
Abstract
Pathogenic bacteria are attracted toward mucosa, as it is their way of entry into the body. However, we know surprisingly little about the phage-bacterium interactions in the mucosal environment. Here, we explored the effect of the mucosal environment on growth characteristics and phage-bacterium interactions in Streptococcus mutans, a causative agent of dental caries. We found that although mucin supplementation increased bacterial growth and survival, it decreased S. mutans biofilm formation. More importantly, the presence of mucin had a significant effect on S. mutans phage susceptibility. In two experiments done in Brain Heart Infusion Broth, phage M102 replication was detected only with 0.2% mucin supplementation. In 0.1 × Tryptic Soy Broth, 0.5% mucin supplementation led to a 4-log increase in phage titers compared with the control. These results suggest that the mucosal environment can have a major role in the growth, phage sensitivity, and phage resistance of S. mutans, and underline the importance of understanding the effect of mucosal environment on phage-bacterium interactions.
Introduction
Many pathogenic bacteria use mucosal layers to invade metazoan hosts. Indeed, the mucosa is an important chemotactic signal for many pathogenic bacteria,1,2 directing the bacteria toward the metazoan host. Mucosal cues cause expressional changes in virulence-related genes of many bacterial species. 3 For example, interaction with mucus and mucin glycoproteins impacts bacterial phenotypic characteristics and increases virulence of many pathogenic species,4–6 including Acinetobacter baumannii 7 and Streptococcus mutans. 8 Further, some bacteria, such as pathogens and many intestinal anaerobes, can use mucosal components directly as a nutrient source. 5
The mucosal surface consists of secreted mucin glycoproteins, which oligomerize to form a complex viscous milieu allowing the retention of antimicrobial molecules (e.g., immunoglobulins A and G, lectins, antimicrobial peptides) and also phages. More than 20 families of mucin glycoproteins exist, and they are expressed in a tissue-specific manner.1,9 Mucin secretion is primarily constitutive, leading to a dynamic gradient of mucins—high close to the epithelial cell surface but lower in the outer surface of the mucosa.
The past 10 years of research have clearly shown that phages are major partners in the mucosal microbiomes,10,11 thus affecting both the invading and local bacterial populations. Understanding the ecology of pathogens and their phages in the mucosa is central for our health, and for the development of phage therapy approaches against antibiotic resistant infections. However, we know surprisingly little about the phage-bacterium interactions in the mucosal environment. This has led to fundamental gaps and biases in our knowledge regarding the antagonistic interactions between phages and bacteria as they happen in the mucosal milieu.
The oral environment is an important habitat for phage-bacterium interactions.12,13 The salivary microbial community has been shown to host 108 virus-like particles per mL (mostly dsDNA phages) 14 whereas in the dental plaque the density is higher, 1010 virus-like particles per gram. 15 Since previous research suggests that mucosa may influence the virulence of bacteria4–6 and phages,6,16,17 and favors phage resistance mechanisms that maintain bacterial virulence (e.g., CRISPR-Cas), 18 it is important to describe how the mucosal environment shapes these interactions in S. mutans, a causative agent of dental caries.
Here, we characterized the effect of mucosal environment on growth features and phage-bacterium interactions in S. mutans. In its natural habitat, S. mutans forms biofilm and is in continuous contact with oral mucosal surfaces. It has been previously demonstrated that several salivary components inhibit S. mutans biofilm formation, but the presence of sucrose can reverse this effect. 19 However, so far there has been no direct demonstration of the impact of mucins on the interactions between S. mutans and its phages.
This aspect is central for understanding the phage-bacterium interactions in the mucosa, and for developing successful phage therapy approaches against caries. Here, we characterized the impact of mucosal environment on S. mutans growth characteristics in physiologically relevant mucin concentrations.20,21 We found that mucin supplementation decreases S. mutans biofilm formation, whereas it increases bacterial survival in starvation. More importantly, mucin has a central role in S. mutans susceptibility to phage infections, and, consequently, phage resistance.
Materials and Methods
Host and phage strains
Streptococcus mutans OMZ381 and its phage M102 were obtained from Félix d'Hérelle Reference Center for Bacterial Viruses (www.phage.ulaval.ca) and stored in −80°C. In the experiments, the bacteria were cultured in Tryptic Soy Broth (TSB) or Brain Heart Infusion Broth (BHI) at 37°C. Phage stocks were prepared by harvesting the soft-agar of a confluent double-agar plate, to which 4 mL of media were added, followed by centrifugation (11,000g) and filtration. Phage titration was made by plaque forming unit counts using the double-agar method.
Bioinformatic analysis
Phage M102 22 genome is publicly available (RefGenome: NC_012884). We used HHpred23,24 to analyze M102 ORFS and to find hits associated to Ig-like domains and carbohydrate binding.
