
Abstract
Background:
Bacteriophages are a promising biotechnological against bacterial pathogens. Currently, phage research is garnering interest in sub-Saharan countries as bacterial resistance to antibiotics becomes widespread. They are sought in all environments as they offer the possibility of a sustainable alternative to antibiotics.
Materials and Methods:
Altogether 30 water samples from urban sewage and environmental water were screened for the presence of bacteriophages able to infect Escherichia coli and Enterobacter cloacae. Their genomic diversity was determined by random amplification of polymorphic DNA (RAPD)-PCR fingerprinting.
Results:
We isolated 35 phages including 9 polyvalent phages that infect simultaneously E. coli and E. cloacae. This study allowed first isolation of E. cloacae-specific phages in Côte d'Ivoire. All phages were distinct based on their RAPD band patterns.
Conclusions:
Sewage systems of Yopougon and the environmental water of Ebrié lagoon were a rich source of phages. The phage collection could be useful for phage application in Côte d'Ivoire.
Introduction
Bacteriophages, the viruses of bacteria, are present in the same ecological niche as their hosts. They are the most abundant biological entities on Earth. Consequently, phages have an impact on bacterial communities and local and global ecological processes.1,2 Phages have been observed or isolated in a wide variety of environments such as human or animal gastrointestinal tract, sewage, environmental water, soil, and sediment.3–6 The study of these bacterial viruses has led to several biotechnological applications in European and American countries. For example, lytic phages have been used for preventing or treating bacterial disease in humans and animals7–10 and for detecting bacteria.11,12 They were applied also in food processing13–17 and for decontaminating of surfaces.18–20
Similar to all viruses, phages are very species-specific regarding their hosts and are often specific to only a few strains within that species 21 ; however, polyvalent phages are detected among Enterobacteria.22–24 These polyvalent phages can infect strains from either different genera or species. 12 Polyvalent phage therapy constitutes a promising biocontrol technology as shown by a study carried out to control antibiotic-resistant bacteria in soil–plant system. 25 Few broad host range phages could be more useful than a larger number of narrow host range phages.26,27
Because of the renewed interest in lytic phages as biocontrol agents due to the multidrug resistance of bacteria, new phages are sought after as they offer the possibility of a sustainable alternative to antibiotics. Among the bacteria with multidrug resistance, Enterobacter cloacae are known to be a major opportunistic and multidrug-resistant pathogen for humans during the past three decades. 28 E. cloacae strains are ubiquitous and found in terrestrial and aquatic ecosystems (environmental water, wastewater, soil, and food), in the environment but also in hospitals. Indeed, they are responsible for nosocomial infections. As early as 1986, E. cloacae were the most frequently isolated bacterium in hospitals in Côte d'Ivoire. 29 Nowadays, E. cloacae is the third Enterobacteria producing extended-spectrum beta-lactamase (ESBL) isolated from human clinical samples. 30 At the top of this ranking, Escherichia coli was involved in severe diarrheal diseases especially in young children. 31 Studies in Africa have revealed the disturbing emergence of antimicrobial resistance and high rates of asymptomatic carriage of diarrheal E. coli. 32 E. coli resistance to third-generation cephalosporins is as high as 87% and 98% for fluoroquinolones. 33 The rate of subjects carrying ESBLs varies from 10% to 50% and is >60% concerning E. coli. 34
To isolate new phages effective against various strains, we investigated E. coli and E. cloacae lytic phages in samples of sewage and environmental water in Côte d'Ivoire.
Materials and Methods
Culture media, bacterial strains, and growth conditions
Single-strength Lysogeny Broth (LB) (10 g/L NaCl, 10 g/L tryptone, and 5 g/L yeast extract) was used for bacterial culture. Double-strength LB broth supplemented with 2 mM CaCl2 was used for phage enrichment. LB 1 × supplemented with 1.5% agar was used as a solid growth medium. Soft agar for spot test assays was LB supplemented with 0.6% agar. Saline magnesium (SM) buffer (50 mM Tris-HCl pH7.5, 100 mM NaCl, and 8.1 mM MgSO4) was used to store purified phages at 4°C.
