
Abstract
Objective:
High-mobility group box 1 protein (HMGB1) serves as an inflammatory mediator and has been implicated in the pathogenesis of asthma. Extracellular HMGB1 transduces cellular signals through the plasma membrane receptor for advanced glycation end products (RAGE). The HMGB1/RAGE axis has proven to be a pivotal factor in the progression of tumor growth and inflammation. In this study, we sought to determine whether HMGB1 signaling through RAGE occurs in asthma.
Methods:
C57BL/6 mice were randomly divided into 4 groups, asthma mice; nonasthma control mice; glycyrrhizic acid ammonium salt from glycyrrhiza root (GL) therapy mice; and high-affinity RAGE-specific inhibitor (FPS-ZM1) therapy mice. Asthmatic mice were sensitized and challenged with ovalbumin (OVA), and control mice were treated with saline. These 2 groups were divided into 4 subgroups, which were sacrificed at days 21, 28, 35, and 42. GL and FPS-ZM1 therapy mice were first sensitized with OVA at days 0 and 7 and then injected intraperitoneally with GL (20 mg/kg) or FPS-ZM1 (5 mg/kg) 30 min before each challenge. The gene expressions were measured by RT-PCR. Hematoxylin-eosin staining was used to observe mouse lung structural changes. Immunofluorescence immunohistochemistry was used to detect HMGB1 and RAGE proteins in lung tissue. The expressions of HMGB1, toll-like receptor 2 (TLR2), TLR4, and RAGE were significantly increased in asthma mice (P < 0.05). In addition, HMGB1 and RAGE displayed similar trends over time; both reached a peak at day 28 in asthma mice and then gradually declined. Infiltration of inflammatory cells into lung tissue, L-4, and IL-13 expression in asthma mice was suppressed after treatment with either the HMGB1 or the RAGE inhibitor. The effect of FPS-ZM1 was weaker than that of the GL therapy.
Conclusion:
In conclusion, a part of the HMGB1 contribution to the pathogenesis of asthma was mediated through binding to RAGE.
Introduction
A
HMGB1, first identified in 1973, is described as a nuclear DNA-binding protein that helps stabilize nucleic acid structures and regulate transcription and gene expression. 3 HMGB1 can be passively secreted by necrotic cells, actively secreted by many types of innate immune cells such as monocytes and macrophages, and plays important roles in the pathogenesis of sepsis, arthritis, pneumonia, endotoxemia, and other inflammatory diseases. 4 Currently, research on HMGB1 and its receptors is active. HMGB1 may bind to TLR2, TLR4, or RAGE to activate intracellular signaling pathways causing inflammatory responses. TLR2 and TLR4, belonging to the toll-like receptor family, were first discovered in 1977 and have the family characteristic extracellular region of leucine-rich repeat motifs. The HMGB1/TLR2 axis was reported to be responsible for myocardial ischemia/reperfusion injury. 5 Wang et al. showed that downregulated expression of HMGB1 and TLR4 reduced the pain caused by chronic pancreatitis. 6 HMGB1 binding to the RAGE receptor may contribute to the pathogenesis of diabetic retinopathy, accelerating rat retinal cell apoptosis. 7 However, the identity of the receptor involved in HMGB1-mediated asthma is still controversial. Therefore, in this study, we attempted to determine whether HMGB1 signaling through RAGE contributes to the inflammatory response in asthma.