Phage binding to mucin
To evaluate phage M102 binding to mucin, BHI (Sigma) agar plates were prepared with or without 1% mucin in its composition. Purified porcine mucin (Sigma; catalog no. M1778) was used as a source. The phage was diluted to a concentration of 2.5 × 102 plaque forming units (PFU) mL−1 in liquid BHI media, and 5 mL of the dilution were added to the plates containing mucin or not. The plates were kept under agitation for 30 min, then the liquid was removed by careful pipetting, and 3 mL of BHI soft agar containing the S. mutans host was added to each plate. Plates were incubated overnight at 37°C, and plaques were enumerated. The experiment was done in triplicate.
Influence of mucin on S. mutans growth and biofilm formation
The influence of mucin supplementation on S. mutans was studied first in 1 × (i.e., undiluted) TSB (Sigma) supplemented with 0.5% yeast extract and 0.5% K2HPO4). Fresh bacterial culture was inoculated into 5 mL of TSB supplemented with 0, 0.05 or 0.1% mucin in triplicate, and it was cultured overnight at 37°C under constant shaking (200 rpm). Optical density (OD) (at 595 nm) of the cultures was measured in 6 technical replicates per tube. Data were analyzed using Kruskal–Wallis non-parametric analysis of variance (ANOVA) in SPSS.
In a second experiment, we explored the ability of S. mutans to grow with mucin without any additional energy source. Bacteria were inoculated in 5 mL of sterile H2O without mucin and with 0.2% and 0.5% mucin supplementation in triplicate. These cultures were grown at 37°C under constant shaking (200 rpm). Bacterial growth was recorded by estimating colony forming units (CFU) mL−1 via applying 2 μL drops of the 10-fold dilutions of culture on BHI plates. Data were analyzed with Kruskal–Wallis non-parametric ANOVA in SPSS.
Finally, we explored how different nutrient conditions and mucin concentrations influence S. mutans biofilm formation. In this experiment, overnight-grown S. mutans was inoculated 1:10 000 into sterile H2O, 0.1 × BHI (i.e., 10 times diluted BHI) and 1 × BHI, supplemented with 0%, 0.2%, or 0.5% mucin. Each treatment was done in six replicates in 150 μL volume, on a sterile 96-well plate, and incubated for 2 days at 37°C without shaking.
To determine the biofilm formation, all medium was removed from the wells. Plates were washed three times with MilliQ water and stained 45 min with 0.1% crystal violet. After staining, the plates were washed again three times with MilliQ water, dried, and de-stained with ethanol for 15 min. OD (at 595 nm) of 100 μL samples were measured to analyze biofilm formation (Multiskan FC; Thermo Scientific, China). Statistical differences between mucin concentrations and the control were analyzed using t-test in GraphPad Prism 9.
Influence of mucin on phage-bacterium interaction
First, we explored the effect of mucin supplementation and culture nutrient concentration on S. mutans-phage interaction in TSB medium. Overnight cultures were diluted to 1:10,000, inoculated to 1 × and 0.1 × TSB supplemented with 0.5% mucin in triplicate (control cultures were done without mucin), and incubated at 37°C for 4 h, after which phage M102 was added (105 PFU mL−1). Samplings of 100 μL were made 20 h after the infection, mixed with 10 μL of chloroform, and titrated. Another sampling was made 48 h after the start of the experiment and also titrated. Kruskal–Wallis non-parametric ANOVA was used to analyze the results.
To observe the effect of mucin and phage on bacterial growth and biofilm formation, 2% porcine mucin was added to BHI to reach a final concentration of 0.2%, and it was inoculated with 104 CFU mL−1 of overnight-grown S. mutans. Control cultures were done in BHI without mucin. Half of the cultures were also (simultaneously) inoculated with bacteriophage M102 at multiplicity of infection of 1. From each treatment, 150 μL culture was distributed into a sterile 96-well plate in six replicates, and it was incubated for 2 days at 37°C without shaking.
After incubation, bacterial and phage numbers were sampled from 3 wells per treatment by standard plating methods. Then, the remaining biofilm from all six replicate wells was determined by crystal violet staining as explained earlier. The data were analyzed with non-parametric ANOVA.
To understand the effect of mucosal environment on phage resistance, we co-cultured S. mutans with M102 phage in 0% and 0.2% mucin supplementation (in BHI, 37°C, 200 rpm) in triplicate 5 mL cultures. Samples were plated after 72 h, and bacterial colony forming units were recorded. Phage titers at the end of the experiment were determined with the original S. mutans host. Up to 10 bacterial colonies were picked from each replicate and purified by three rounds of colony-picking and plating. Purified colonies were then inoculated into fresh BHI, grown at 37°C with shaking (200 rpm), and preserved into −80°C freezer stocks with 10% glycerol. Phage resistance of the isolates was determined with a plaque assay using the ancestral M102 phage.
