
Abstract
Background:
Escherichia coli O157:H7, causing a severe public health concern, is currently an emerging food-borne pathogen. Bacteriophages (phages), to this effect, can provide alternative measures for their control. This study aimed at identifying and characterizing environmental Escherichia coli O157:H7, the phages infecting them, and some selected pathogenic bacteria.
Materials and Methods:
The bacteria and phages were isolated from six (6) different locations within Jos, Nigeria. The bacterial strains were identified using the Bruker matrix-assisted laser desorption ionization-time of flight mass spectrometry system, and their antibiotic susceptibility profiles were determined. The phages were screened against Escherichia coli O157:H7 and some selected pathogenic bacteria and examined for their physiological and morphological characteristics.
Results:
The mean bacterial viable counts indicated that although Angwa Rukuba_1 had the highest count of 7.69 ± 8.89 log10 CFU/mL, Student Village Hostel_1 had the lowest count of 7.52 ± 2.65 log10 CFU/mL. Bacillus spp. had the highest occurrence (30%), followed by Escherichia coli (28%), Pseudomonas aeruginosa (26%), Klebsiella spp. (8%), Proteus spp. (4%), and Staphylococcus aureus (4%). The results indicated that all the isolates were resistant to ceftazidime, whereas others were either susceptible or resistant to the eight common antibiotics used. The four bacteriophages designated as E. coli phage Anayo (ECPA)_1, ECPA_2, ECPA_3, and ECPA_4 had mean titers ranging from 9.80 ± 38.55 to 11.85 ± 13.45 log10 PFU/mL. The phages displayed broad activity against Escherichia coli O157:H7 and some selected pathogenic bacteria. The phages were relatively stable over a wide range of temperatures (45–55°C) for varying time intervals, NaCl concentrations (0.5–15%), chloroform (10%), and pH values (2–9) after 24 h of exposure.
Conclusions:
This study indicated that the Escherichia phages have activity on Escherichia coli O157:H7 strains as well as on other bacterial pathogens used. The experimental phages obtained in this study can be exploited in the formulation of phage cocktails for alternative therapy and bio-control of Escherichia coli O157:H7 because of their lytic activity and stability to different storage conditions.
Introduction
Escherichia coli is a commensal microorganism present in the gastrointestinal tract and feces of humans and animals and was first isolated by Escherich over a century ago.1–3 Its pathogenicity was first established in 1975, when Escherichia coli O157:H7 was found in a patient with bloody diarrhea in California. 4 In addition, this bacterial strain was the primary cause of a foodborne (ground beef) outbreak of disease. Subsequently, verocytotoxin-producing E. coli, Shiga-toxin-producing E. coli, and enterohemorrhagic E. coli (EHEC) were characterized and reported across the world because of their similarity to toxins produced by Shigella dysenteriae.1,5–7
The pathogenicity of EHEC appears to be associated with some virulence factors, including the production of several cytotoxins, their tolerance to some chemical and physical treatments, the severity of the illness, low-dose infectiousness, and widespread diffusion.8,9 Consumption of undercooked meat and ground beef is the most common source of outbreaks of this foodborne pathogen. 10 Recently, a diarrheagenic strain, enteroaggregative E. coli, has been reported.
In the current scenario, serious medical and social problems result from the increasing antibiotic resistance of bacterial strains. 11 As a result, pharmaceutical industries are withdrawing from research on new antibiotics and exploring the current system of microbial control, such as the use of probiotics, bacteriophages, or phage-derived products, as alternative therapy.12–14
A bacterial virus called bacteriophage is considered to offer a great advantage over antibiotics. The normal gut microflora is not affected because bacteriophages target only the pathogens of interest. They kill bacteria as they self-replicate in the bacterial host. 15 Activities against zoonotic pathogens in live animals, bacteriophages have been used.16–19
As recently investigated, the application of bacteriophages as a food safety intervention with a few commercial preparations has been approved and marketed. Bacteriophages are frequently employed in high doses to inactivate foodborne pathogens such as Escherichia coli O157:H7, Salmonella, Listeria, and Campylobacter in various foods.
Phages have been used to control specific bacteria at the pre-harvest and post-harvest stages of food production and storage in production facilities.20,21 The application of bacteriophage is dependent on the biological properties of the phages, which affect their activity in the biological system.
