
Abstract
Transforming growth factor beta 1 (TGFB1) is a multifunctional cytokine with a key role in asthma airway inflammation and remodeling. Since elevated levels of this cytokine in airways might be associated with response to asthma therapy, the aim of this study was to investigate whether the presence of the polymorphism C-509T in the promoter of the TGFB1 gene is associated with response to montelukast. A group of 102 asthmatic patients was genotyped for the presence of the C-509T polymorphism by DNA sequencing and subjected to induced sputum sampling. Cells from sputum samples and BEAS 2B cells were treated with montelukast and endogenous TGFB1 expression was measured by quantitative real-time polymerase chain reaction. The promoter activity was analyzed by luciferase assays in BEAS 2B cells transfected with constructs carrying variants −509C and −509T of the TGFB1 gene promoter. After treatment with montelukast, the decrease in TGFB1 gene expression was greater for the −509TT genotype (58.9%) than for the −509CC and −509CT genotypes (49.6% and 31.8%, respectively) (P = 0.071). In BEAS 2B cells, expression of endogenous TGFB1 was reduced by about 27% after montelukast treatment, while luciferase activity of both promoter variants was increased after montelukast treatment (−509C allele: 48.3%, P = 0.060; and −509T allele: 100.5%, P = 0.062). A more intensive response was registered in the promoter containing the −509T allele, which had 135% higher activity than the −509C variant (P = 0.035). This study showed that the presence of the −509T allele in the TGFB1 promoter might modulate effects of montelukast on TGFB1 gene expression, but future studies are necessary, taking into consideration other genetic and nongenetic factors. It is of potential importance for clinical management of asthma to clarify the influence of the C-509T polymorphism on the response to treatment with montelukast.
Introduction
A
One of several genes implicated in asthma pathogenesis is transforming growth factor beta 1 (TGFB1). This gene encodes an important inflammatory modulator and profibrotic mediator with a key role in remodeling asthmatic airways.6,7 The main sources of TGFB1 in asthmatic lungs are epithelial and inflammatory cells.8,9 Elevated levels of plasma and sputum TGFB1 are observed in asthmatic patients.10–12 The most common polymorphism in the TGFB1 gene promoter, C-509T (rs1800469), is suggested to be associated with increased transcriptional activity and is involved in the pathogenesis of uncontrolled asthma.13,14 It is well studied in different populations and has been investigated as a risk factor for asthma development. 15
Medications used for asthma treatment could also alter the expression of TGFB1.10,11 Direct investigations on the influence of corticosteroids on expression of this cytokine have given contradictory results depending on the experimental model and type of corticosteroids used.16–19 The impact of LTRAs on TGFB1 expression has been rarely evaluated. The effects of TGFB1 on airway remodeling are stimulated by CysLTs and could be attenuated through a receptor-dependent mechanism by montelukast, the most prescribed LTRA for asthma treatment.12,20–23 Montelukast is considered to have additional effects independent of its action against receptors for CysLTs. 24
Asthmatic patients are characterized by elevated levels of TGFB1 in serum and sputum. Administration of LTRAs might stimulate TGFB1 gene expression and potentially increase TGFB1 levels. Overproduction of TGFB1, a potent proinflammatory mediator, could diminish the anti-inflammatory influence of LTRAs. It is therefore necessary to identify genetic factors that may affect the efficacy of LTRAs in asthma therapy.
Since possible synergistic effects of LTRAs and TGFB1 gene variants on expression of TGFB1 have not been examined so far, the aim of this study was to investigate whether the presence of the polymorphism C-509T in the promoter of the TGFB1 gene is associated with response to montelukast treatment.