Results
Phage M102 adhesion to mucin-containing plates
A brief survey of the M102 genome (NC_012884) revealed that it has a putative carbohydrate-binding domain in ORF13 (HHPred 25 : Lactobacillus phage J-1 carbohydrate binding module, Probability: 99.95%, E-value: 8.8e-26) as a tail component, which may mediate binding to mucin, or to the bacterial host. We evaluated the ability of M102 to bind in vitro to purified mucin. Such an approach has been applied earlier to evaluate the mucin-binding capacity of different phages.6,10 Despite a trend in having slightly more phages in mucin-containing plates (on average 1.48 more phages on mucin plates), there is no significant difference between the tested conditions (Fig. 1, unpaired t-test, p = 0.1310).
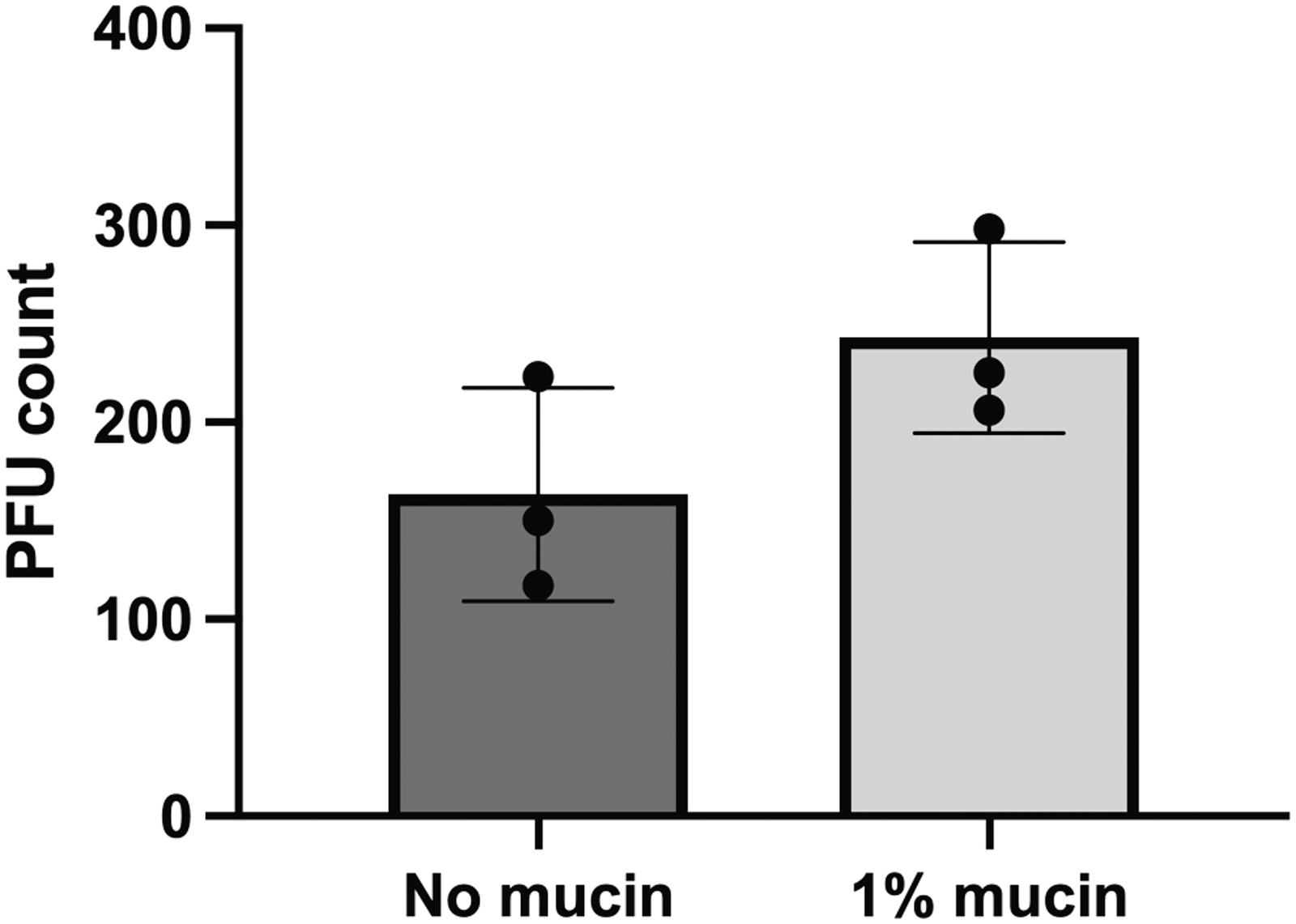
Streptococcus mutans phage M102 adhesion to mucin. Each data point represents an individual replicate that consists of phage plaques count in an agar plate. The mean and standard deviation of each condition is indicated in the graph.
