
Abstract
Background:
Bacterial leaf blight (BLB) disease caused 80% of disease incidence in paddy in Kedah and Selangor states of Malaysia. The pathogenic bacterium, Xanthomonas oryzae pv. oryzae (Xoo), is one of the destructive pathogens infecting lowland irrigated and rainfed paddy in Asia’s tropical and temperate environments. Bacteriophages (or phages) have been proposed to control the pathogen due to their efficacy and safety aspects.
Material and Methods:
In this study, a total of 70 Xoo-phages were isolated from termite which living in rice-growing area.
Results:
2 lytic phages NΦ-1 and NΦ-3 were selected due to the high titer of the virus. Electron microscopic analysis showed that those phages belonged to the family Podoviridae, order Caudovirales with short noncontracted tails. Moreover, these phages have a narrow host range specifically target Xoo with a higher burst size. Whole-genome sequencing showed that the Xoo-phage NΦ-1 and NΦ-3 consists of a linear double-stranded DNA molecule of length 41,151 and 38,454 bp, respectively.
Conclusion:
This study successfully characterized two novel Xanthomonas phages and their potential as antimicrobial agents against BLB disease in rice.
Introduction
Rice (Oryza sativa L.) is the primary food grain consumed by almost half of the world's population. 1 It plays a significant role in world food production, which contributed 90% of production and consumption in Asian countries1–3 Moreover, the demands for rice increased every year due to the increase in the world population. It provides 40% of the total calorie and supports 60% of the population; more than 3.5 billion people count on rice for their daily calorie intakes.4,5
Rice is vulnerable to several diseases such as bacterial blight, blast, and sheath blight. 6 In September 2017 and April 2018, Azizi et al. reported that rice varieties (MR 269 and CL varieties) were infected with bacterial leaf blight (BLB) disease and caused 80% disease incidence in Kedah and Selangor states of Malaysia. 7 The disease is mainly caused by the infection of Xanthomonas oryzae pv. oryzae (Xoo), particularly at the lowland irrigated and rainfed paddy in Asia's tropical and temperate environments. 8 It is quickly developed when the temperatures fall to 28–34°C with >70% relative humidity.9,10 Moreover, infection of plants of BLB disease is highest at tillering stage, causing a 20–50% reduction in the harvest.11,12
Bacteriophages (phages) are abundant in the environment, for example, soil and seawater, oceanic and terrestrial surfaces, and extreme environments. 13 They have also been detected in hospitals, in wastewater, on animal and human tissues where their bacterial host can be found. 14 Many phages have been isolated and characterized from the nature against plant pathogens such as Xanthomonas arboricola pv. juglandis, 15 Xanthomonas syringae pv. porri, 16 Xanthomonas axonopodis, 17 and Xoo.18,19
Phage therapy is a technique of utilizing phage as an antimicrobial agent to treat infectious diseases caused by pathogenic bacteria. 20 It is specific in targeting bacteria, ability of self-limiting, and self-replication in the new cycles of infection.13,21,22 In addition, phages infect bacterial host that displays specific receptor on their surface that allows these phages to bind and penetrate into the cells. 23 By exploiting this characteristic, phages that target specific bacterial host could be isolated, for instance, Xanthomonas spp. phages, have been described.15,24, Recently, Dong et al. have successfully isolated Xoo-sp2 phage from the soil of the Xoo-infected rice-growing area. 19 The phages were characterized by morphology, host range, and genomic analysis that helps in understanding their efficacy in controlling the bacteria.
In this study, novel phages against rice pathogen Xoo were isolated from termites from the paddy field located at Serdang, Selangor. Two isolated phages were characterized based on morphology, host range, a multiplicity of infection (MOI), adsorption rate, and one-step growth curve. In addition, whole genome sequencing was also conducted.
Materials and Methods
Sample collection
Two jars of termite samples were obtained from the rice fields at Ladang 10, UPM (2.9993°N, longitude: 101.7079°E). Phages were isolated immediately in the laboratory.
Isolation of bacteriophage
One milliliter of an overnight Xoo culture was mixed with 10 mL of fresh Luria Bertani (LB) broth (Merck, Darmstadt, Germany). The mixture was allowed to incubate for 37°C with agitation, till the absorbance of the culture reach OD600nm 0.6–0.8. The termite sample was ground into powder and mixed with 10 mL of sodium magnesium (SM) buffer (100 mM of NaCl, 8 mM of MgSO4·7H2O, 50 mM of Tris-HCl, pH 7). The mixture was centrifuged (10 min, 12,000 rpm), and then the supernatant was obtained. Five milliliters of the mixture was mixed with Xoo culture and incubated for 12 h with 200 rpm agitation at 37°C. After that, 10 mL of the mixture was centrifuged (15 min, 12,000 rpm) to sediment the bacteria and debris. The supernatant was collected and filtered through a 0.22-μm syringe filter (Bioflow, Malaysia).
