IL-18, and IL-12p70, but comparable levels of IL-6 and IL-10 compared to adult CD4+ T- cells.56,57 While adult and neonatal monocytes have similarity in expressing CD14 and TLR4 on their surface, LPS-mediated TLR4 responses in UCB monocytes are less potent in inducing TNFα secretion. Although there is an equal phosphorylation of ERK, p38, and IκBα phosphorylation, the myeloid differentiation antigen 88 (MyD88), a TLR adaptor protein, is reduced in neonatal monocytes, 58 a direct correlation between adult and newborn. Since TLR-4 signals intracellularly through MyD88, it may be that MyD88-dependent factors are responsible for delayed and decreased cytokine secretion in neonatal monocytes. Hence, reduced MyD88 expression may have repercussions in generating antigen-specific CD4+ T-cell responses and is another contributing factor to the neonatal immune immaturity. 59
TLR-Mediated Cytokine Production in Different APC Subsets
More recently, with advanced multicolor flow cytometric analysis, it became feasible to look at TLR-mediated cytokine production in different APC subsets simultaneously (plasmacytoid and myeloid dendritic cells (DC), and monocytes). 21 In response to most TLR ligands, neonatal APCs produce lower levels of IL-12p70 and IFN-α; however, they produce higher IL-1β, IL-6, IL-23, and IL-10 than adult APCs. On a per-cell basis, neonatal APC subsets are less polyfunctional than adult APCs. 21 Thus, the data suggest that neonatal APCs are not less able than their adult counterparts in responding to TLR stimulation, but rather neonatal APCs are qualitatively distinct.
Reduced IL12p70 and Type-I IFNs in Neonates
A profound reduction in IL12p70 may contribute to the lower ability of neonatal CD4+ T-cells to secrete IFNγ. 60 Reduced IL-12p70 by neonatal APCs appears to result in a selective decrease in the production of IL-12p35, but not IL-12/23p40.61,62 Therefore, a dominance of an IL-23 response is found among neonatal APCs stimulated with a range of TLR agonists, including TLR2, TLR 8, and TLR7/8. 21 In some cases, IL-12p40 can also combine with p19 to form the heterodimer IL-23 and may contribute to elevated IL-23 levels in neonatal immune cells.63,64 IL-1β, IL-6, and IL-23 together are key differentiation signaling factors for Th-17-lineage T-cells, suggesting that in the early stages of life, triggering of TLR on APCs favors Th-17 over Th-1 T-cell development. 65 However, there are no reports on T-cell responses that support the notion of Th-17-biased differentiation among neonates.
Absence of IL-17a-Producing CD4+ T-Cells in Infants and Young Children
A role of Th17 cells in providing antibody-independent protection against pneumococcal infection is observed in a murine model,66,67 and Th17 cells can eliminate other extracellular bacterial and fungal infections. 68 However, our data suggest very low frequencies or complete absence of IL-17a-producing CD4+ T-cells in response to individual S. pneumoniae and H. influenzae antigens are observed in infants and young children. 13
Neonatal Monocytes Exhibit Low Baseline Expression of MHCII and Costimulatory Molecules
Naïve T cells require 2 signals that are provided by APCs. 69 Signal 1 is from the pMHC complex expressed by APCs and is recognized by T-cells bearing cognate TCRs. Signal 2 is provided by costimulatory markers (CD80 and CD86) expressed on the APC surface that T-cells sense through CD28. 69 Neonatal monocytes exhibit low baseline expression of the MHCII and costimulatory molecules, CD86 and CD40. Despite APC stimulation by potent activators (combinations of IFNγ and CD154), expression of CD86 and CD40 does not upregulate. 70 UCB-derived myeloid dendritic cells are also functionally immature, exhibiting defective endocytosis, which could result from decreased expression of the mannose receptor.71,72 Lower proliferation is observed in plasmacytoid dendritic cell (DC)-stimulated cultures compared to total UCB-DC-stimulated cultures, which is linked to an increased proportion of apoptotic T-cells in the former cultures. 73 Despite immaturity in neonatal APCs, some infants with congenital HIV infection demonstrate activation of CD4+ and CD8+ T-cells to certain HIV-peptide antigens. 74 Further, when compared with infants born to uninfected mothers, infants born to HIV-infected mothers demonstrate env peptide-specific memory (CD45RO+) CD4+ T-cells. 75 While the use of such peptide antigens bypasses the classical MHC pathway, and hence overcomes APC defects in infants to generate an effective T-cell response, even use of MHC-binding peptides results in suboptimal T-cell activation in infants and young children compared to adults. 8
Poor T-Cell Memory Generation and Phenotypic Divergence of Antigen-Specific T-Cells in Newborns and Young Children