Three bacterial strains were used for phage isolation. Escherichia coli B and C reference strains were provided by the Félix d'Hérelle Reference Center for Bacterial Viruses, Laval University, Canada. E. cloacae was isolated in wastewater sample in 2017, and was provided by the Plateforme de Biologie moléculaire of Institut Pasteur de Côte d'Ivoire. Bacteriophage T4 (HER27) was used as a control for spot test assay. Bacterial strains were grown overnight and stored in 200-μL aliquots in medium with 30% glycerol at −20°C. A hundred microliters from stored bacteria were added to 3 mL of medium and grown overnight at 37°C before use for phage isolation and characterization.
Samples collection
The sampling was carried out periodically from February 2018 to November 2019. Three samples were collected from each site giving a total of 15 samples from urban sewage (Sites 1–5) and 15 samples from Ebrié lagoon (Sites 6–10). The open-air drainage system in Yopougon received domestic, industrial, and hospital wastewater from the municipality. The content of those collectors and those from the other municipalities of Abidjan are discharged directly in Ebrié lagoon without treatment (Fig. 1).

Sampling sites. The map of Abidjan showing different sampling sites.
Enrichment of bacteriophage
Filtered water samples were enriched for bacteria-specific phage isolation as described previously with some modifications. 35 In brief, 10 mL of filtered water were added to 10 mL of double-strength LB broth containing 2 mM CaCl2 with 100 μL of the enrichment bacterial strain. The mixture was incubated at 37°C, 48 h. Then, the culture was centrifuged at 12,000 g, 5 min, and the supernatant, containing phages, filtered through a 0.45-μm membrane filter.
Phage isolation, enumeration, and purification
The double-layer agar method was used to obtain a single isolated plaque. In brief, serial decimal dilution up to 10−8 of filtered lysate was carried out. Then, 100 μL of overnight bacterial culture (18–24 h) was mixed with 0.6% soft agar and poured on a 1.5% bottom agar plate. When the top agar layer was set, 10 μL of serial dilutions of filtered lysate were spotted onto the overlay, and plates were incubated overnight at 37°C and observed for countable isolated plaques.
Plaque purification was carried out by stabbing a sterile truncated tip through a single well-isolated plaque that was immediately added to 500 μL of SM buffer, mixed and refrigerated for 1 h at 4°C. After incubation, the mixture was centrifuged and 300 μL of supernatant was filtered and transferred to a new tube. The once purified phage was diluted again and spotted on an agar lawn. Phage plaques were purified three times consecutively to ensure phage purity. 3
Phage stocks
Phage stocks were prepared following the protocol of Raya and Hébert with some modifications. 36 A 100 μL of an overnight host strain culture was added to 10 mL of 1 × LB broth supplemented with CaCl2 to a final concentration of 10 mM. The mixture was incubated at 37°C, 1 h. Then, 100 μL of purified phage was added to the mixture and incubated for 8 h and cell lysis was regularly monitored by visually comparing phage-containing tubes to the control tube containing host strain. The tubes were centrifuged at 10,000 g for 10 min, 4°C. The supernatants were filtered into new sterile tubes. The filtered solutions were titrated and stored at 4°C.
Host range determination
The spot assay was used to assess the bactericide ability of isolated phages to form clear zones on diverse bacterial strains. 18 This was repeated three times for each phage. In brief, the potential host strain was grown overnight and 100-μL aliquots were mixed with soft agar and plated onto LB bottom agar plates. Once set, 10 μL of diluted phage lysate was dropped on LB lawns of agar containing a potential host strain. Plates were incubated overnight at 37°C. Phage activity was indicated by the appearance of a clearing zone on the bacterial lawn at the site of phage application, and by the development of individual plaques as the phage lysate is diluted.