Influence of mucin on S. mutans growth and biofilm formation
We measured the effect of mucin supplementation on S. mutans growth in 5 mL triplicate cultures in TSB alone or supplemented with mucin. Mucin supplementation significantly influenced bacterial growth (Kruskal–Wallis non-parametric ANOVA, test statistic = 47.167, df = 2, p < 0.01), measured as OD of the cultures. In pairwise comparisons, all treatments significantly differed from each other (p-values Bonferroni-corrected) (Fig. 2A).
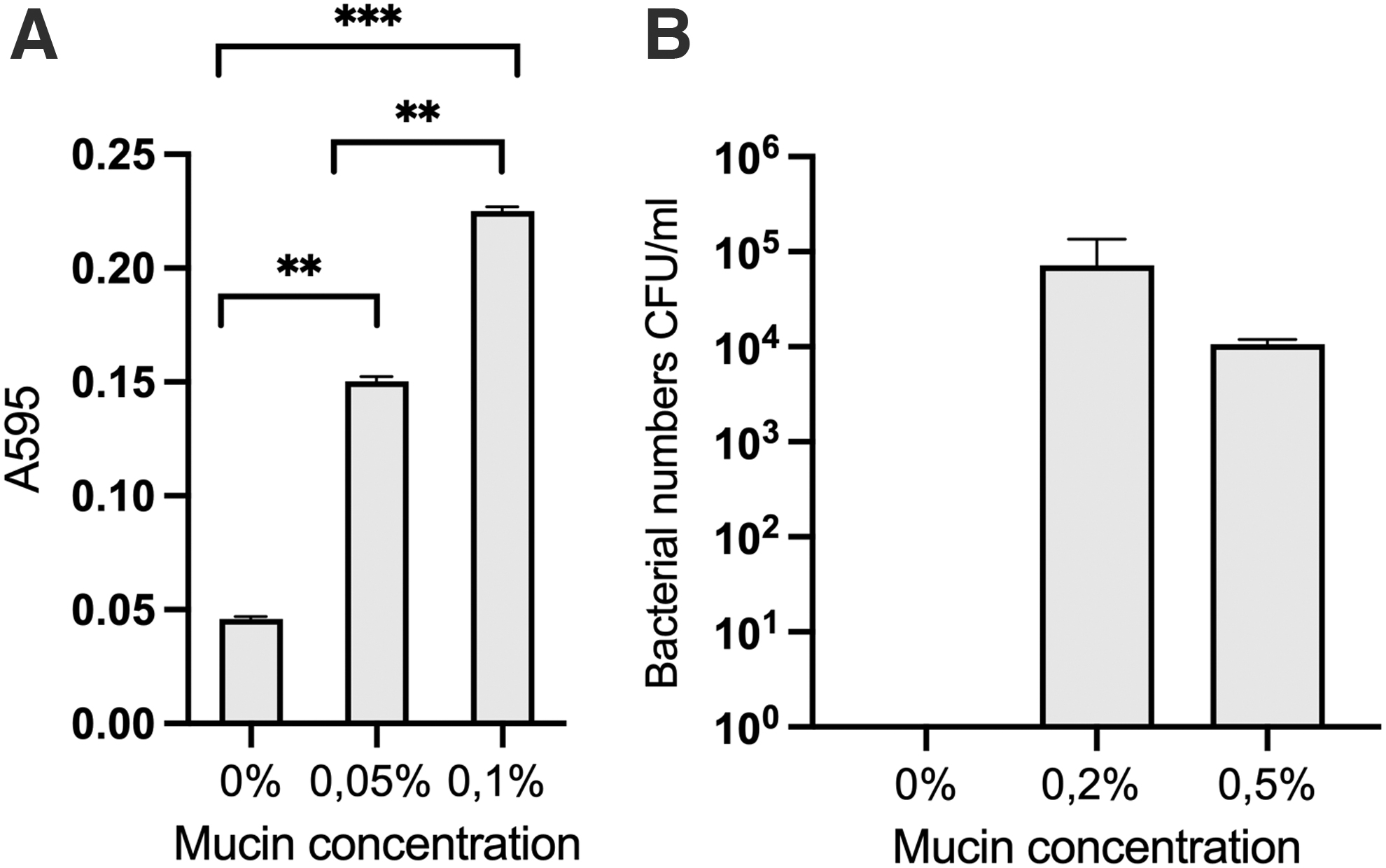
Effect of mucin supplementation on Streptococcus mutans growth.
When the bacteria were inoculated in pure water, there was no bacterial growth observed without mucin supplementation, whereas 0.2% and 0.5% mucin resulted in numbers of 7.2 × 105 and 1.1 × 105 CFU mL−1, respectively (Fig. 2B). There was no statistical difference between these two treatments.
Although it was evident that mucin supplementation benefits S. mutans growth, this was not reflected in biofilm formation (Fig. 3). Mucin presence in water cultures did not lead to biofilm formation, and the same was seen when diluted (0.1 × ) medium was used. In complete (1 × ) medium, biofilm was detected in all conditions, with or without mucin. However, t-tests revealed that mucin addition significantly decreased biofilm formation in 0.1% BHI (p < 0.001 0% vs. 0.2% mucin and 0% vs. 0.5%) and in the 1 × BHI (p < 0.001 for 0% vs. 0.2% mucin and 0% vs. 0.5%). It may be that the presence of nutrients (culture media) favors biofilm formation whereas mucin could serve as an inducer of the planktonic state.