Glycyrrhiza root (GL) was first used in the treatment of hepatitis. 8 Recently, GL was shown to reduce inflammation of lung tissue and AHR and to inhibit the expression of IL-4 and IgE in an asthma model.9,10 Several studies have shown that GL, as an HMGB1 inhibitor, may combine directly with HMGB1, inhibit its activity, thereby suppressing inflammatory responses induced by HMGB1.11–13
FPS-ZM1 is a high-affinity, RAGE-specific inhibitor of ligand binding to the V domain of RAGE. FPS-ZM1 has been characterized as nontoxic, efficient, and tissue penetrating. Reports have shown that FPS-ZM1 reduces vessel density, VEGF expression, and blood–brain barrier damage after intracranial hemorrhage and amyloid beta-mediated brain disorder in Alzheimer's disease by inhibiting RAGE.14–16
Methods
Experimental animals and treatments
Six-to-eight-week-old, male C57BL/6 (SLACCAS, Shanghai, China) mice (SLACCAS) were bred and housed in a pathogen free, authorized facility, where the temperature was maintained at 18°C–22°C, humidity at 30%–50%, with a 12 h day and night cycle. The mice were fed with laboratory water and fodder. All animal studies were approved and complied with the guidance of the Institutional Animal Care and Use Committee of Soochow University. The 24 asthma mice were sensitized by intraperitoneal injection of 100 μg/0.5 mL ovalbumin (OVA) (Sigma, St. Louis, MO) with 400 μg Al(OH)3 (Sigma) as adjuvant on days 0 and 7 and were then challenged with 1% OVA aerosol for 30 min on days 14–20. All the 24 control mice were similarly treated, with saline replacing OVA in both the sensitization and challenge processes. Six mice were randomly sacrificed on days 21, 28, 35, and 42 in the asthma and control groups. The mice in the GL or FPS-ZM1 groups were sensitized with OVA on days 0 and 7, in a similar manner to the other groups, and then received intraperitoneal injections of 20 mg/kg GL (Sigma) or 5 mg/kg FPS-ZM1 (Calbiochem, LaJolla, CA), respectively, on days 14–20, 30 min before each OVA challenge. GL and FPS-ZM1 therapy mice (n = 6 each group) were all sacrificed on day 21. The conduct of this project fully satisfied the animal welfare requirements and all other criteria of the Medical Ethics Committee of the Children's Hospital of Soochow University, which approved the study.
Real-time PCR
Lung tissue RNA was extracted in TRIzol (Invitrogen, Carlsbad, CA) following the manufacturer's protocol. In brief, 1 μg of total RNA was used for reverse transcription according to the manufacturer's protocol (Takara RNA PCR kit, Shiga, Japan). The resulting cDNA was analyzed and amplified using the real-time PCR (ABI PRISM7500). The reaction solution, consisting of 2 μL of cDNA, 10 μL of SYBR Green I Master (Roche, Mannheim, Germany), 1 μL of forward primer and 1 μL of reverse primer, was prepared in a total volume of 20 μL. The solution was initially denatured at 95°C for 10 min and then subjected to 30 thermal cycles. Each cycle included the steps of denaturation at 95°C for 15 s, annealing at 60°C for 15 s, and extension at 72°C for 30 s. All samples were analyzed in triplicate. The real-time quantification of transcription was measured by the comparative threshold cycle number (Ct). The sequences of all primers are shown in Table 1. β-Actin served as internal control.
Histology of the lung: immunofluorescence and immunohistochemistry
Lung tissue was soaked in 10% neutral formalin until lobe separation, then embedded in paraffin, cut into 3 μm thick slices, and stained with hematoxylin and eosin (H&E).
Tissue sections on slides were deparaffinized, rehydrated, and subjected to antigen retrieval by heating. Tissue sections were incubated in 3% H2O2 for 10 min and in bovine serum albumin for 1 h. The primary antibody to HMG-1 (sc-12523, 1:50; Santa Cruz Biotechnology, San Francisco) or RAGE (BA1789, 1:50; BOSTER, Wuhan, China) was then added and incubated overnight at 4°C. On the following day, the secondary antibody (Donkey Anti-Goat IgG, P1073, 1:1,000; Beyotime, Shanghai, China) or Goat Anti-Mouse/Rabbit IgG, GK5005; Gene Tech, Shanghai, China was added on the slides and incubated for 30–60 min at room temperature. Slides incubated with anti-HMG-1 were incubated with DAPI for 10 min. Tissue sections incubated with anti-RAGE were stained for 40 s with H&E and then dehydrated and sealed. Finally, the samples were observed and photographed with an Olympus microscope.
Statistical analysis
All data are shown as the mean ± standard deviation. Data analysis of multiple comparisons was carried out by ANOVA. A t-test was used to analyze differences between groups. Statistical significance was accepted for P < 0.05. All statistical analyses were performed using SPSS 17.0 statistical software.