Phage titration
The double-overlay agar method was used according to Adams with slight modifications. 25 In brief, 1 mL of overnight Xoo strains culture was transferred into 10 mL of LB broth, which was incubated at 37°C with agitation (200 rpm) until it reached the mid-log phase (108 cfu/mL) of bacterial host strains, with an absorbance of OD600nm 0.6–0.8. Then, 100 μL of the Xoo was incubated together with 100 μL of phages suspension at 37°C for 20 min to achieve phage attachment to bacteria. Next, 2% of 1 M CaCl2 was introduced into the mixture to stabilize the phage adsorption process. The mixture was then added into the top agarose (1% of Bacto-tryptone, 0.5% of yeast extract, 0.5% of NaCl, 0.1% of MgCl2·6H2O, and 0.7% of agarose; 4 mL at 45°C) and poured onto a sterile agar plate. The plates were allowed to solidification and incubated at 37°C for 12 h. Then, plaques formed and phage titer was determined, respectively.
Purification of phages
The double-overlay agar method has been described previous to obtain single plaques. Plaques of varying sizes and morphologies were picked from the overlay plates and separately mixed with a 10 mL of mid-log phase Xoo strains culture and incubation (37°C, 200 rpm, 24 h). After 24 h, phages were centrifuged (10,000 rpm, 15 min, 4°C), and the supernatants were collected with filtered, as described previously.
Polyethylene glycol precipitation
Phage was precipitated using polyethylene glycol (6 × polyethylene glycol (PEG)/NaCl [20% PEG 8000 and 2.5 M NaCl]; Merck) at 4°C overnight. The precipitated phage was obtained by centrifugation (60 min, 12,000 rpm, 4°C). The phage pellet was resuspended in SM buffer and stored in −20°C freezer as virus stock for the subsequent studies.
Propagation of bacteriophages
Propagation of phage was performed by the double-overlay agar method as described previously. After 24 h, phage particles were obtained by adding 5 mL of SM phages buffer onto each plaques-containing plate for 10 min, and the top agar layer from all plates was discarded. The collected overlay was obtained and incubated at 37°C with agitation for 1 h. These mixtures were treated with chloroform at a final concentration of 0.2% (v/v), then centrifuged (10,000 rpm, 15 min; Centrifuge 5810 R; Eppendorf). The obtained supernatant was filtered, and the phages solution was precipitated with PEG as described earlier. The purified phages were stored at −20°C freezers for subsequent studies.
Characterization of Xoo phages
Phage host range study
The host range of the isolated phages in this study was determined based on phages' ability to infect different types of bacteria species, as detailed in Table 1. Host range was analyzed according to the method described by Manchester et al. with slight emendation. 26 All testing bacteria were grown in a fresh LB broth with shaking until the mid-exponential phase (about 108–109 CFU/mL). Phage activity was scored on the basis of plaque formation, which indicated host cell lysis. The presence of plaques was recorded as (+), whereas (−) when no plaque formation was detected. The plaques were categorized as clear or turbid. The experiment was performed in triplicates.
Bacterial Strains Used for Phage Sensitivity Determination
Microbial Culture Collection Unit, Institute of Biosciences, Universiti Putra Malaysia.
Phage morphology using transmission electron microscopy
A sterile high-titer phages lysate (108 pfu/mL) was precipitated using PEG/NaCl and centrifuged (60 min, 12,000 rpm; Centrifuge 5810R; Eppendorf). The supernatant was removed, after which ammonium acetate solution (0.1 M) was introduced. The procedure was repeated twice. The final sediment was resuspended in SM buffer. The purified bacteriophage samples were negatively stained with 2% (w/v) aqueous uranyl acetate (pH 4.0). It was then deposited on the U.V. activated carbon-coated Formvar films, air-dried, and examined under FEI Tecnai transmission electron microscopy (TEM) Machine (FEI, Eindhoven, The Netherlands) at magnifications of 10,000 × to 50,000 × . Phages were classified, based on their morphology, into their respective families as per the International Committee's guidelines on Taxonomy of Viruses. 27
Multiplicity of infection
The ratio between virus particles and host cells can be measured by MOI. According to Gašić et al.'s method with minor emendation to analyze the optimum MOI of Xoo phages. 24 The Xoo culture were incubated and with 200 rpm agitation until it reached the mid-exponential phase (OD600nm of 0.6–0.8, ∼108 CFU/mL). Two milliliters of the bacteria culture was infected with phages separately at four various MOI ratios (∼0.001, 0.01, 0.1, and 1) and was subjected to incubate (37°C, 30 min).