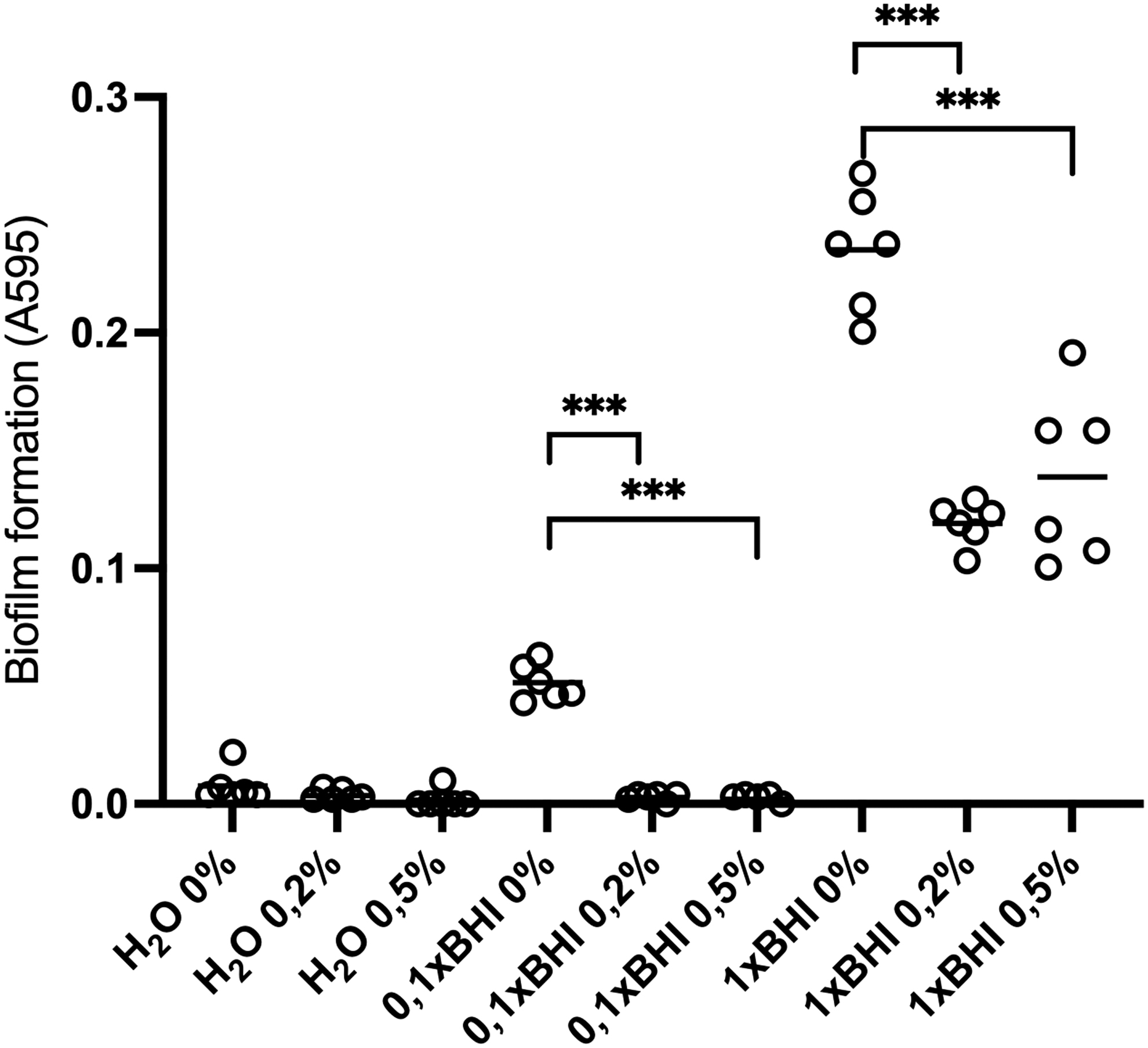
Effect of nutrients and mucin concentration on Streptococcus mutans biofilm formation. Bacteria were cultured without nutrients (H2O), in 0.1 × (10 times diluted) and 1 × (normal) BHI under 0%, 0.2%, and 0.5% mucin supplementation for 48 h. Asterisk indicates statistical significance (***p-value <0.001). BHI, Brain Heart Infusion Broth.
Influence of mucin on phage-bacterium interaction
First, we explored the effect of mucin supplementation on S. mutans-phage interaction in TSB medium. Twenty hours after phage inoculation, there was a significant increase in phage titer when 0.1 × TSB was supplemented with 0.5% mucin (Kruskal–Wallis one-way ANOVA p = 0.0094, Bonferroni-corrected p-value for pairwise comparisons 0.027) (Fig. 4). After 48 h, phage titers were similar in all treatments (data not shown).