Due to the high degrees of phenotypic diversity within populations of both phages and bacteria, the application of phages to control a certain bacterial pathogen is complicated. 14 Individual strains of a pathogen may be resistant to or more or less susceptible to different co-occurring phages. It is necessary to understand in detail the phage-host interaction, which is affected by both biological and physical factors, before therapeutic application. 22 The main physical factors affecting phage adsorption and bacterial growth are temperature and pH, whereas the biological aspect is related to bacterial resistance.
This study is, therefore, aimed at identifying and characterizing environmental Escherichia coli O157:H7 and bacteriophages infecting them, as well as some selected pathogenic bacteria from Jos metropolis. The Jos Plateau is home to a sizable population of Fulani pastoralists and is, therefore, a significant cattle-producing region in Nigeria.
The specific objectives of this research will be focused on the isolation, identification, and screening of environmental bacteria isolated from effluent water and soil for use as permissive hosts; determining the antibiotic susceptibility profile of the bacterial isolates; isolation, purification, propagation, and determining the phenotypic characteristics of isolated bacteriophages from effluent water and soil; determining the stability of the isolated phages under varying storage conditions; and determining the lytic activity (host range) of the isolated phages against Escherichia coli O157:H7 and other common pathogens.
Materials and Methods
Sample collection
Under aseptic conditions, a total of 12 samples, including wastewater and soil (2 samples for each location), were obtained from 6 different locations (Student Village Hostel 1 and 2, Old Jos University Teaching Hospital, JUTH 1 and 2, and Angwa Rukuba 1 and 2), within Jos North Metropolis, Plateau State, Nigeria. Latitude and longitudes of their various locations were noted as follows: latitude: 9.96565, 9.96571, 9.918695, 9.918323, 9.93922, and 9.934003; longitude: 8.87116, 8.87128, 8.890219, 8.890219, 8.909185, and 8.908757 respectively.
This was accomplished by using 50 mL sterile vials with cover tops. The containers were immediately disinfected with 70% ethanol at the point of collection, labeled, kept in a super cool flask, and transported for analysis to the Microbiology Laboratory Research Unit, Africa Centre of Excellence in Phytomedicine Research and Development (ACEPRD) University of Jos, Jos, Plateau State, Nigeria (Supplementary Figs. S1 and S2).
Bacterial isolation
A total of 10 mL of each sample (after mixing the wastewater and sand and allowing it to decant in a conical flask) was diluted in 90 mL of sterile 0.9% NaCl normal saline and homogenized. 23 A 100 μL of the fourth and fifth diluents of the samples were then inoculated on Eosin Methylene Blue Agar agar plates for enteric bacteria isolation, MacConkey agar plates for both lactose and non-lactose fermenters, and Sorbitol-MacConkey for Escherichia coli O157:H7.
For 24 h, all of the bacteria plates were kept at 37°C. The total viable count was calculated by utilizing the spread plate technique on nutrient agar and counting the colonies that formed after 24 h of incubation at 37°C. 24
Identification of bacterial isolates
Gram staining, Motility, Hemolysis test, Methyl-Red test, Voges–Proskauer test, Catalase test, Coagulase test, Oxidase test, Indole test, Citrate-utilization test, Urease test, Fermentation test, and Triple Sugar Iron (TSI) test were used to identify the bacterial isolates.25–27
Bruker matrix-assisted laser desorption ionization-time of flight mass spectrometry identification
The isolates were cultured onto chromogenic culture medium (CHROMagar) agar plates and incubated at 37°C for 24 and 48 h. Identifications were performed using Microflex Bruker Daltonics/BioTyper™ version 2.0 after 24 and 48 h of incubation, according to the manufacturer's instructions. Each isolate was identified in duplicate, and the best identification score was used. Each of the bacteria isolates tested was spotted twice on the same target slide.
According to the manufacturer's suggested settings, with automated collecting spectra, measurement was performed on a MALDI Biotyper. MALDI Biotyper automation control and Bruker Biotyper 3.1 software containing the library number 3995 enabled the captured spectra to be analyzed. For each run, to calibrate the instrument and validate the run, a standard (E. coli test standard, Bruker Daltonics) was included.