Phage DNA isolation
Phage DNA was extracted using DNeasy Blood and Tissue Kit (Qiagen, Hilden, Germany) as described previously 37 from 450 μL of phage stocks (1010–1014 pfu/mL).
Genomic fingerprinting by random amplification of polymorphic DNA analysis
Random amplification of polymorphic DNA (RAPD) was carried out according to a modification of the method described previously. 38 P1 (5′-CCGCAGCCAA-3′) and P2 (5′-AACGGGCAGA-3′) were used. 39 PCR mixture (50 μL) consisted of 1 μL of phage DNA, 5.0 μL 5 × Green Reaction Buffer, 5.0 μL 5 × Green Reaction Buffer, 3.0 μL MgCl2 (25 mM), 1.0 μL dNTP's (25 mM) (Promega, Madison, WI), 1 μL primer, 0.5 μL GoTaq G2 Flexi DNA polymerase (5 U/μL) (Promega) and 29.5 μL DNase/RNase free water (Promega). Reactions were performed at 95°C for 5 min followed by 45 cycles of consecutive primers annealing (26°C and 31°C for 3 min), extension (72°C for 2 min), denaturation (94°C for 1 min), and final extension (72°C for 10 min).
Processing, comparison of RAPD patterns
DNA band patterns were obtained after gel electrophoresis on 2% agarose gel of the RAPD-PCR reaction products (15 μL). Gels were run for 120 min at 80 V using a 1-kb DNA ladder (Promega) as a molecular weight marker. The gels images were recorded using the Gel Doc EZ Gel Documentation System (BioRad). The RAPD profile was analyzed using the software GelJ. 40 The similarity matrix was calculated based on the Pearson correlation coefficient, and its corresponding dendrogram was deduced using the complete linkage clustering method.
Results
Phage collection
In this study, altogether 30 water samples were collected from urban sewage and Ebrié lagoon and screened for the presence of E. coli and E. cloacae-specific phages. A collection of 35 phages has been obtained from both biotopes. Phages isolated from urban sewage and Ebrié lagoon were represented, respectively, 54.29% and 45.71% of the collection. Twelve phages were isolated with Escherichia coli B, 10 with Escherichia coli C, and 13 phages with E. cloacae (Table 1). Coliphages were found in almost all sites, except in samples from Site 2.