Effect of mucin supplementation on M102 phage titers in TSB (1 × : normal TSB, 0.1 × : 10 times diluted TSB). Asterisk indicates statistical significance (*p-value <0.05).
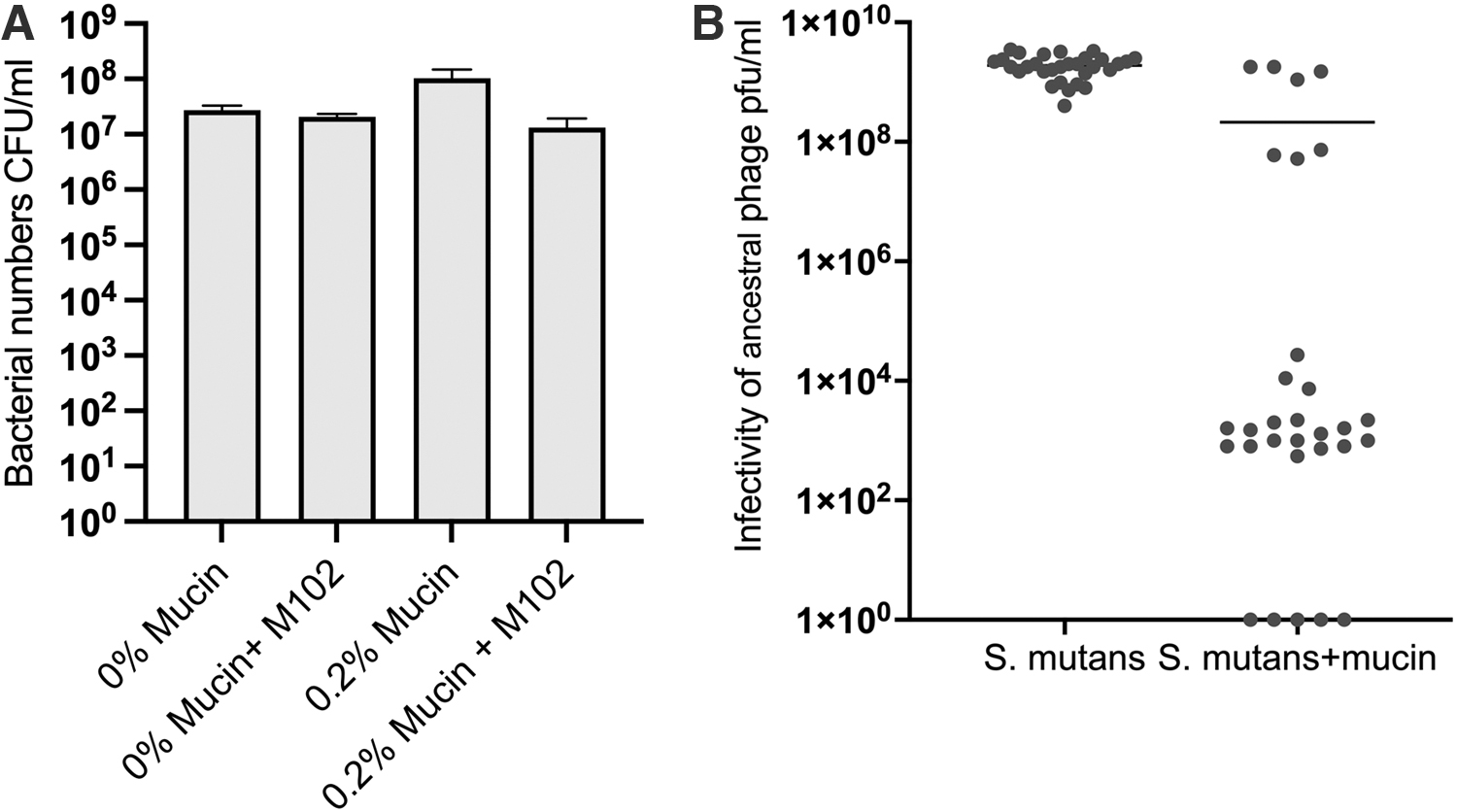
To understand whether the improved phage growth was because of mucin presence or increased bacterial population in the mucin condition, we cultured S. mutans in the presence of phage M102 and mucin for 48 h in 1 × BHI. In the non-infected cultures, bacteria titers did not differ between control and mucin conditions. However, bacterial numbers significantly differed between treatments (Kruskal–Wallis one-way ANOVA; test statistic = 9.359, df = 3, p = 0.025) in the presence of the phage (Fig. 5).

Streptococcus mutans growth and biofilm formation when cultured with 0.2% mucin supplementation and phage M102 for 48 h.