As the manufacturer recommended: a score ≥2.000 indicated species-level identification, a score between 1.700 and 1.999 indicated genus-level identification, and a score <1.700 was interpreted as no identification was used as identification criteria. The subsequent deposit was done with the on-slide formic acid extraction technique, followed by the in-tube formic acid extraction technique if a good identification was not obtained with the direct colony technique.28,29
Antibiotics susceptibility profile
The antibiotic susceptibility profile of the test organism and the environmental isolates were determined using the standard Kirby-Bauer disk diffusion method. The antibiotics used have the following disk concentrations: Ceftazidime (10 μg), Cefuroxime (30 μg), Gentamicin (10 μg), Ciprofloxacin (10 μg), Nitrofurantoin (300 μg), Ampicillin (10 μg), Ofloxacin (10 μg), and Augmentin (30 μg).
Bacterial culture suspension equivalents of 0.5 tube McFarland turbidity standards were spread on Muller–Hinton agar plates using sterile swabs and incubated aerobically at 37°C for 24 h; then, the zone of inhibition diameters around the antibiotic disks was measured.
The obtained results were compared with the standard performance chart for antimicrobials disk susceptibility testing provided by the Clinical and Laboratory Standards Institute (CLSI), and the frequencies of sensitivity and resistance were recorded. According to the criteria recommended by CLSI, the results were expressed as susceptible or resistant. 30
Phage isolation
Direct phage isolation from water/sewage samples
To remove debris and other undissolved particles, 50 mL of water/sewage was centrifuged at 10,000 g for 10 min. Parts A and B of the supernatants were separated (saved portion B for enrichment). A piece of Portion A was separated into two parts, A1 and A2. To kill any bacteria that had survived, 0.5 mL chloroform was added to part A1. It was kept at 4°C for 30 min. It was centrifuged at 21,000 g for 10 min after 1 h and filtered using 0.45 μm filters. A2 was centrifuged at 21,000 g for 10 min before being filtered using 0.45 μm filters. Portion B was centrifuged for 10 min at 21,000 g and filtered using a 0.45 μm filter.
Phage isolation from the enrichment of water/sewage samples
In a conical flask, 10 mL filtered sample B, 10 mL double-strength Luria-Bertani (LB) broth, 40 μL, 1 M CaCl2, and 100 μL overnight broth culture of bacteria (0.5 McFarland standard) were added. It was incubated at 37°C for 48 h with 50 rpm shaking. After 24 h, 5 mL of the sample (B1) was taken and centrifuged for 10 min at 4200 rpm. The supernatant was filtered through 0.45 μm filters after centrifugation at 21,000 g for 10 min. The residual fraction was centrifuged and filtered after 48 h (Sample B2).
Phage isolation from soils
A conical flask was filled with the following: 1 g of soil/mud, 5 mL LB broth, gently inverted at 20 rpm for 30 min, and then incubated at 4°C for 30 min. The sample was centrifuged at 4200 rpm for 10 min. The supernatants were split into two groups: C and D. Portion C was centrifuged for 10 min at 21,000 g and then filtered using 0.45 μm filters (direct soil phage isolation). The following was added to a flask: 100 μL overnight bacterial culture, 5 mL double-strength nutrient broth supplemented with CaCl2, and 5 mL soil/mud centrifuged supernatant D.
It was incubated at 37°C for 48 h with 50 rpm shaking. Five milliliters of the sample (D1) was collected after 24 h and centrifuged at 15,000 g for 10 min before filtering through 0.45 μm filters. The residual fraction was centrifuged and filtered after 48 h (Sample D2).
Phage detection from extracted supernatants
Lawns of the bacterial isolates were created after the overlays had been set; a part of the agar plate was spotted with 10 μL of the filtered samples A1, A2, B1, B2, C, D1, and D2. They were allowed to dry before being incubated at 37°C overnight. On the bacterial lawns the next day, lysis/clearance zones were visible. The lysed zones were scraped out and transferred to 1 mL sodium-magnesium (SM) buffer in a microfuge tube, where they were mixed by inversion. The vortex was avoided, and the mixture was kept at 4°C for 30 min. They were centrifuged for 10 min at 15,000 g and then filtered (Supplementary Fig. S3).31,32
Plaque purification
Plaque assay
Day 1
A 96-well micro-titer sterile plate was used to perform a 10-fold serial dilution of 10–1 to 10–8 of the lysed zone filtrates from two B1 and D1 samples in cold SM buffer. The nutrient agar plate for each tube was labeled with a number from 10–1 to 10–8. In a sterile microfuge tube labeled 10–1 to 10–8, a 100 μL overnight standardized culture of (Escherichia coli O157:H7 host bacteria 0.5 McFarland standard) was added. A 100 μL sample was taken from the 10–1 serial dilution of the filtrates and pipetted up and down many times into a microfuge tube containing 100 μL of overnight culture labeled 10–1.