Phage adsorption
The adsorption experiment was performed according to Ellis and Delbrück with minor modification, the amount of phage adsorbed to the host cell for a specific period of time was determined. 28 Xoo in mid-exponential phase growth (108 CFU/mL; 100 mL) was infected with the phage suspension at MOI of 0.1 and was subjected to incubate at 37°C with 200 rpm agitation. At desired interval time (1, 3, 5, 7, 10, and 20 min), the mixture was treated with chloroform (final concentration of 0.2% [v/v]), and to sediment the phage-adsorbed bacteria by centrifuge (10,000 rpm, 2 min). Titers of unadsorbed free phages in the supernatant were processed by the double-overlay agar method as described earlier. Plaque form was counted. All experiments were performed in triplicates.
One-step growth curve
The one-step growth curve experiment was carried out the protocol adapted from Ellis and Delbrück. 28 Xoo culture was allowed to grow in 10 mL of LB broth until it reached the mid-exponential phase (OD600nm of 0.6–0.8, ∼108 CFU/mL). Meanwhile, phage (108 pfu/mL) was prepared and kept in the 37°C for 10 min. Then, 1 mL of the phage was added into 9 mL of the host culture, and was mixed well. The mixture was subjected to incubate at 37°C with shaking for 10 min to achieve absorption of phages to bacteria. The phage-adsorbed cells were obtained after centrifugation (10,000 rpm, 5 min). Ten milliliters LB broth was transferred into tube to resuspend the precipitate of the bacteriophage debris and incubated (37°C, 200 rpm). An aliquot of 0.1 mL was collected every 10 min for a period of 180 min. The mixture was serially diluted in SM phage buffer from 10–1 to 10–9, and phage titer was then determined. All assays were performed in triplicates.
DNA extraction of Xoo-phages
Five hundred microliters of phage lysate was mixed with lysis buffer (10 μL of 20 mg/mL proteinase K, 2.5 mg CTAB, 5 μL of 1 M EDTA 50 μL of 1.5 M Tris-HCl [pH 8.5], 375 μL NaCl [2 M] and 1 μL of 2-mercaptoethanol) for 60 min at 56°C to extract the phage's DNA. Subsequently, The DNA band was visualized using Gel documentation system. The genome DNA was sent to Apical Scientific Sdn. Bhd. to analysis. Illumina raw reads were first removed of primer sequences using bbduk of the BBTools Packages. Unassembled short reads were mapped to the two reference genomes (NC_020205.1 and NC_012742.1) using Bowtie2. To verify presence of phage DNA, QC reads were assembled de novo using Spades and polished using Pilon version 1.23, implemented in Unicycler. All contigs were subjected to BLASTN analysis against the NCBI nucleotide database.
Results
Bacteriophages isolation, purification, and plaque morphological analysis
A total of 70 phage isolates were successfully isolated from termites at Ladang 10, UPM, using Xoo as a host. All phages formed clear zones with different sizes and morphologies, as represented in Figure 1. Sixteen phages were further propagated and purified. A high-titer suspension ranged from 105 to 109 pfu/mL was obtained. Among these phages, NΦ-1 and NΦ-3 were selected for further study based on the plaque size and high titer, as shown in Table 2. These phages formed >4-mm-diameter plaques on the lawns of host strain Xoo strain.
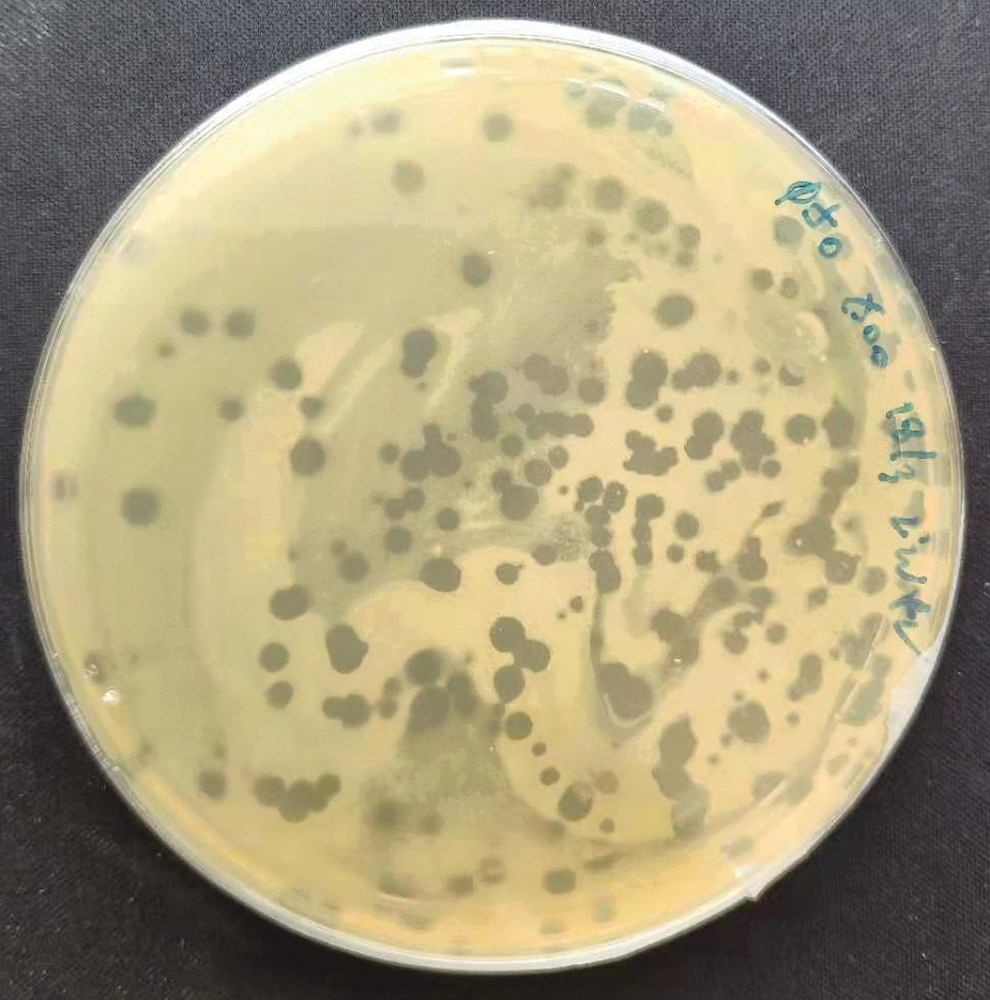
Plaque formed on LB agar isolated phages from termites and plaque assay was performed in agar plate with Xoo. LB, Luria Bertani; Xoo, Xanthomonas oryzae pv. Oryzae.