Patients and Methods
Patients
The study included 102 children and adolescents (age range 5–19 years) with mild and severe asthma. The patients were monitored in the period from 2013 to 2015. Asthma was diagnosed according to the applicable guidelines of the Global Initiative for Asthma (GINA). 2 Skin prick tests for common inhalants and environmental and food allergens were performed according to the European Academy of Allergy and Clinical Immunology (EAACI) guidelines. 25 All patients included in the study had stable asthma at the moment of recruitment. For analysis of correlation between genotypes and phenotypes, patients were divided into 2 groups: mild and severe asthmatics. Patients with mild asthma suffered intermittent symptoms, were treated with a low dose of ICSs, and had a forced expiratory volume in 1 second (FEV1) higher than 80%. Patients with severe asthma had frequent symptoms that were controlled with medium to high doses of ICSs in combination with beta 2 agonists and had impaired lung function (FEV1 < 80% predicted). None of the patients involved in the study required oral corticosteroids. Asthma control was evaluated from the scores of the asthma control questionnaire (ACQ). In addition, analyses of induced sputum and spirometry were used for prediction of future risk of exacerbations. The patients were considered to have well-controlled asthma if they fulfilled the following criteria: an ACQ score below 0.75, normal lung function, and absence of eosinophilia in induced sputum.
This study was in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki Declaration and its later amendments or comparable ethical standards. Written informed consent was obtained from the parents of all patients and the investigation was approved by the hospital ethics committee.
Ex vivo treatment
An induced sputum sample was taken from each patient. Before sputum collection, patients were pretreated with a short-acting beta 2 agonist, salbutamol (200 μg), to prevent bronchoconstriction and spirometry was performed at baseline. Sputum was induced with 4.5% hypertonic saline. Patients inhaled hypertonic saline through an ultrasonic nebulizer in 4 time intervals that were progressively increased for 5 min. After each inhalation period, FEV1 and FVC were measured. Sputum samples were collected into a sterile container.
A set of 22 samples of induced sputum was available to study gene expression in response to montelukast. The sputum samples were processed using a previously described method with some modifications. 26 Sputum cells resuspended in phosphate-buffered saline (PBS) at the end of the procedure were split into 2 equal parts for ex vivo stimulation. The first part was treated with montelukast sodium (100 μM) dissolved and diluted in PBS, while the second part was treated with PBS as the control. Both parts were incubated at 37°C for 30 min and centrifuged at 2,200 rpm for 5 min. The supernatant was discarded, while the cells were resuspended in cold TRI Reagent (Ambion) and stored at −70°C for further ribonucleic acid (RNA) extraction.
Peripheral blood samples from 3 patients with controlled asthma were treated with montelukast in the same way as samples of induced sputum. After lysis of erythrocytes, leukocytes were washed with PBS and resuspended in cold TRI Reagent for RNA extraction.
Analysis of TGFB1 C-509T polymorphism
All patients were genotyped for TGFB1 C-509T polymorphism by direct sequencing. Deoxyribonucleic acid (DNA) was extracted from whole blood using a GeneJET Genomic DNA Purification Kit (Thermo Scientific) following the manufacturer's instructions. The region of the TGFB1 promoter containing polymorphism C-509T was amplified by polymerase chain reaction (PCR). Primers and conditions of PCRs are given in Table 1. Amplified DNA was purified using a GeneJET PCR Purification Kit (Thermo Scientific) according to the manufacturer's protocol. The obtained PCR products (669 bp) were sequenced using an ABI Prism BigDye Terminator Kit (Applied Biosystems) and a 3130 Genetic Analyzer (Applied Biosystems). Sequences were analyzed using the Sequencing Analysis software (Applied Biosystems).
GAPDH, glyceraldehyde-3-phosphate dehydrogenase; PCR, polymerase chain reaction; qRT-PCR, quantitative real-time PCR; TGFB1, transforming growth factor beta 1.
Treatment of BEAS 2B cells
The human bronchial epithelial cell line (BEAS 2B) was maintained at 37°C and 5% CO2 in LHC-9 medium (Life Technologies) supplemented with fetal bovine serum (2%), penicillin (10 U/μL), and streptomycin (10 ng/mL).
The cells were seeded at a density of 3 × 105 in 35-mm cell culture dishes and treated the next day for 1 h with montelukast (100 μM) diluted in medium. Untreated cells served as the control. After treatment, the cells were lysed with TRI Reagent for RNA extraction. The experiment was performed in triplicate.
Analysis of TGFB1 mRNA
Total RNA was extracted from sputum cells, leukocytes, and BEAS 2B cells using TRI Reagent according to the manufacturer's protocol. The RNA concentration and purity were determined by UV absorption spectrophotometry at 260/280. Reverse transcription was performed using a High-Capacity cDNA Reverse Transcription kit (Applied Biosystems) according to the manufacturer's protocol.