Results
Histological examination of lungs
Lung tissue was collected 24 h after the last challenge with 1% OVA aerosol at day 20 for H&E staining. Compared with the control mice, there was an obvious increase in inflammatory cell infiltration around the bronchioles and alveoli characteristic of lymphocytes and eosinophils. Furthermore, increased goblet cell hyperplasia, airway epithelial damage, and interstitial edema were found in OVA challenged mice. Following treatment with GL or FPS-ZM1, asthmatic mice showed a marked reduction in inflammatory cell infiltration, airway epithelial damage, and interstitial edema. Furthermore, mice treated with GL showed significantly greater improvement than mice treated with FPS-ZM1 (Fig. 1).
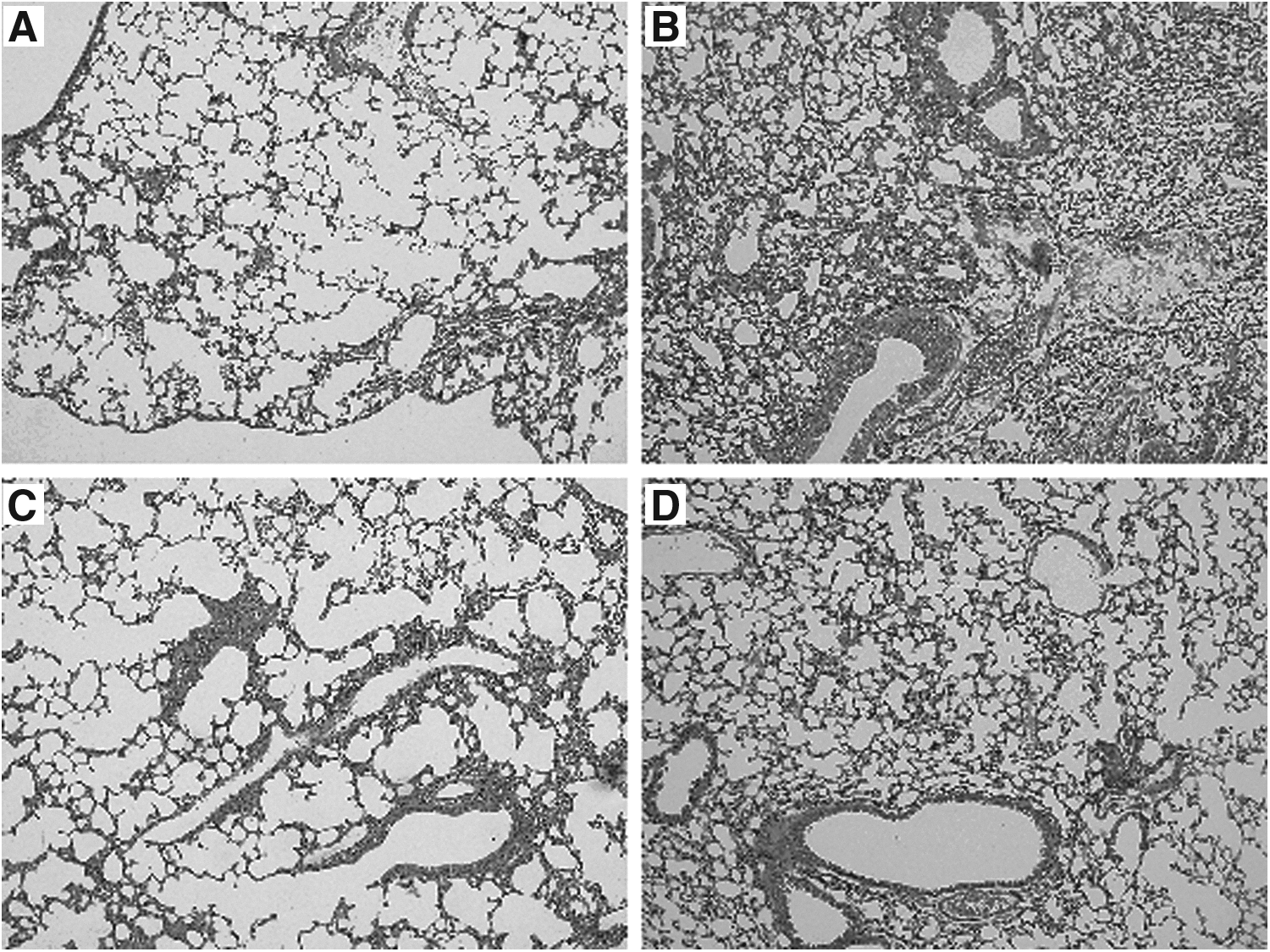
Histological sections of lung tissue were essayed using H&E staining 24 h after the last challenge (magnification, × 200).