All 200 μL was dispensed into a 3 mL overlay agar (for 90 mm Petri dishes) in a tube and mixed by inversion. The slurry was dispensed and allowed to be set onto the suitably labeled nutrient agar plate. This was done for each serial dilution and incubated aerobically at 37°C overnight.
Days 2, 3, 4
As each plate was visually evaluated, the one with distinctive distinct plaques was chosen. A microfuge tube containing 500 μL of diluent (SM buffer) was produced for each plaque morphology on the plate. From the chosen plate, a pipetman with a pipette tip was chosen, and the overlay agar was gently penetrated in the middle of the plaque. The prepared tube was instantly filled with the cored plaque.
This was repeated three times using the most recently cored plaque in 500 μL of diluent instead of the enhanced phage lysate each time (SM buffer). Plaques with the same plaque morphology as the parent plaque (i.e., the previous plaque selection) were selected with care (Supplementary Fig. S4).
Phage propagation
A 2 mL Escherichia coli O157:H7 broth (0.5 McFarland standard), 20 μL of phage lysate, 10 μL of nutrient broth, and 40 μL of 1 mM CaCl2 were added to a sterile conical flask in a shaker incubator at 180 rpm for 24 h at 37°C and the PFU/mL was calculated.31,32
Physicochemical characterization of the isolated bacteriophages
Thermal stability test
A total of 64 nutrient agar plates were prepared and incubated overnight for the sterility test. Each of the four temperatures (45°C, 55°C, 65°C, and 75°C) comprises 16 plates for each of the four phages isolated. Ninety microliters of SM buffer was added to sterile test tubes. They were heated to 45°C, 55°C, 65°C, and 75°C in a water bath and kept at ±0.05°C.
The water level was kept 2 cm above the treatment using a stirrer. A 100 μL aliquot of phages was added to the 900 μL and boiled for 60 min for each test tube of the relevant temperature. A serial dilution to 10–8 was prepared using a sterile microtiter plate and SM buffer. Every 15 min, 10 μL of the sample or treatment phages were pipetted into a 90 μL micro-titer plate and diluted. The host bacteria were seeded onto the overlay, which was then dispensed over a nutrient agar plate with the diluent diluted to 10–8. It was allowed to dry before being incubated at 37°C for an overnight period. This method was repeated for the individual phages.
pH stability test
For the sterility test, 24 agar plates were prepared and incubated overnight. Nutrient agar overlays were standardized using 1 M NaOH and HCL to provide pH values of 1, 2, 5, 7, 9, and 11 using SM buffer. Nine hundred microliters of each pH and 100 μL of phages were prepared and cultured for 24 h at 37°C.
After that, a serial dilution (10-folds) was performed, with 90 μL of SM buffer and 10 μL of phage lysate. Ten microliters of each diluent was inoculated into a solidified overlay agar and incubated for 24 h at 37°C.
Salinity stability
SM buffer with NaCl concentrations of 0.5%, 5%, 10%, and 15% was made with negative control of SM buffer with no salt. For the four phages, a total of 20 agar base plates were made, each with a different proportion. As indicated earlier, a 10-fold serial dilution was made. For 24 h, 90 μL of SM buffer + NaCl and 10 μL of individual phages was incubated at room temperature. A test organism-seeded overlay was made and left to dry. They were spotted with 10 μL of the diluents and allowed to dry before being inverted at 37°C for 24 h.
Chloroform stability test
There were eight nutrient agar basic plates made. The four phages treated with chloroform had four plates, whereas the four phages not treated with chloroform had four as negative controls. An overlay was prepared, Escherichia coli O157:H7 (0.5 McFarland standard) was prepared, and 10% v/v chloroform was prepared using SM buffer.