Expression of TGFB1 mRNA was measured in triplicate by quantitative real-time PCR (qRT-PCR) using SYBR Green chemistry (Agilent Technologies). Melting curve analysis was performed for all reactions to ensure specificity of the products. For all experiments, glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as the internal housekeeping gene control. The PCR efficiencies of GAPDH and TGFB1 primers were equal. Primers of the genes and the program of amplification are given in Table 1.
A 7500 Real-Time PCR System (Applied Biosystems) was used for qRT-PCR, applying the ddCt method for relative quantification. The TGFB1 expression level was calculated, normalized to GAPDH, and compared with the mean for untreated samples used as the calibrator.
Analysis of TGFB1 promoter activity
Samples of DNA from patients with previously determined genotypes, −509CC and −509TT, were used for amplification of a 669-bp-long fragment consisting of the TGFB1 basal promoter and part of the silencer region containing position −509. The obtained PCR products were cloned into the PCR cloning vector pBluescriptT/A using restriction enzymes HindIII and NcoI (Fermentas). 27 The fragments were then recloned into luciferase expression vector PGL4.10 (Promega) using the same enzymes to obtain constructs PGL-TGFB1C and PGL-TGFB1T carrying the different C-509T variants.
The BEAS 2B cells were transfected by constructs, PGL-TGFB1C and PGL-TGFB1T, and cotransfected with Renilla luciferase expression vector (for normalization of luciferase activity) employing Lipofectamine 2000 (Invitrogen) according to the manufacturer's protocol. Five hours after transfection, cells were pretreated with TGFB1 (0.1 ng/mL) overnight. The next day, cells were treated with montelukast (100 μM) for 1 h. Fresh medium without montelukast was added to the control cells. The promoter activity was measured in 3 independent experiments using the Dual-Luciferase Reporter Assay System (Promega) on a GloMax 20/20 Luminometer (Promega).
Statistical analysis
The Statistical Package for Social Sciences 20.0 (SPSS, Inc.) was used for statistical analysis. Data are expressed as percentages and mean ± standard deviation for continuous variables and percentages for categorical variables. To test for normal distribution of the parameters, the one-sample Kolmogorov–Smirnov test was used. Differences between groups for categorical data were evaluated by χ 2 analysis, while for continuous data, the following were employed: independent sample t-test, independent samples Mann–Whitney U test, Kruskal–Wallis test, and related samples Wilcoxon signed-rank test. P values <0.05 were considered statistically significant.
Results
The study has included a group of 102 asthmatic children and adolescents (57% boys, aged 5–19 years, average 11.7 ± 3.5 years). One-third of the patients (33.3%) had partly controlled asthma, while 66.7% had good control of the disease.
All study subjects were genotyped for C-509T polymorphism in the promoter of the TGFB1 gene by direct sequencing. The allele −509C was present at a frequency of 52.9%, while frequency of the −509T allele was 47.1%. The distribution of genotypes was 30.4% for −509CC, 45.1% for −509CT, and 24.5% for −509TT. Monocytes and macrophages were observed with higher frequency in induced sputum of genotypes −509CC and −509CT than in induced sputum of genotype −509TT, but the differences were not statistically significant (7.6% and 7.8% versus 5.0%, P = 0.051 for monocytes; 16.3% and 16.5% versus 13.5%, P = 0.066 for macrophages).
Clinical characteristics, distribution of genotypes, and allele frequencies were compared between patients with different clinical phenotypes (Table 2). No significant differences were found in the allele distribution and genotype frequencies between mild and severe asthma. There were more sputum eosinophils in patients with severe asthma, but sputum cell profiles did not differ significantly between the groups (P = 0.089). There was a significant difference in control of asthma between patients with mild and severe asthma (P < 0.001).
Statistically significant.
FEV1, forced expiratory volume in 1 second; FVC, forced vital capacity; SD, standard deviation.