Summary of Phage Isolation from Water Samples from Yopougon Urban Sewage and Ebrié Lagoon
The distribution of enriched phage titer regarding the sampling collection site revealed that samples from Ebrié lagoon produced under the same enrichment conditions, higher phage titer than samples from urban sewage (p = 0.000246) (Fig. 2). Indeed, phage titer obtained with Ebrie lagoon enrichment varied from 106 to 1011 pfu/mL, against 104 to 108 pfu/mL for urban sewage.
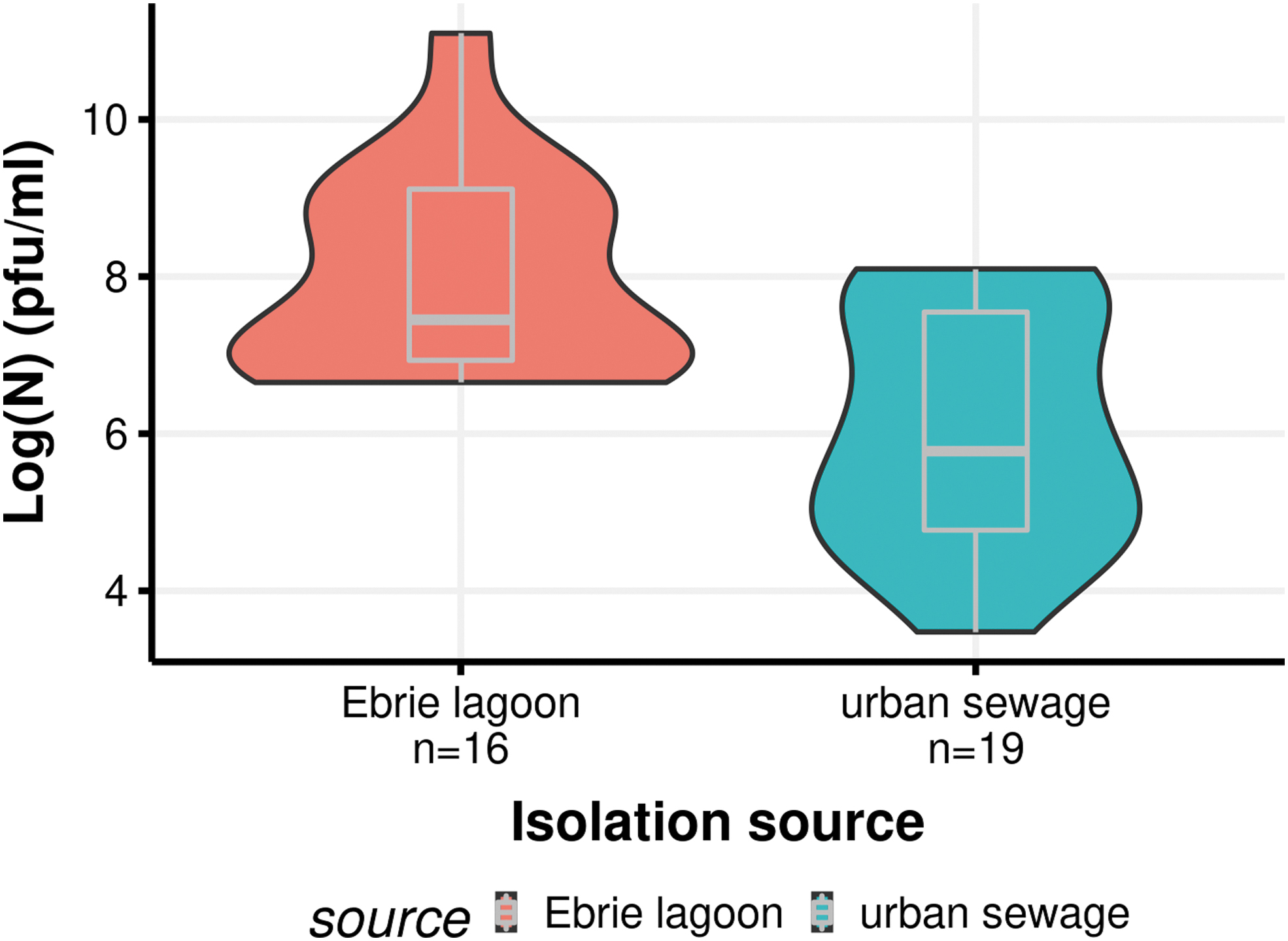
Distribution of phage titer (log pfu/mL) regarding isolation source.
Host range spectrum
The host range of a phage is defined as the ability of the phage to infect bacterial strains other than its enrichment host strain. 3 Strains were only considered susceptible if discrete plaques could be observed as the sample was diluted, indicating phage replication.
Four of six phages isolated with Escherichia coli C in urban sewage were capable to infect Escherichia coli B. In contrast, only EbM3 has shown lytic activity on Escherichia coli C (Fig. 3). Phages infecting Escherichia coli B isolated in Ebrié lagoon were specific to the isolation strain. One-fourth of the collection produced individual plaques simultaneously on E. coli and E. cloacae lawns. All coliphages issued from Ebrié lagoon have not shown lytic activity on the E. cloacae lawn. On the contrary, four coliphages isolated in urban sewage were able to produce cell lysis on the E. cloacae lawn. E. cloacae’ phages isolated in urban sewage were mainly specific except EnB2 and EnM2. Among the collection, 16 phages were able to infect strains other than their isolation strain, and EnBor1, EnCar1, and EbB1 infected the three studied strains (Tables 2 and 3).