Phage Isolated Against Xanthomonas oryzae pv. oryzae from Termite
pfu, plaque forming units.
NΦ-1 and NΦ-3 showed high potential to inhibit Xoo strains' growth on plates. All phages formed large (4–6 mm) with high titer (108–109 pfu/mL) (Table 2) and clear plaques, as shown in Figure 2. A clear plaque is indicating complete lysis of bacterial host in the plaque.
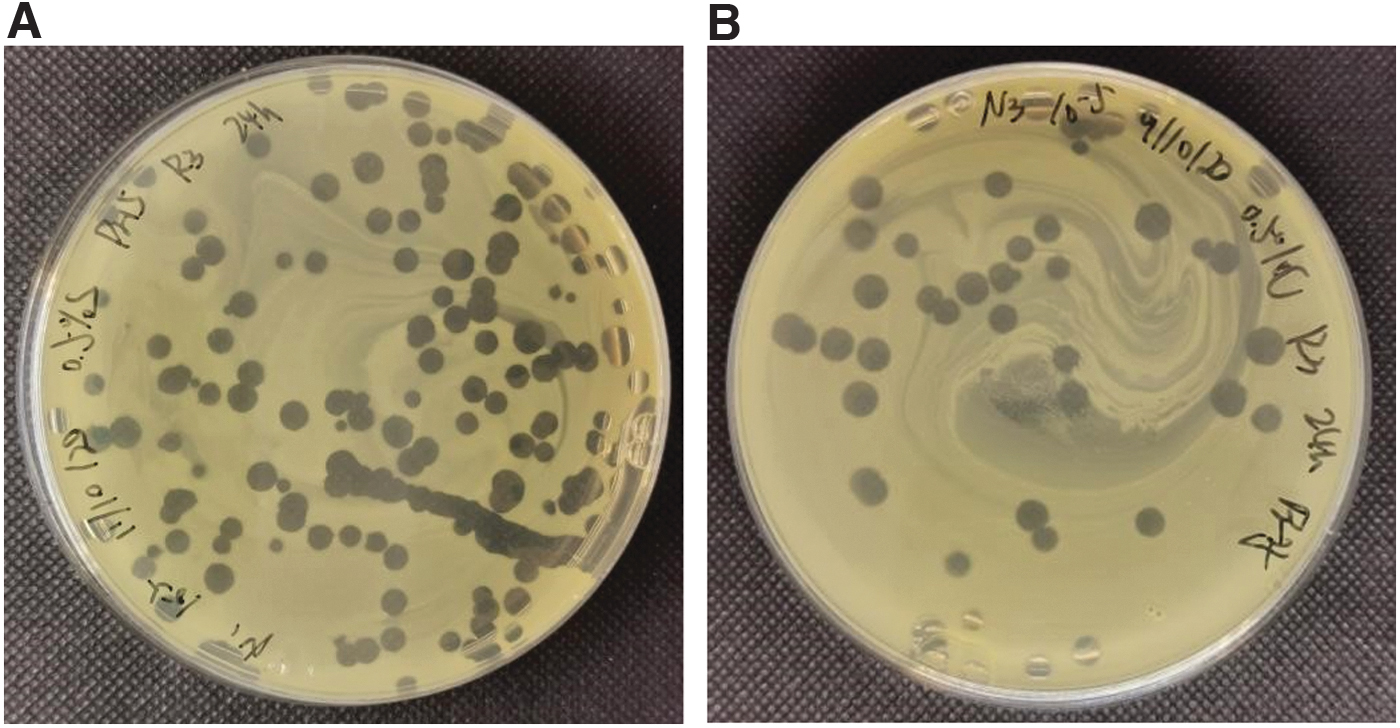
Double layer agar plates showing plaque purified after repeating plaque assay and filtering phage lysate through 0.22-μm filter. Plaques obtained were in the range of 4.5–6 mm. Plate shows plaques of high dilution of
Host range study
The host range of NΦ-1 and NΦ-3 phages were investigated on various bacterial isolates (Table 3). Interestingly, these two phages were unable to lyse Pseudomonas spp., Bacillus siamensis, Bacillus methylotrophicus, Bacillus subtilis, Bacillus marisflavi, and Bacillus pumillus. This result showed that the isolated phages have a narrow host range
Host Range of Isolated Phages Against Different Phytobacteria Strains
+, presence of plaque; −, absence of plaque.