The impact of montelukast on TGFB1 gene expression was analyzed by qRT-PCR in sputum cells, leukocytes, and BEAS 2B cells. TGFB1 gene expression in response to montelukast was analyzed on a set of 22 samples of induced sputum and in 3 blood samples (used as the reference for gene expression in leukocytes). A decrease in gene expression was observed in treated sputum cells taken from 21 patients, while in 1 patient, an increase was found. Changes in mRNA expression between treated and untreated sputum cells were similar for each genotype (49.6% for −509CC, 31.8% for −509CT, and 58.9% for −509TT; P = 0.071) (Fig. 1). We found no statistically significant difference in TGFB1 gene expression between treated and untreated sputum cells in the dominant or the recessive genetic inheritance model (CC versus CT+TT, P = 0.704; CC+CT versus TT, P = 0.056, respectively).
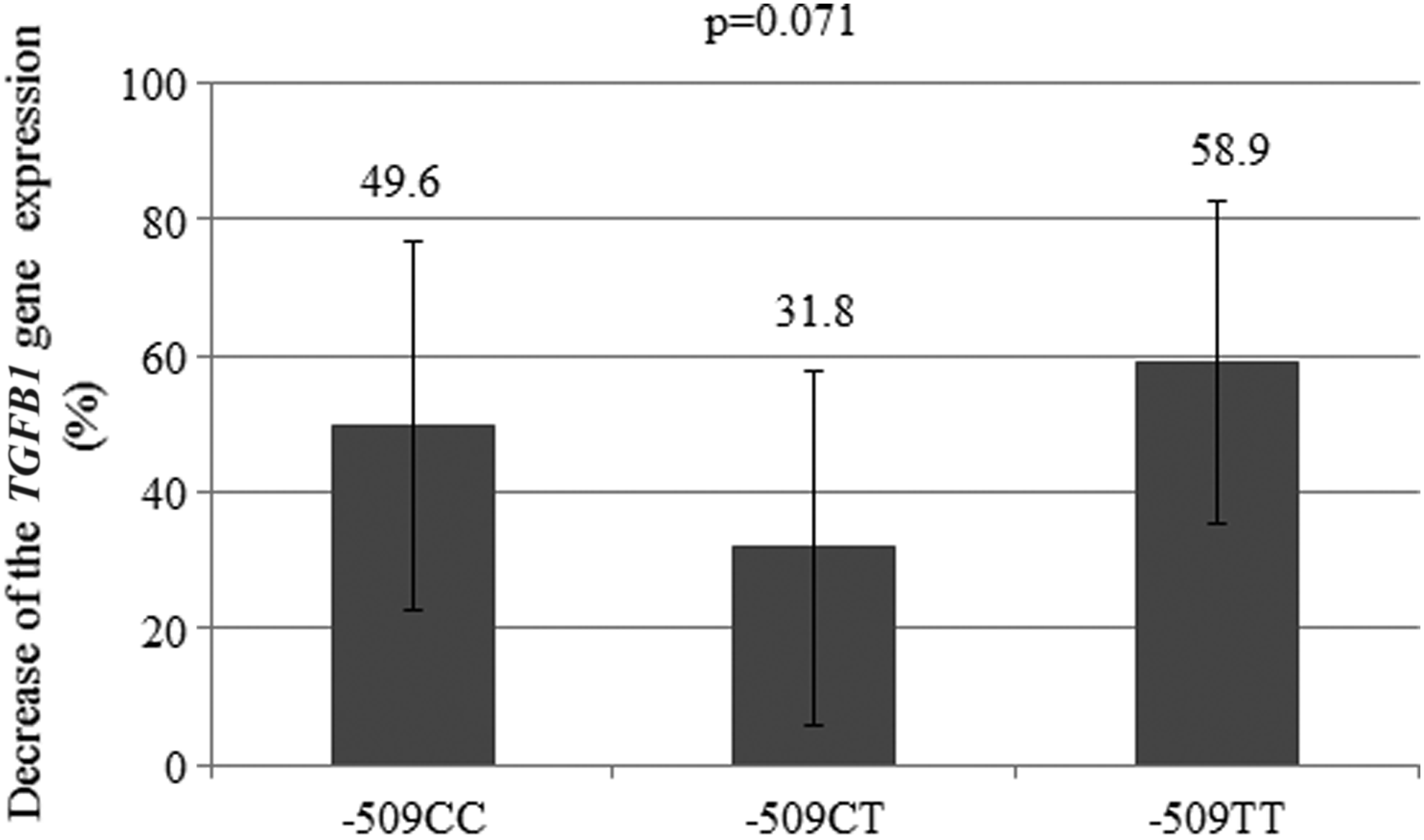
The decrease in TGFB1 gene expression measured by qRT-PCR in sputum cells after montelukast treatment of groups of patients with different genotypes. qRT-PCR, quantitative real-time polymerase chain reaction; TGFB1, transforming growth factor beta 1.