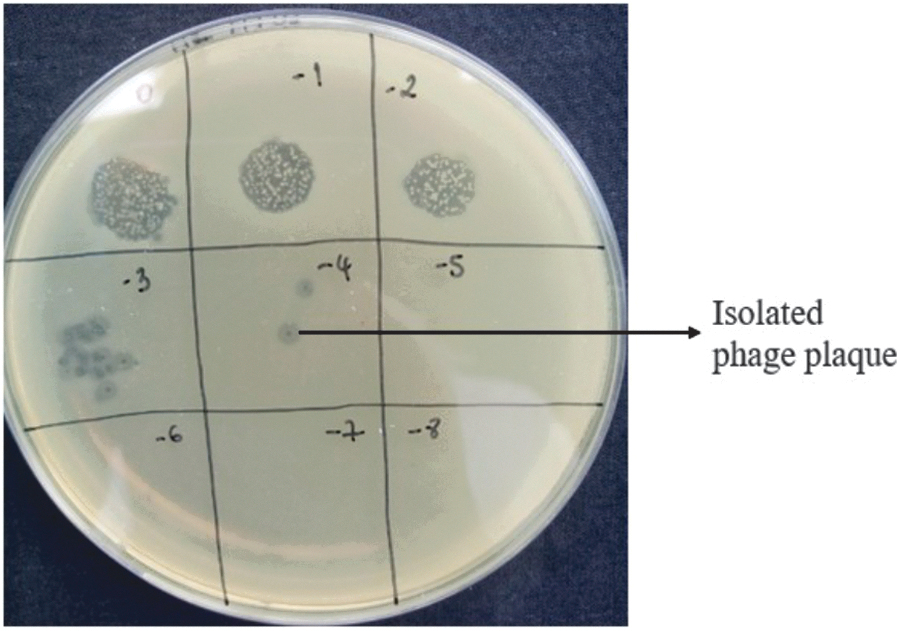
Phage EbM3 isolated with Escherichia coli B shown lytic activity on Escherichia coli C lawn. (0) = 109 pfu/mL; (−1) = 108 pfu/mL; (−2) = 107 pfu/mL; (−3) = 106 pfu/mL; (−4) = 105 pfu/mL; (−5) = 104 pfu/mL; (−6) = 103 pfu/mL; (−7) = 102 pfu/mL; (−8) = 10 pfu/mL.
Host Range of Phages Isolated in Yopougon Urban Sewage
(+) Distinct plaques seen; (−) No plaques observed.
Host Range of Phage Isolated in Ebrié Lagoon
(+) Distinct plaques seen; (−) No plaques observed.
Phage genetic diversity analysis
RAPD-PCR on phage DNA is a suitable approach to quickly assess the genetic diversity among newly isolated bacteriophages. Two different primers were used, out of which only P2 (5′-AACGGGCAGA-3′) provided bands with all the phages. The genomic fingerprints from the phages were all distinct and have shown different numbers of bands, fragment size, and intensity as shown for phages infecting Escherichia coli B in Figure 4.