Morphological study by TEM
Morphology of the NΦ-1 and NΦ-3 isolates was observed by TEM (Fig. 3). From the TEM images obtained, all three phages were assigned to the order Caudovirales. They have short noncontracted tails and belong to the family Podoviridae, which are tailed phages with dsDNA. 27 Head of NΦ-1 and NΦ-3 isolates were 105.2 ± 0.6 and 53.6 ± 0.4 nm in diameter (means ± standard deviations), respectively. The length of tails was 32 ± 0.7 nm for phage NΦ-1 and 12.3 ± 1.4 nm for phage NΦ-3.

Transmission electron micrographs of lytic phages.
Multiplicity of infection
The optimum MOI of phages NΦ-1 and NΦ-3 was determined by counting the average of pfu at four different MOIs. The result showed that NΦ-1 and NΦ-3 isolates have optimum MOI of 0.1 (Table 4) showing 3.5 × 108 and 4.1 × 107 pfu/mL, respectively.
Determination of Optimal Multiplicity of Infection for NΦ-1 and NΦ-3 Xanthomonas oryzae pv. oryzae-Phage at 37°C
Plaques formed were counted and calculated as pfu/mL.
Phage adsorption
The percentage of absorption for NΦ-1 increased rapidly from the beginning and reaching a peak of 62.57% at 7 min (Fig. 4). It decreased slightly after 10 min but rebounded again until it reached 57.89% at 20 min postinfection. Similarly, the NΦ-3 phage reached an apex point at 7 min with a percentage of adsorption of 74.77%. It was reduced after 10 min and again increased to 72.83% after 20 min.

Adsorption of phages (%) to Xoo at various time point. The experiment was performed to determine the number of phage particles adsorbed to host cells within a particular period (0, 1, 3, 5, 7, 10, 15, and 20 min) for
One-step growth curves