Blood leukocytes treated with montelukast showed a decrease in TGFB1 gene expression (30%–40%) in comparison with untreated leukocytes.
Treatment of BEAS 2B cells with montelukast also led to lower expression of endogenous TGFB1 by about 27% when compared with the untreated control. To analyze the effects of C-509T polymorphism and the influence of montelukast on TGFB1 gene promoter activity, BEAS 2B cells were transiently transfected with constructs PGL-TGFB1C and PGL-TGFB1T. In untreated cells, the presence of the −509T allele increased luciferase activity by about 75% in comparison with that for the −509C allele (P < 0.001). Treatment with montelukast raised promoter activity in cells transfected with either PGL-TGFB1C or PGL-TGFB1T in comparison with the respective control (48.3% for the −509C allele, P = 0.060; and 100.5% for the −509T allele, P = 0.062) (Fig. 2). Luciferase activity of the promoter with the −509T variant was increased after the treatment by 135% in comparison with the activity of the promoter containing the −509C variant (P = 0.035).

Luciferase activity of the reporter constructs carrying the TGFB1 gene promoter in BEAS 2B cells.
Discussion
In this study, we aimed to investigate the influence of the polymorphism C-509T in the TGFB1 gene promoter on response to montelukast. We used 2 different model systems, induced sputum cells taken from children with asthma (ex vivo) and a human bronchial epithelial cell line (in vitro).
Each subject in the study group was genotyped for C-509T and the distribution of genotypes and alleles was similar to that previously observed for adult asthmatics in Serbia. 28 We examined the correlation between C-509T and selected clinical parameters and found that the −509TT genotype might be associated with lower production of monocytes and macrophages in sputum, which was expected considering that the −509T allele is associated with greater production of TGFB1, so it may increase its inhibitory effect on these inflammatory cells. 29
A positive correlation between the −509TT genotype and asthma severity has been found in adults. 30 In our study, there were no statistically significant differences between patients with mild and severe asthma concerning genotype distribution. Nevertheless, the frequency of the −509T allele was somewhat higher in our patients with severe than in those with mild asthma (54.3% versus 41.8%; P = 0.076). Our finding is consistent with previous reports for Brazilian children and adolescents and adults from Poland and Japan.12,14,31 These divergent results may be due to disparate selection criteria, different sample sizes, and/or interactions of variant −509T with other genetic and environmental factors. However, asthma control did differ significantly between our groups (P ≤ 0.001). Almost all patients with mild asthma controlled the disease well (94.5%), whereas that was true for less than a third of the severe asthmatics (31.9%). Despite high cell variability in sputum, it was noticed that patients with severe asthma had more sputum eosinophils than those with mild disease. This finding supports the hypothesis that eosinophils have a crucial role in persistent airway inflammation and remodeling in severe asthma.10,11
The influence of C-509T polymorphism on the response to montelukast was investigated using ex vivo treated and untreated sputum cells from 22 available samples of induced sputum. Montelukast showed a tendency to reduce expression of the TGFB1 gene in sputum cells, more so for the −509TT genotype (58.9%) than for the −509CC and −509CT genotypes (49.6% and 31.8%, respectively), but the difference failed to reach statistical significance (P = 0.071). Although, the patients were pretreated with salbutamol before sputum collection, the use of cells originating from the same sputum sample allowed us to avoid the potential influence of external factors on TGFB1 gene expression, including salbutamol. Moreover, an earlier study showed that beta 2 agonists do not alter TGFB1 plasma levels after several days of treatment. 32 Therefore, it could be suggested that carriers of genotype −509TT might respond to montelukast by lower production of TGFB1. This finding should be confirmed in a larger cohort of patients and especially in asthmatics on montelukast monotherapy.