In a sterile test tube, an equal proportion of 500 μL lysate and 500 μL chloroform was combined and allowed to stand for 60 min with mild shaking at intervals. They were centrifuged for 10 min at 10,000 g. As previously indicated, a serial dilution to 10–8 was performed. The test organism-seeded overlay was prepared and allowed to solidify. Individual diluents were spotted (10 μL) and allowed to dry aseptically before being inverted for 24 h at 37°C.33–35
Host range of the bacteriophages
Base plates of nutrient agar were made. Overlay agar containing individual organisms was dispensed on the base agar and allowed to gel. The overlay was spotted with 10 μL of each of the four phages. They were dried, inverted, and incubated at 37°C for 24 h.36,37
Statistical analysis
The descriptive analysis was completed using SPSS software version 21. Duncan's multiple-range test was used to determine the analysis of variance (JMP version 12 software; SAS Institute, Cary, NC, USA). To detect significant differences, p-values <0.05 (p = 0.05) were calculated.
Results
Total viable count for environmental isolates
Figure 1 reveals that the mean microbial load of the environmental isolates differs significantly, with Angwa Rukuba_1 having a mean value of 7.698.89 log10 CFU/mL, which is significantly different from Angwa Rukuba_2 (7.573.61 log10 CFU/mL) and Student Village Hostel_1 (7.522.65 log10 CFU/mL). Student Village Hostel_2 (7.646.35 log10 CFU/mL) and Old JUTH_1 (7.645.20 log10 CFU/mL) had no significant differences in mean values; however, Old JUTH_2 (7.633.79 log10 CFU/mL) has a marginally significant difference in mean values.

Total viable count for environmental isolates. Values shown means of triplicate measurement ± standard deviation. Figures with different superscripts in the columns are significantly different (P < 0.05).
Cultural identification of environmental bacteria isolates and screening for permissive host
Table 1 shows that the isolates were distinguished based on morphological, microscopic, and biochemical features. The isolates differ in terms of Gram reaction, cell morphology, motility, hemolysis, lactose fermentation, citrate test, urease, catalase, and the TSI agar test.
Cultural Identification of Environmental Bacteria Isolates and Screening for Permissive Host
Keys: −, negative; +, positive; A, acidic; EIA, environmental isolate Anayo; K, alkaline.
Three isolates were found to be permissive hosts for the phages: EIA 18 (E. coli), EIA 19 (Bacillus spp.), and EIA 26 (Pseudomonas spp.).
Matrix-assisted laser desorption/ionization-time of flight mass spectrometry identification
In Table 2, matrix-assisted laser desorption ionization-time of flight (MALDI-TOF) mass spectrometry (MS) was used to confirm the environmental isolates found in Table 4. The probability of the first and second isolations was calculated using the phenotypic data on CHROM agar.
Matrix-Assisted Laser Desorption/Ionization-Time of Flight Mass Spectrometry Identification
Keys: −, not good; +, slight good; ++, moderate good; +++, highly good.
Percentage occurrence of environmental isolates
Figure 2 shows that Bacillus spp. has the highest prevalence (30%), followed by E. coli (28%), and Pseudomonas aeruginosa (26%). Klebsiella spp., Proteus spp., and Staphylococcus aureus had the fewest occurrences.