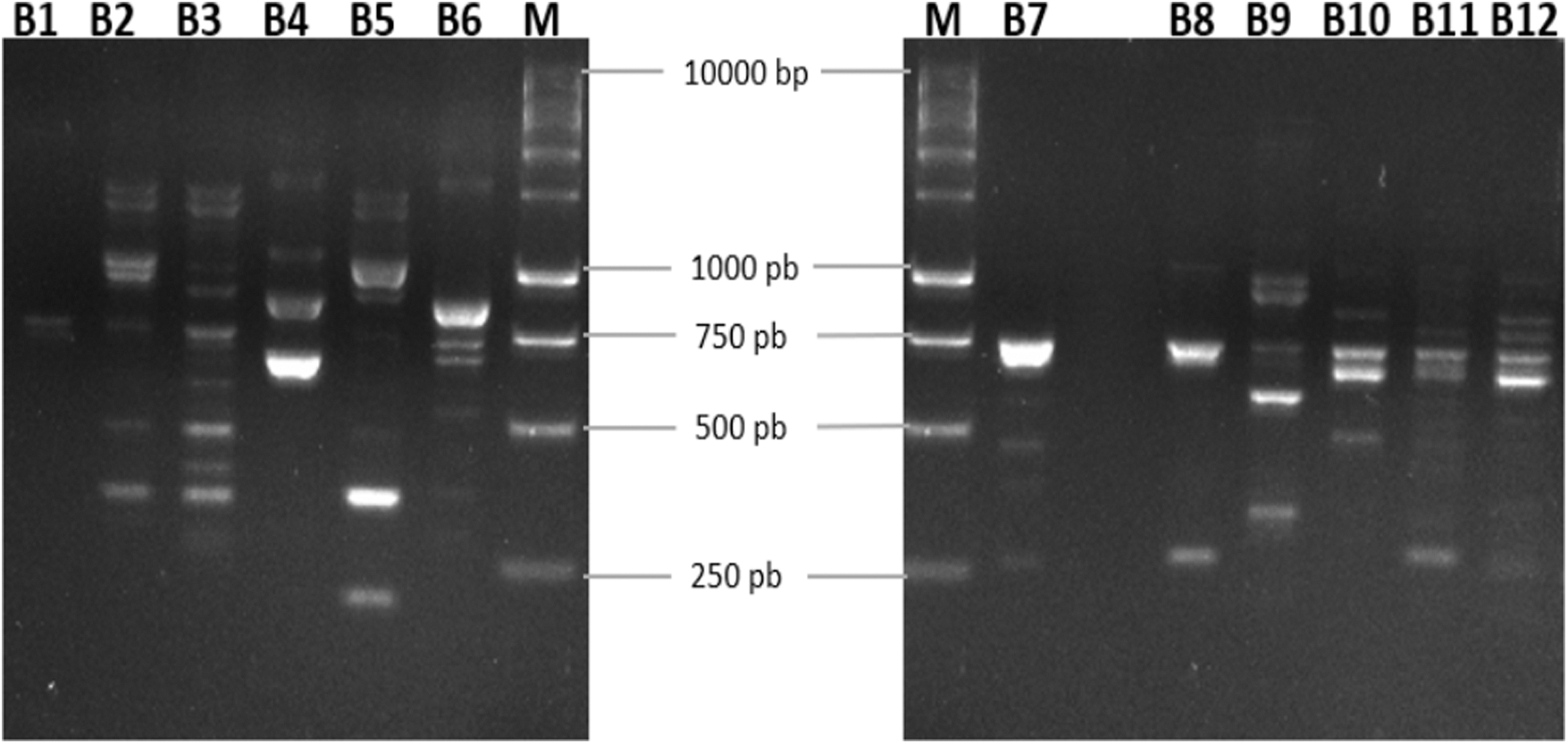
RAPD band patterns obtained from 12 different bacteriophages infecting Escherichia coli B using primer P2. B1: EbM3; B2: EbLoc1; B3: EbAbo1; B4: EbBor1; B5: EbAdi1; B6: EbCar1; M: 1 kb DNA Ladder; B7: EbB1; B8: EbB2; B9: EbB3; B10: EbBo1; B11: EbZ1; B12: EbZ2. RAPD, random amplification of polymorphic DNA.
The dendrogram issued from RAPD profiles analysis revealed the existence of four main clusters. These clusters matched largely with the bacterial species and most of the phages infecting the same bacterial species were clustered together excepted for cluster B that regroup phages infecting Escherichia coli C and E. cloacae (Fig. 5).