HMGB1/RAGE/TLR2/TLR4 dynamic changes in gene expression levels
HMGB1, TLR2, TLR4, and RAGE gene expression differed significantly depending on treatment and time (P < 0.05). HMGB1 and RAGE gene expression reached a peak at day 28 in asthmatic mice and then gradually decreased. Compared with the control mice, HMGB1 was significantly higher only at day 28 in asthmatic mice, while RAGE was still elevated compared with controls on day 42. TLR2 and TLR4 levels gradually decreased over time and the increase of the former was still significant on the 35th day. The TLR4 showed no significant difference between asthma and control mice after the day 28. In addition, RAGE expression was significantly higher than TLR2 and TLR4, 12- and 5-fold, respectively, at day 28, as shown in Fig. 2.

Gene expression of HMGB1, RAGE, TLR2, and TLR4 in asthmatic and control mice at days 21, 28, 35, and 42. All time points showed significant (P < 0.05) effects with different treatments compared with controls. *Significance higher than controls (P < 0.05). Graphic depicts the mean ± SEM.
HMGB1 expression in lung tissue sections
Immunofluorescence microscopy was used to examine the location and expression of HMGB1 in the lung. As shown in Fig. 3, the cytoplasm was significantly stained, suggesting that HMGB1 was mainly localized in the cytoplasm. In addition, the lung tissue from asthmatic mice exhibited abundant HMGB1 compared with the lung tissue from mice treated with GL, indicating that GL inhibited the expression of HMGB1.

Immunofluorescence staining of HMG-1 of the asthma mice/asthma mice treated with HMGB1 inhibitor (GL). The GL-treated mice had less HMGB1 proteins compared with the asthma mice. GL, glycyrrhiza root.
RAGE expression in lung tissue sections
After treatment with FPS-ZM1, RAGE protein was detected in lung tissue by the immunohistochemistry method. As predicted, RAGE was significantly reduced in the lungs of mice administered FPS-ZM1 compared with asthmatic mice, by ∼50% (P < 0.05). Moreover, RAGE was mainly distributed in alveolar epithelial cells and expressed in the cell membrane, as shown in Fig. 4.

Immunohistochemistry staining of RAGE of the asthma mice/asthma mice treated with RAGE inhibitor (FPS-ZM1). The arrow showed the positive staining cell. The FPS-ZM1-treated mice had less RAGE proteins compared with the asthma mice (*P < 0.05).
Effect of inhibitors on cytokines in the lung
Following administration of GL or FPS-ZM1, the levels of IL-4, IL-5, and IL-13 in lung tissue decreased; however, only IL-4 and IL-13 decreased significantly (P < 0.05). GL therefore downregulated the expression of IL-4 and IL-13 compared with asthmatic mice, by ∼1.5% and 16%, respectively. The gene expression of the cytokines IL-4 and IL-13 was also inhibited after treatment with FPS-ZM1 by ∼26% and 31%, respectively. Further analysis showed that the expression of IL-4 and IL-13 in FPS-ZM1 therapy mice was higher than those of GL therapy mice, by ∼17- and 2-fold, respectively, as shown in Table 2.
1, the asthma mice and the GL therapy mice; 2, the asthma mice and the FPS-ZM1 therapy mice; GL, glycyrrhizic acid ammonium salt from glycyrrhiza root; IL, interleukin; OVA, ovalbumin.
Discussion
In this study, we found that HMGB1 plays an important role in the pathogenesis of asthma. First, the level of HMGB1 was significantly increased in the lung tissue in asthmatic mice compared with control mice, consistent with previous studies. 17 Second, GL significantly reduced the infiltration of inflammatory cells in lung tissue and the expression of inflammatory cytokines IL-4 and IL-13, but not IL-5.