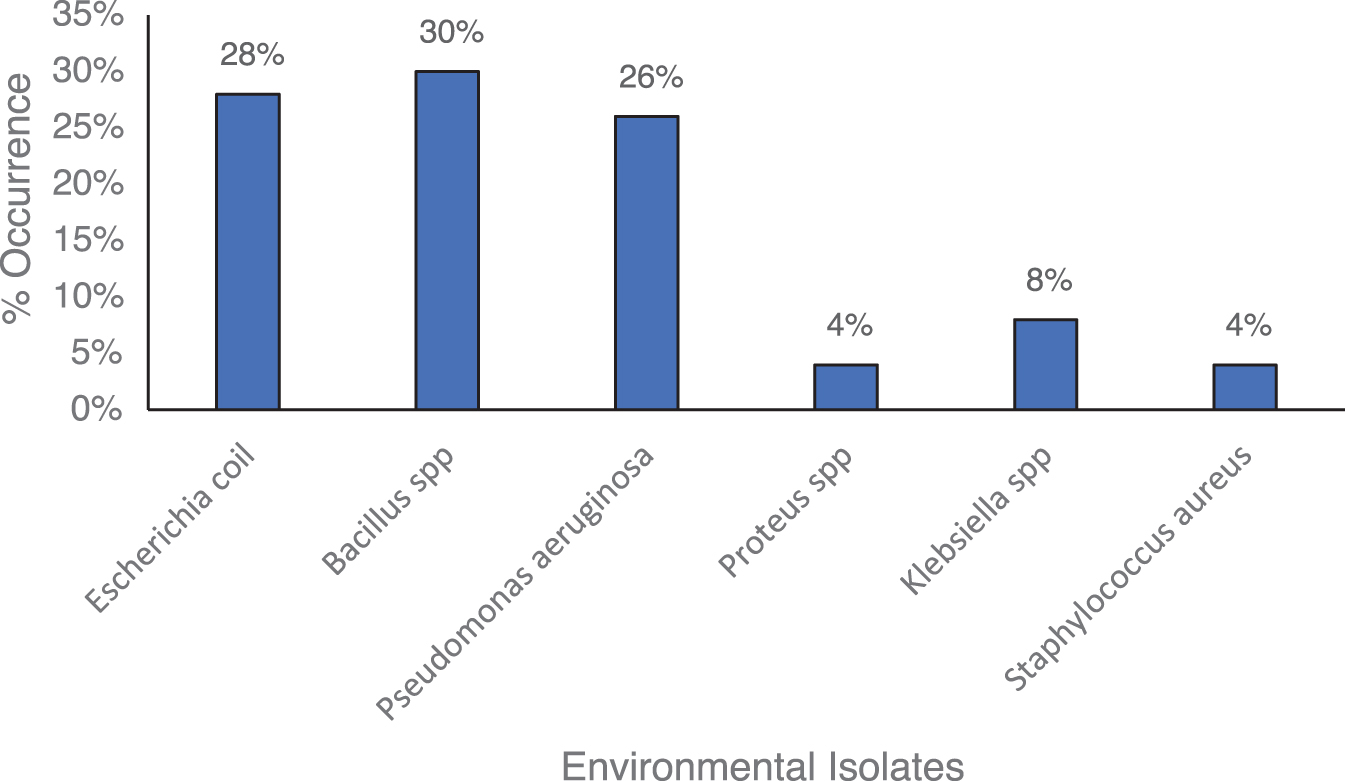
Percentage occurrence of environmental isolates.
Antibiotics susceptibility profile of the test organism and environmental isolates
Table 3 illustrates the test organisms and environmental isolates' antibiotic susceptibility profiles. The results demonstrate that all of the isolates were Ceftazidime-resistant, with low resistance to Cefuroxime, Ampicillin, and Augmentin but high susceptibility to Gentamicin, Ciprofloxacin, Nitrofurantoin, and Ofloxacin.
Antibiotics Susceptibility Profile of the Test Organism and Environmental Isolates
Amp, Ampicillin; Aug, Augmentin; Caz, Ceftazidime; Cpr, Ciprofloxacin; Crx, Cefuroxime; Gen, Gentamicin; Nit, Nitrofurantoin; Ofl, Ofloxacin.
Distribution of bacteriophages in relation to sample location
In Table 4, only four of the six sample locations, Student Village Hostel 1, Student Village Hostel 2, Old JUTH 1, and Old JUTH 2, had bacteriophages designated E. coli phage Anayo (ECPA) 1, ECPA 2, ECPA 3, and ECPA 4 present.
Distribution of Bacteriophages in Relation to Sample Location
−, absent; +, present.
Bacteriophage titer
Figure 3 indicates the phage's average titers. It shows that the titers of phages ECPA_1 and ECPA_4 are significantly different, with 11.582.65 and 10.0435.64 log10 PFU/mL, respectively, whereas ECPA_2 and ECPA_3 have a titer of 11.8513.45 and 9.8038.55 log10 PFU/mL, respectively, and are not statistically different.