Notably, phage significantly reduced bacterial numbers only when mucin was present. Phage titration of the treatment indicated that phage replicated only in the presence of 0.2% mucin (titer 107 PFU mL−1 vs. 103 PFU mL−1 in treatment without mucin addition). Phage presence also significantly affected biofilm formation, but only when mucin was present (Fig. 5B).
Effect of mucin on phage-bacterium interaction and phage resistance
To understand the effect of mucosal environment on phage resistance, we co-cultured S. mutans with phage M102 in 0% and 0.2% mucin supplementation (in BHI, 37°C, 200 rpm) in triplicate 5 mL cultures. Compared with the previous experiment, we used a longer incubation time to allow phage resistance to evolve. After 72 h, bacterial colony-forming units were recorded and phage titers were determined with the original S. mutans host (Fig. 6).
Phage resistance experiment in mucosal environment. Phage and bacteria were co-cultured for 72 h to allow phage resistance to evolve.
Ten bacterial colonies per replicate were pure cultured from treatments evolving with the phage (and three colonies per replicate from phage-free conditions) and their phage resistance was determined by plaque assays. Under mucin supplementation and phage exposure, 26 of 30 colonies showed a reduced susceptibility to M102 after 72 h. All colonies isolated from the control conditions remained sensitive to phage infection.
Bacterial densities did not statistically differ between experimental conditions but, again, phage replication was detected only in 0.2% mucin (average 1.05 × 107 PFU mL−1). In replicates without mucin, no plaques were detected. Since the phage-only control was also negative for phage plaques, the phage may not have survived the 3-day culture conditions, or the titer was under the detection limit.
Discussion
Streptococcus mutans causes tooth decay, that is, dental caries, a common condition that is hard to treat. The challenges of the increasing antibiotic resistance in general and the susceptibility of the aging population to infections require developing approaches that target bacterial pathogens also in the oral context. This has led to a growing interest toward phage therapy. Phages against S. mutans have already been shown to be potential preventive agents against caries in in vitro and in vivo models, highlighting its relevance as a non-invasive treatment. 26 Therefore, we need to understand how phages infect bacteria and how bacteria resist phage infections in the mucosal environment.
To our knowledge, there has not been prior studies addressing S. mutans phage infections in the presence of mucins. In general, phage-bacterium interactions are most often explored in standard defined growth media, where the possible signals from the vertebrate host targeted by the bacteria are absent. However, in real life, the interactions between pathogenic bacteria and phages often occur on the mucosal surfaces of vertebrate hosts. Our results show that the mucosal environment (achieved by mucin supplementation of growth media) significantly alters S. mutans growth, phage sensitivity, and phage resistance. These results underline the importance of understanding the effect of mucosa on phage-bacterium interactions.
Our results indicate that S. mutans can benefit from mucin as an additional energy source, as mucin supplementation increases bacterial replication at least if added in TSB (Fig. 2), where additional nutrients are available for growth. This is in line with results previously obtained, 7 where the addition of mucin and glucose enhanced S. mutans growth in chemically defined medium. Further, the presence of mucins increases S. mutans survival. 17 This may explain the higher bacterial numbers in mucin-supplemented H2O observed in our study, in contrast to none in replicates without mucin (Fig. 2B). However, it is clear that the interactions between S. mutans and mucins are more complex in vivo. For example, the microbial diversity found in the oral environment can influence S. mutans behaviour and phenotype. Mucins have been shown to influence microbial interactions and support the coexistence of S. mutans and its competitors. 27
Interestingly, mucin decreased S. mutans biofilm formation, which was more dependent on the concentration of nutrients. A previous study using human salivary mucins suggested that mucin decreases S. mutans surface attachment by inducing planktonic growth form. 27 This may be linked directly to sensitivity to phage, as bacteria growing in the planktonic phase are not protected by biofilm, allowing phage infections. Further, the presence of oxygen has been shown to decrease biofilm formation in S. mutans, 19 which may be one of the reasons why the biofilm formation was low in general in our experiments, which were made in aerobic conditions. The influence of mucin under anaerobic conditions is a factor to be considered in future research.
Our results show that mucins significantly influence S. mutans phage survival and replication, exemplifying the role of mucosal surfaces on the phage-bacterium interaction and potentially phage therapy approaches. Studies comparing the density of free and metazoan-associated phages have found that phages tend to concentrate on mucosal surfaces. 10 For example, 108 virus-like particles per mL can be found in the human gut microbiota, but the density is higher in the gut mucosa (109 phages per mL). 28
This can be explained by the affinity of some phages toward mucins: Some tailed dsDNA phages have (structural) protein domains that mediate their adherence to mucin glycoproteins found in the mucosal surfaces. A bioinformatic analysis of 246 dsDNA tailed-phage genomes suggested that roughly 25% have proteins with Ig-like folds, all related to the viral structure. 29 More recently, also other domains30,31 have been found and suggested to facilitate phage binding to mucosa. Phage M102 22 has a putative carbohydrate-binding domain in ORF13 that might mediate the phage-mucus interaction, but the role of this domain remains unknown so far. Nevertheless, although M102 mucin binding in vitro was not significant, the ratio between plaque count in mucin divided by plaque count in control was 1.48.