The response to montelukast was also investigated using BEAS 2B cells. Expression of the TGFB1 gene in these cells was reduced by about 27% after treatment with montelukast in comparison with the untreated control. The smaller reduction in expression of TGFB1 in this case may be due to differences in the model systems. Since eosinophils are the main source of CysLTs in sputum, the significant decrease in expression of TGFB1 in sputum cells after treatment with montelukast might result from their effects on receptors for leukotrienes. The in vitro analysis lacked eosinophils and CysLTs. It is still unclear whether montelukast may affect TGFB1 expression through mechanisms independent of receptors for leukotrienes. In mice, montelukast was found to attenuate TGFB1 gene expression by reducing SMAD3 expression, which is involved in canonical TGFB1 signaling. 33 On the other hand, CysLTs were shown to alter TGFB1 expression through a p38 mitogen-activated protein kinase pathway. 34 It appears that montelukast might regulate TGFB1 transcription activity through pathways that are either SMAD dependent or independent, but the exact mechanism remains unknown.
The impact of C-509T on promoter function was investigated by comparing luciferase activity of constructs containing either the C or T variant at position −509 transiently transfected into BEAS 2B cells. We found that the −509T allele increased basal promoter activity by about 75% compared with the promoter containing the −509C allele. The detected difference in the promoter activity between these 2 alleles was double that previously reported. 13 Since BEAS 2B cells were pretreated overnight with TGFB1, which was necessary to obtain detectable activity in the luciferase assay, this finding might be due to stimulatory effects on TGFB1 gene expression. In contrast to the expression of endogenous TGFB1 in sputum and BEAS 2B cells, treatment with montelukast showed a tendency to increase luciferase activity of both promoters with a more intensive response from the promoter containing the T allele (48.3% for the C allele and 100.5% for the T allele).
The increase in promoter activity measured by the luciferase assay, as opposed to the decrease of gene expression measured by qRT-PCR, appears to be a contradictory result. For that reason, we tested the influence of montelukast on leukocytes from peripheral blood of asthmatic patients. Our results showed that montelukast leads to increased promoter activity, but a decreased level of TGFB1 mRNA. However, it should be highlighted that the luciferase assay included analysis of only the portion of the TGFB1 5′ regulatory region that encompasses the basal promoter and part of the silencer region containing the C-509T polymorphism. The effects of other regulatory sequences in the TGFB1 promoter have not been investigated in this context. Post-transcriptional regulatory mechanisms could also contribute to the different effects of montelukast registered for promoter activity and gene expression levels. To clarify the effect of montelukast on TGFB1 gene regulation and the influence of the C-509T polymorphism, all the above mentioned factors should be taken into consideration in future studies.
Conclusions
The main finding of this study is that the base change C to T at position −509 in the TGFB1 gene promoter might be associated with response to montelukast. This finding is worth further investigation and may prove valuable if polymorphism C-509T in the TGFB1 gene promoter is confirmed as a predictor of response to montelukast in asthmatics. More comprehensive studies are necessary, especially in the clinical setting with a larger group of patients. The response to montelukast should be more closely followed, especially in patients on montelukast monotherapy. It would be of potential importance for clinical management of asthma to clarify the influence of the C-509T polymorphism on the response to treatment with montelukast.
Footnotes
Acknowledgments
For the BEAS 2B cell line used for this research, the authors owe gratitude to Dr Peter Lackie (School of Medicine, University of Southampton, Southampton, United Kingdom). The Department of Pharmaceutical Chemistry, Faculty of Pharmacy, University of Belgrade, Belgrade, Serbia, kindly provided the montelukast. The authors wish to thank Ana Kojic (Laboratory for Molecular Biology, Institute of Molecular Genetics and Genetic Engineering) for help with cloning. The research was supported by a grant 173008 of the Ministry of Education, Science and Technological Development of the Republic of Serbia.
Author Disclosure Statement
No competing financial interests exist.