In this article, the change in gene expression of IL-5 was not significantly different between asthmatic mice and GL therapy mice. We speculated that the expression of IL-5 in mice did not increase after sensitization and challenge by OVA. A few studies have shown similar results. Oh JW found that the concentration of IL-5 in sputum from children with asthma was not significantly increased compared with controls. 18 In addition, IL-5, which is a typical TH2 type cytokine, produced by CD4+ T cells that contribute to the production, mobilization, activation, recruitment, proliferation, and survival of eosinophils and inhibit apoptosis,19,20 may be specific to eosinophil-mediated inflammation. Zhou et al. noted that the level of HMGB1 in lung tissue was positively correlated with neutrophils, 21 and therefore the HMGB1 inhibitor may have no effect on eosinophil associated inflammation.
HMGB1 binding to cell surface receptors RAGE, TLR2, and TLR4 triggers phosphorylation of MAPK (including p38, JNK1/2, ERK1/2), nuclear transformation of nuclear transcription factor NF-κB and live protein 1, and increased expression of inflammatory cytokines such as IL-1β, TNF-α, and IL-8, initiating inflammation or leading to its reduction.22–24 Our findings suggest that TLR2, TLR4, and RAGE expression was increased in asthmatic mice. Previous studies have indicated that TLR2, TLR4, and RAGE modified antigen-induced airway inflammation responses in an experimental asthma model.25–27 However, in our study, only RAGE displayed a similar trend to HMGB1. Their gene expressions peaked at day 28, and RAGE expression was significantly higher than TLR2 and TLR4 at day 28. In addition, the RAGE-specific inhibitor, FPS-ZM1, inhibited inflammation around the lung tissue and decreased the expression of IL-4 and IL-13; however, the effect was weaker than GL. Therefore, we propose that HMGB1 contributed to the pathogenesis of asthma, at least, in part, via the RAGE receptor.
RAGE, first determined to be a potential mediator of diabetes, 28 is a multiligand transmembrane receptor, part of the immunoglobulin superfamily, and binds a variety of ligands, including HMGB1, S100 proteins, amyloid β, CD11b, and heparin. Van Zoelen found that RAGE, using I-HMGB1 ligand-binding analysis, was a major receptor for HMGB1. 29 Studies on the HMGB1/RAGE axis previously reported have indicated its involvement in tumor development, cholesteatoma, and the renal fibrosis of diabetic nephropathy30–32 ; however, its role in asthma is still controversial. Studies have shown that levels of HMGB1 and RAGE were increased in asthma patients' sputum, indicating that HMGB1 participated in asthmatic airway inflammation through the RAGE receptor.21,33 Interestingly, Fu et al. found that the expression of HMGB1 and RAGE was not significantly increased in lung tissue sensitized and challenged by OVA. 34 Lee et al. argued that TLR2 and TLR4, but not RAGE, were involved in the pathogenesis of asthma mediated by HMGB1. 17 The mice in the above-mentioned studies were sacrificed within 24 h after the last challenge and the expression of HMGB1 and RAGE analyzed. Analysis of our data revealed no significant difference in RAGE gene expression between asthma and control mice at day 21 within 24 h after the last challenge, consistent with previous results. However, 1 week later, the expression of RAGE began to significantly increase in asthmatic mice compared with the control mice, in a similar manner to HMGB1, and, therefore, RAGE appears to be involved in HMGB1-mediated inflammation in asthma.
Conclusion
In this study, inflammatory cytokine and inflammatory cell infiltration were significantly reduced in asthma mice after treatment with GL or FPS-ZM1. However, the effect of FPS-ZM1 therapy was weaker than GL therapy. Therefore, we conclude that HMGB1 may partially act via RAGE to contribute to the pathogenesis of asthma in a mouse model and the HMGB1/RAGE axis may be a new target for the treatment of asthma.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (Grant No. 81570016), the Program of Science and Technology Plan of Suzhou (Grant No. SS201535) and the Jiangsu Provincial Key Laboratory of Radiation Medicine and Protection.
Author Disclosure Statement
No competing financial interests exist.