With the intent to establish memory responses to combat infections, vaccination is carried out beginning with neonatal hepatitis B vaccination and at age 2 months for other routine immunizaions. 76 However, the memory phenotype of T-cells developed in newborns is not well defined. Central memory T-cells express CCR7, but not CD45RA, and are likely to represent a long-lived population, which expands rapidly in lymph nodes after antigen encounter.77,78 In contrast, effector memory cells do not express CCR7 and CD45RA (CCR7LowCD45RALow) and act immediately after antigen encounter with a limited proliferative capacity. 77 A third subset, terminally differentiated memory cells, express CD45RA, but not CCR7 (CD45RAHighCCR7Low). Naïve and non-antigen-experienced T-cells characteristically express CD45RA and CCR7.77,79 While these combinations of markers are useful in delineating naïve/memory cells among adults, complexity in the expression of these surface markers exists in infants and young children. 80 Hence, the magnitude and phenotype of pathogen-induced or vaccine-induced memory generation in neonates, infants, and young children are not always similar to what is observed in adults.8,80 Further, based on the aforementioned classification, distinct cytokine patterns are expressed by different phenotypes of T-cells among individual infants. For example, among vaccinated infants, BCG-specific central memory T-cells are more likely to produce IL-2, while effector cells primarily express IFNγ. Altogether, neonatal vaccination with BCG induces a complex pattern of cytokine expression and phenotypes not seen in adults. 80
Expression of Chemokine Receptor CCR7 and CD27
Another classification model based on expression of chemokine receptor CCR7 and CD27 has been described. 79 This allows learning of stepwise differentiation of antigen-specific CD4+ T-cells and is useful to compare early-stage antigen-specific responses. Based on differences in their telomere lengths, in vitro stimulation has demonstrated the order of CD4+ T-cell differentiation as naïve to CCR7+CD27+ to CCR7−CD27+ to CCR7−CD27−. CCR7+CD27+ cells are least differentiated, whereas CCR7−CD27− are fully differentiated CD4+ T cells. We utilized a multiparametric flow cytometry approach and analysis method that allowed simultaneous detection of multifunctionality and phenotypes of CD4+ T-cells induced as a result of DTaP vaccination in infants compared to adults. 11 Functional CD4+ T cells that are DTaP-specific expressed CCR7 and CD27 in infants, which were significantly higher than adults. Adults have significantly higher percentages of CCR7−CD27− cells that are considered as fully differentiated CD4+ T cells. 79 Thus, in infants, the phenotype of DTaP-specific CD4+ T cells is predominantly an early-differentiated phenotype, whereas in adults, CD4+ T cells are fully differentiated (CCR7−CD27−). Similarly, phenotypic changes in influenza vaccine antigen-specific CD8+ T cells among young children and adults are impaired due to maturational delay. 5
Other combinations of surface markers are used to phenotypically define T-cells in infants and young children with CMV infections.81–83 The concentration of undifferentiated T-cells (represented as CD27+CD28+CCR7+) in the peripheral blood is unaffected by CMV; however, there is a large increase in differentiated (CD28−CD57+) CD8+ T-cells with a smaller increase in differentiated CD4+ T-cells. A substantial proportion of infants' antigen-specific CD8+ T-cells that do not express CCR7 still retain CD45RA on their surface, and this differentiated population is prone to senescence. 84 We observed a similar pattern of differentiation among S. pneumoniae and H. influenzae antigen-specific CD4+ T cells among young children. The susceptibility of death may prevent prolonged circulation and further differentiation of these antigen-specific CD4+ T cells. IFNγ-producing CD4+ T cells of older children or adults that are reactive to CMV antigens are found deficient in CCR7 expression (CD45RO+CD45RA−). 8 Most of the CMV-specific IFNγ-producing cells in infants are undifferentiated (revealed by CD27+CD28+), 81 accounting another divergence in the phenotypes of memory T-cells among infants and adults.
Conclusion
In conclusion, several mechanistic failures exist within neonatal and young child CD4+ T-cells. TCR-signaling cascade in T-cells, as well as downstream signaling in neonatal APCs after TLR engagement, is impaired at many checkpoints of the cascade. Since optimal activation of T-cells involves crosstalk with activated APCs by synapse formation, apart from their intrinsic defects within CD4+ T cells, defective neonatal and young child APCs will further fail to induce effector/memory CD4+ T-cell generation after infections or vaccination.
Footnotes
Author Disclosure Statement
No competing financial interests exist.