The one-step growth curves for the two Xoo phages NΦ-1 and NΦ-3 were carried out to determine the burst sizes and latent periods. According to the triphasic curves obtained, the latent time for NΦ-1 and NΦ-3 was estimated to be about 10 and 20 min, respectively, whereas the burst size of these phages was 3.4 × 105 and 4.2 × 105 pfu/mL, respectively (Fig. 5).
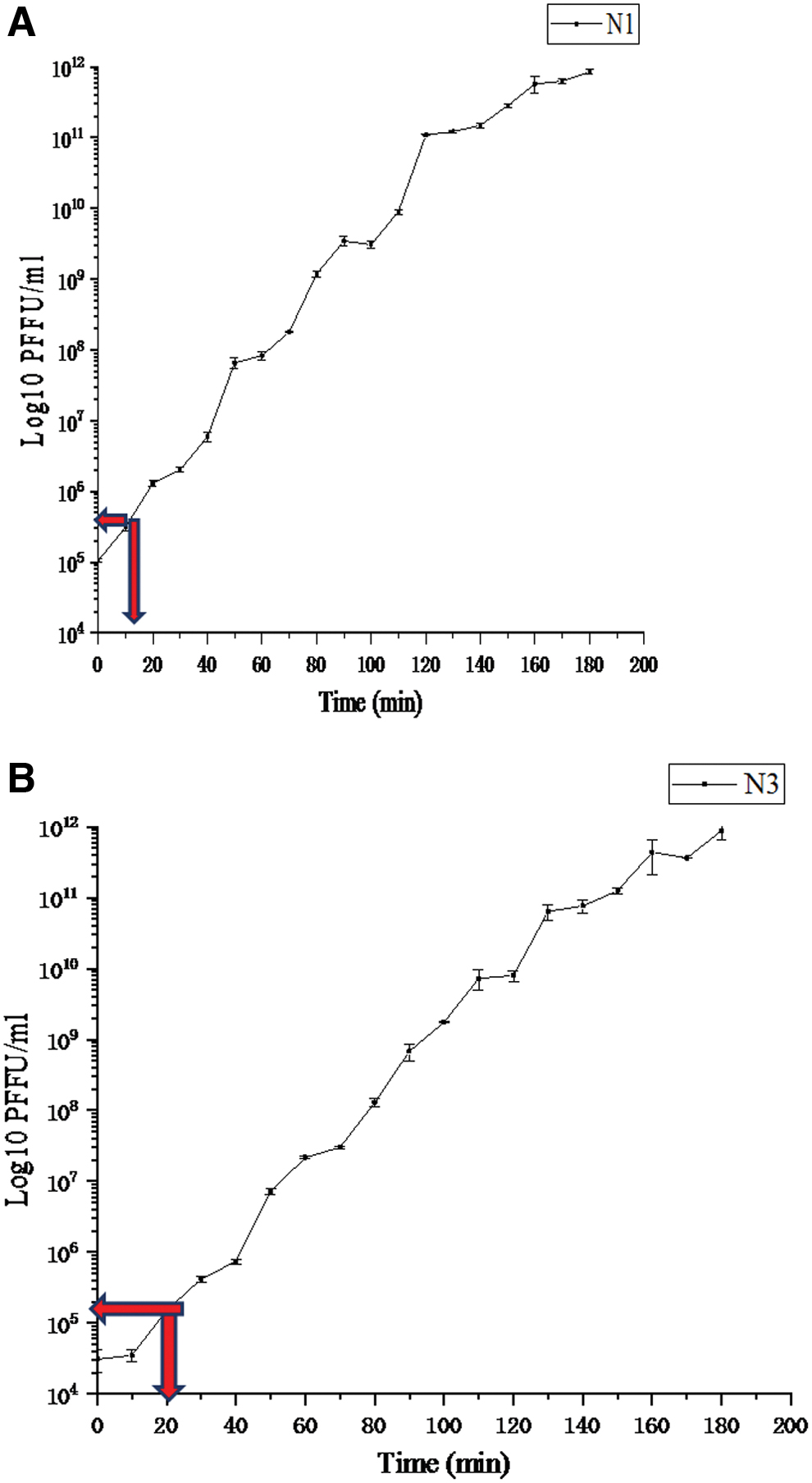
One-step growth curves showing latent period and burst size values of Xoo phages
Genomic features of Xoo phages
The sequencing of the NΦ-1 and NΦ-3 was sequenced and deposited in NCBI. These genomes revealed a circularly permuted linear dsDNA genome with a size of 41,151 and 38,454 bp, respectively (Fig. 6). the NΦ-1 and NΦ-3 phage with a same G + C content, which is 62%. In addition, the both genomes showed 29 predicted genes and no tRNAs. Then, the majority of structural protein will focus on 18 proteins through encode for both phages. The genomes were compared against available phages genome in GenBank database. Holin and endolysin genes are found both in these two phages genomes. The holin gene, unlike the conserved endolysin gene, is found by predicting the transmembrane domains structure in SWISS-MODEL. A nucleotide comparison using Blastn to available sequences in the NCBI NR database shows that the NΦ-1 phage genomes have the largest similarity with Xanthomonas phage Pagan MK903278.1 (7% query coverage, 74.56% identity, and 44,448 bp); Xanthomonas phage phi Xc10 NC_047840.1 (10% query coverage, 73.89% identity, and 44,597 bp); and Xanthomonas phage f30-Xaj NC_030937.1 (3% query coverage, 76.00% identity, and 44,262 bp). However, The NΦ-3 phage also showed the most significant similarity with these phages, as has been described earlier, whereas the similarity is 8% query coverage, 74.56% identity; 11% query coverage, 73.89% identity; and 3% query coverage, 76.00% identity, respectively.