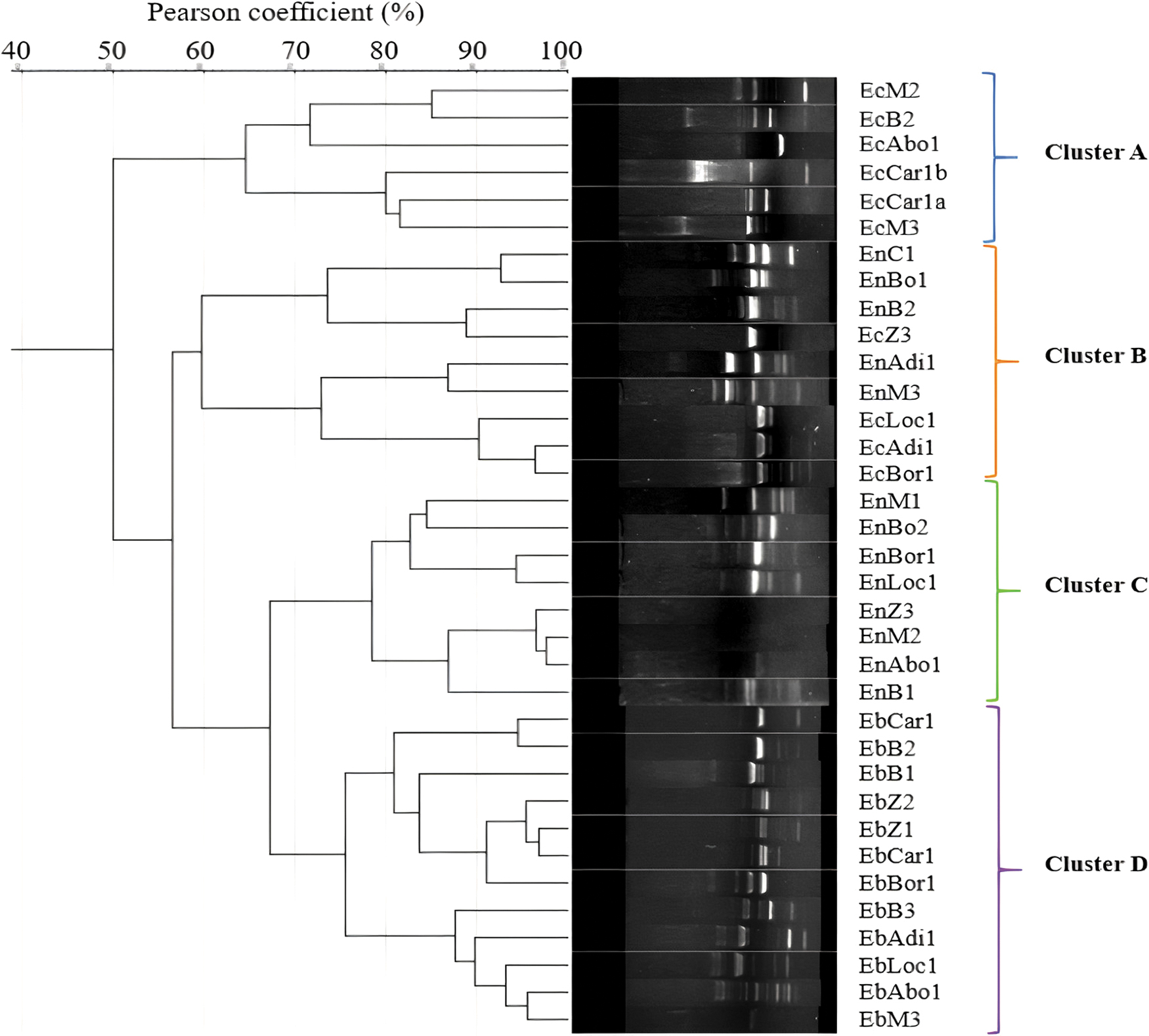
Dendrogram obtained after the analysis of RAPD band patterns. The similarity between samples was calculated based on the Pearson correlation coefficient and its corresponding dendrogram was constructed using the complete linkage method.