Since it has been shown that phage-mucin interactions are transient but constant, 20 even a slightly positive ratio allied to the numerous interactions between the phage and mucins may still play a role in maintaining phages in the mucosa. Further, although the phage titers in mucin-free cultures were below detection limit, it is also possible that the presence of mucins stabilizes phage particles, influencing their survival in the mucosal environment.
Although phage-host interactions in S. mutans have been previously explored,26,32,33 there is a lack of knowledge on how the mucosal environment influences bacterial susceptibility to phage. We observed that mucin has a major role for phage susceptibility: Mucin supplementation caused a 4-log increase in phage titer of S. mutans. Although the exact mechanisms for this phenomenon are unknown, it is possible that phages benefit from higher bacterial replication or that they exploit their changed metabolic state.
It is also possible that the upregulation of bacterial virulence factors (related to cell wall structures, secretion, metabolism, or motility) in the mucosal environment increases the availability of phage receptors. This could explain the previously observed increased virulence of pathogens in the mucosal environment.4–6 In our experiments, it seems evident that the choice of medium influences the impact of mucin on bacterial growth and phage sensitivity. In TSB, which is a less rich medium than BHI, phage replication was improved only in the diluted medium (Fig. 4) whereas in BHI phage replication (and a drop in bacterial population density) was observed only under mucin supplementation.
However, in BHI, mucin supplementation did not cause any differences in bacterial numbers in the absence of phage (Fig. 5). This suggests that mucin-induced expressional changes are more likely to cause sensitivity to phage infection. Consequently, phage resistance was elicited only under mucin supplementation when the bacteria were co-cultured with phage for 72 h. This is seen as a comparable bacterial population size in phage-treated and control culture in Figure 6A. Although phage resistance in S. mutans has been observed in previous studies without mucin,32,33 it remains to be studied how the mucosal environment influences the evolution of resistance in S. mutans, for example, CRISPR-Cas. 18
Phage resistance often leads to decreased virulence capacity in bacteria,34,35 but the role of resistance in mucosal environment is not yet completely understood. Although mucin seems to be connected with increased suscpetibility to phage infection, in some cases, mucins may also impair phage infectivity.6,17 Such phages are probably more efficient in targeting pathogens outside the mucosa. Differences in the capacity to adhere to mucus (or to replicate in mucosal environment) could be considered as one additional phenotypic feature of phage-bacterium interaction, which could be exploited in building phage cocktails for therapy purposes. A combination of phages with different strategies (mucin-binding or not) may produce efficient treatment and prevention tools that target pathogens both from the infection site and during transmission.
The mucin concentrations used in these experiments are relevant and comparable to what is observed in the body, although mucin species and concentrations vary depending on body site and in response to physical conditions, disease, and age.36–39 For example, in the gastric environment, the concentration of the mucins is ∼5%, 21 but the density of the mucosal layer is not consistent, as the mucin concentration is highest near the cell surface and less dense at the outer layer. The oral mucosal environment contains several different mucins; secreted salivary mucins MUC5B, MUC7, and MUC19; and membrane-associated mucins MUC1 and MUC4. 40
The salivary mucin levels are generally higher in patients with dental caries, for example, with MUC5B concentration varying from 0.06 to 2.34 ng mL−1. 38 Although the exact mucin species composition of the porcine mucin used in our experiments is not known, it mainly consists of MUC2. It, thus, seems that S. mutans may not have a specific affinity toward certain mucins as observed in some other bacteria. 40 Further, it is important to note that porcine mucin has been previously shown to be relevant for phage-bacterium studies in other species also, eliciting a similar increase in phage replication. 6
Combined with previous research, these results highlight the effect of mucosal environment on phage-bacterium interactions, and they emphasize the ecological relevance of microbial interactions in the eukaryotic mucosa, with implications for the outcome and prevention of diseases.
Footnotes
Authors' Contributions
Conceptualization: G.M.d.F.A., L.-R.S. Methodology: G.M.d.F.A., N.R., L.-R.S. Data curation: G.M.d.F.A., N.R., L.-R.S. Writing—original draft preparation: L.-R.S. Writing—review and editing: G.M.d.F.A., N.R., L.-R.S. Visualization: G.M.d.F.A., L.-R.S. Supervision, project administration, and funding acquisition: L.-R.S.
Author Disclosure Statement
G.M.d.F.A., L.-R.S. and University of Jyväskylä have patented the commercial use of mucin in a patent titled “Improved methods and culture media for production, quantification and isolation of bacteriophages” (FI20185086, PCT/FI2019/050073).
Funding Information
This study was funded by the Academy of Finland (no. 314939 for L.-R.S.).