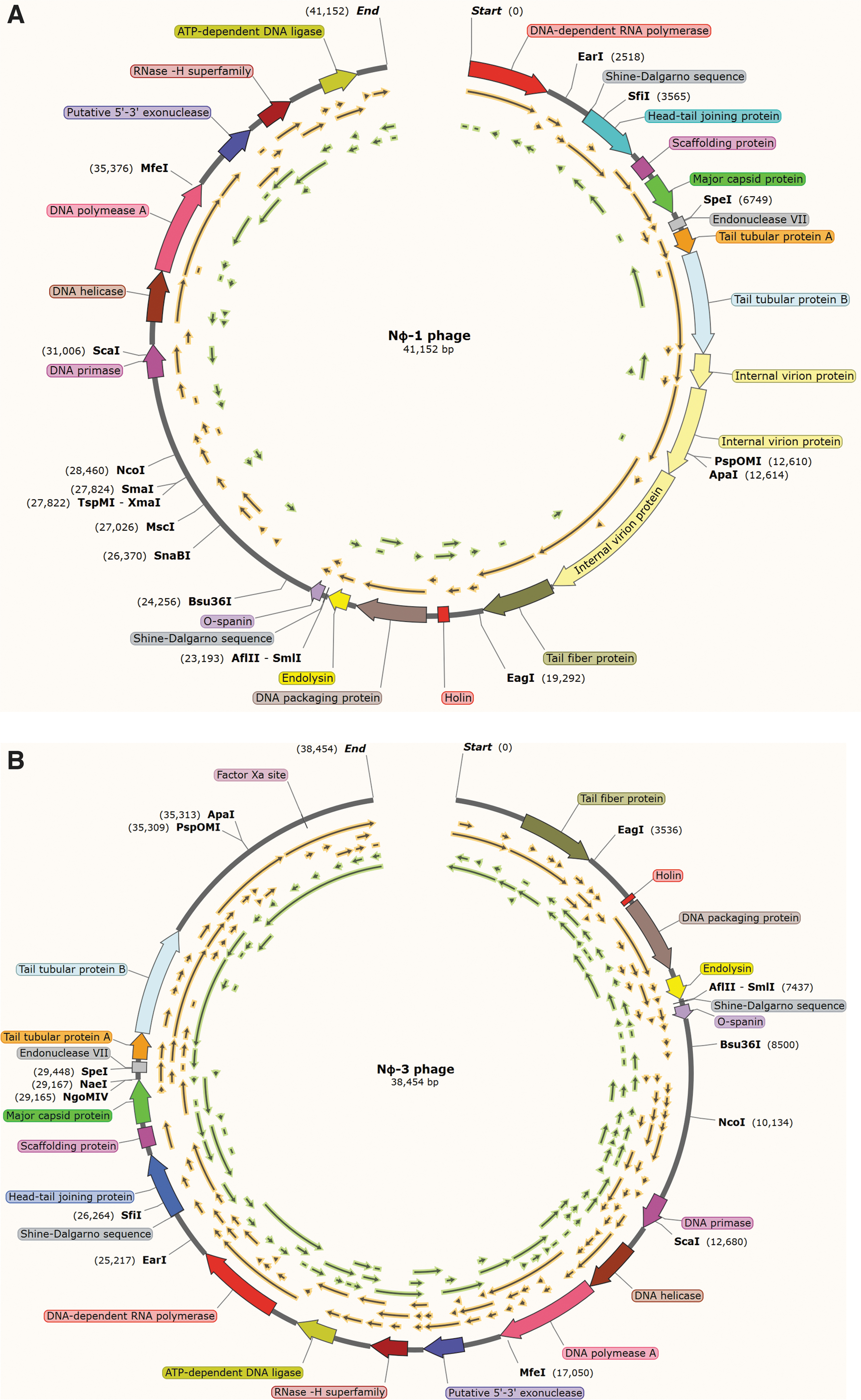
Illustration of phage genome and the respective gene encoded for
Discussion
Xoo is one of the destructive pathogens that causes BLB disease leading to considerable rice losses.29,30 Chemical control agents such as bactericides, streptomycin, chloramphenicol, and niclosamide are used broadly to control this disease31–33 Nevertheless, these chemicals are ineffective in controlling the disease development and outcome. 34 Therefore, alternative approaches such as biological control are anticipated. Bacteriophage (Phage), a virus that infects explicitly bacteria, could be applied as a biological product to control plant diseases attributed to bacteria. 35 In 2018, Dong et al. successfully isolated and characterized Xoo phages from the paddy field soil sample. 19 In our study, we are interested to isolate novel Xoo phages from termites that live in paddy field. Termites are economic pests in tropical and subtropical environments36–39 They are also found abundantly in tropical rainforests. 40 The gut bacteria in termite are subjected to many environmental phages since termite is a soil-dwelling insect. It was reported that Citrobacter phage CVT22 was successfully isolated from the gut of the Formosan subterranean termite 41 indicating the presence of an unexplored phages community in the termite gut.41,42 Similarly, we have successfully isolated phages from the termite Xoo as the bacterial host. During the course of selection, calcium ion, Ca2+ was added during the adsorption of virus to bacterial host. It was reported that Ca2+ could enhance the adsorption and hence increase the virus titer.43–46 Two novel phages, NΦ-1 and NΦ-3, which are capable of producing large clear plaques and high titers were selected for the subsequent characterization.
The specificity of the isolated phages to infect targeted bacterial host is an advantage despite their narrow host range. This prevented nonspecific infection, sparing the nontargeted beneficial plant growth-promoting rhizobacteria (PGPR). 47 PGPR are free-living root colonizing bacteria that are beneficial to plants. They reduce disease severity and enhance many crops' yield.48,49 Azotobacter, Bacillus, and Pseudomonas are considered as PGPR that enhance plant growth through nutrient recycling and nitrogen fixation to increase the yield of crops. 50 In addition, Udayashankar et al. demonstrated that seven PGPR strains, namely, Bacillus pumilus INR7, SE34, and T4, Bacillus amyloliquefaciens IN937a. B. subtilis IN937b, and GB03, and Brevibacillus brevis IPC11, were capable of protecting rice plant from BLB caused by Xoo under in vivo and in vitro conditions. 51 Similarly, Vasudevan et al. also showed that Pseudomonas spp. and Bacillus spp. have the potential to suppress the infection of Xoo in rice preventing the BLB disease. 52 In this study, the isolated phages NΦ-1 and NΦ-3 are considered PGPR-friendly as they were unable to infect those bacterial strains of PGPR group.