Discussion
Bacteriophages are easily isolated from sewage,5,6 and also from environmental water such as Ebrié lagoon41,42; however, according to our knowledge, E. cloacae-specific phages in sewage and environmental water have not been isolated previously in Côte d'Ivoire. In this study, water samples from urban sewage of Yopougon and Ebrié lagoon in Abidjan, Côte d'Ivoire were screened for the presence of E. coli and E. cloacae-specific phages.
With 30 samples and 90 attempted isolations, the collection contained 35 phages, with 9 infecting simultaneously E. coli and E. cloacae. Commonly, bacteriophages are thought to be species specific and even strain specific 43 as they are assumed to infect a narrow range of bacteria that are closely related. Recent studies have shown that polyvalent phages may be more widespread than previously assumed.22,23,44,45 Other studies have shown that sequential or simultaneous addition of multiple hosts is the most reliable way to produce phages that were able to infect bacteria of distantly related taxons.44,46 However, with these methods, the host range of these phages seems limited to the bacterial species used during their isolation.44,46 Nevertheless, as seen in this study, phages isolated on a single host could infect other hosts.22,47
Phages enriched with Ebrié lagoon samples have shown the highest titers. The occurrence of phages against a bacterial strain depends on the abundance of the host in sample. 6 This abundance may vary according to the environmental conditions such as temperature, pH, and concentration in cationic/anionic elements. We noticed the presence of detergent, discharge of chemical agents issued from domestic and industrial use in the wastewater drainage system of Yopougon. This could explain the lower phage titer observed in sewage samples. This difference in titer can also be explained by the enrichment conditions. Indeed, a variation of the culture conditions (quantity of culture medium, filtered sample, incubation temperature, and the choice of the host) can vary the results obtained after enrichment. 48
The genomic fingerprinting was useful to determine the diversity among the phages infecting the same species and/or isolated in the same location. A previous study has demonstrated that primers P2 provided high performance than P1. 38 The similarity matrix of RAPD profiles was calculated based on the Pearson correlation coefficient, and its corresponding dendrogram was deduced using the complete linkage clustering method. Usually, dendrograms from molecular typing in ecology are constructed based on average similarities among objects or on centroids of clusters.39,49 The resulting dendrogram does not show clearly separated groups. On the contrary, complete linkage clustering demands, for a group or object to agglomerate with another group, that all objects be related at the given similarity. As result, complete linkage clustering is much more robust, constraining, and contrasting. The clustering obtained with the dendrogram was not surprising because of the phylogenetic relations among phages. As it has been described previously, phages infecting distantly related bacterial hosts typically share few or no nucleotide sequence similarity, whereas phages infecting a specific bacterial host are more similar. 50 Cluster B that regroup phages infecting E. coli and E. cloacae based on their similarity despite they have different hosts may be due to module exchanging. The analysis of the genome of these phages may provide more data to evaluate this phenomenon among the studied phages. 51
Conclusions
Phages from two environmental sources have been investigated to screen for the presence of lytic phages. A collection of 35 phages infecting three bacterial strains has been obtained, 12 phages were isolated from Escherichia coli B, 10 phages from Escherichia coli C, and 13 with E. cloacae. The collection included nine polyvalent phages infecting simultaneously E. coli and E. cloacae. The RAPD-PCR revealed a genetic diversity among the isolated phages, rendering them interesting for application purposes.
Deep characterization is needed before any application. The sequencing of the bacteriophage genomes will be carried out to ensure that the phages carry no deleterious genes such as integrases or antimicrobial resistance genes. Transmission electron microscopy will be performed to determine phage morphology and classification. For practical use, testing on a large scale of bacterial strains is required.
Footnotes
Acknowledgments
We thank Sylvain Moineau of Félix d'Hérelle Reference Center for Bacterial Viruses (Laval University, Canada) for E. coli strains and bacteriophage T4 used in this study. Ms. Addablah was financially supported by a scholarship from the Ministry of Higher Education and Scientific Research, Lomé, Togo. We are grateful to Anicet Ebou who helped with the sampling map and statistical analysis.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The authors received no financial support for the research, authorship, and/or publication of this article.