Most phages are classified into the family Siphoviridae (61%) with long and noncontractile tail, Myoviridae (24.5%) with contractile tail, or Podoviridae (14%) with a short tail. 53 NΦ-1 and NΦ-3, which possess a short noncontractile tail, belong to the family Podoviridae. Classification of phages using electron microscopy is important as phages that target the same genus Xanthomonas could be from different family. Romero-Suarez et al. reported that of 36 phages against X. arboricola pv. Juglandis, 16 phages isolated from the phyllosphere belong to the family Podoviridae, whereas 20 phages isolated from the rhizosphere belong to the family Siphoviridae. 15 Most Xoo phages, including Xanthomonas virus CP1, OP1, phil7, XOP411, XP10, and Xoo phage XOO-sp2, belong to the family Siphoviridae as it is the most abundant phage in the environment.19,23,54 Apart from Siphoviridae and Podoviridae, phages of Myoviridae such as Xanthomonas euvesicatoria, phages KΦ1 and KΦ15, were also reported.24,18
In one-step growth curve analysis, NΦ-1 and NΦ-3 showed a short latent period with high burst size, a favorable characteristic that we sought after. This indicated that they are specifically active against a large number of Xoo host strains. 55 Similar to our findings, Ranjani et al. also showed that ϕXOF4 has a short latent period and high burst size (1.8 × 107 pfu/mL). 56 However, a short latent period with low burst size was also mentioned, of which only 153 and 92 pfu/mL of ϕIBB-PF7A and ϕPSA2 phages were produced.57,58 A long latent period with low burst size, such as P3k, P4A, P7A, and P8D phage, was reported with only 60, 23, 29, and 331 per infected host cell, respectively. 59
For the phage adsorption, NΦ-1 and NΦ-3 were rapidly increased to 62.57% and 74.77% at an apex point of 7 min. It was subsequently stabilized and reached the highest percentage at 20 min. Researchers are seeking for phages that have fast adsorption rate whereby it increases the changes of phages to infect the targeted pathogens, whereas reducing the environmental induced inactivation of free phage. High phage adsorption was reported for KΦ 1, KΦ 8, and KΦ 15 phage toward Xanthomonas euvesicator, showing 90%, 88%, and 72.4% at 20 min, respectively. 24 PRΦL2 phage prevents the bacterial blight disease in pomegranate caused by Pseudomonas sp. and shows 66% at 20 min. 60 In addition, Ranjani et al. also showed that Φs;XOF4 phage against Xoo exhibited high adsorption (58%) at 20 min and continued to increase to 70% at 60 min. 56
Bacteriophages have been listed in innovative biotechnology to devastate foodborne infective bacteria. 61 Detailed analysis of the phage genome is crucial to identify potential gene(s) that is/are responsible for the bacterial host lysis. The complete genome of NΦ-1 and NΦ-3 was sequenced and analyzed. Two important genes, namely holin and lysin, were identified in the genome. Holin is important for host cell membrane lysis, regulating the timing of lysis, as well as degrading the peptidoglycan. 62 In addition, Lysin (i.e., endolysin), a hydrolytic enzyme that is capable of degrading peptidoglycan of bacterial cell wall, would promote the release of phage. 63 The lysins were reported to affect against G (+) bacteria owing to lack of an outer membrane.64–67 Lysins also demonstrate specificity for certain peptidoglycan types, which could help antimicrobial substances specifically reach to targets. 63 Taken together, it can be concluded that the novel Xoo phages NΦ-1 and NΦ-3 are good candidates for phage therapy, which warrants a further investigation of their potential in controlling the BLB disease in rice. It is known that holin is related to the host cell membrane lysis cycle. Moreover, certain phage late proteins such as lysins and holins are intimately associated with the lysis of host cell and burst of virion to the environment. 68 It is also feasible to make some mutations to designed loci to make holin functional and stable in different conditions, which can be applied to varied soil environment. There is a promising future of holin to be used as an antibacterial agent and consequently benefiting in planting industry.
Footnotes
Acknowledgments
Authors thank Ministry of Higher Education (MOHE) of Malaysia for funding support under Fundamental Research Grant Scheme (FRGS/1/2018/WAB01/UPM/02/7) and Universiti Putra Malaysia (UPM) for funding support under GP Grant (9652100).
Ethical Statement
There is no animal study in this work.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by Ministry of Higher Education (MOHE) of Malaysia under Fundamental Research Grant Scheme (FRGS/1/2018/WAB01/UPM/02/7) and Universiti Putra Malaysia (UPM) under GP Grant (9652100).