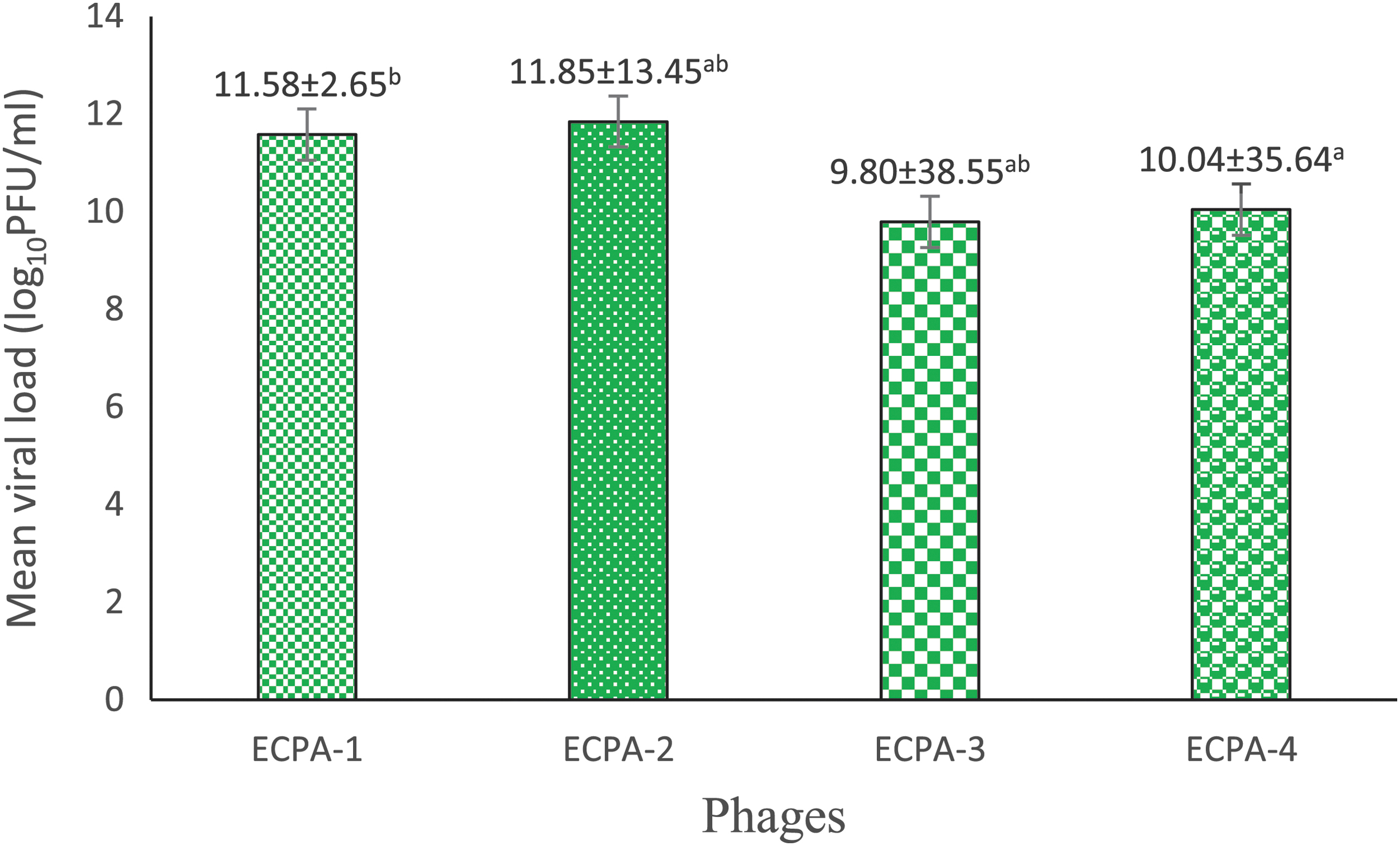
Bacteriophage titer. ECPA, E. coli phage Anayo. Values shown means of triplicate measurement ± standard deviation. Figures with different superscripts in the columns are significantly different (P < 0.05).
Thermal stability test
Figure 4 demonstrates the thermal stability of the phages at 45°C, 55°C, 65°C, and 75°C while altering the time (minutes) at 15, 30, 45, and 60 min. At 45°C, phage ECPA_1 shows a slight decrease in titer from the working concentration of 12.58 ± 1.00 to 10.11 ± 1.00, 9.11 ± 1.00, and 4.11 ± 1.00 log10 PFU/mL as the time increased from 15, 30, and 45 min, respectively, whereas at 60 min, there was no visible plaque detected.
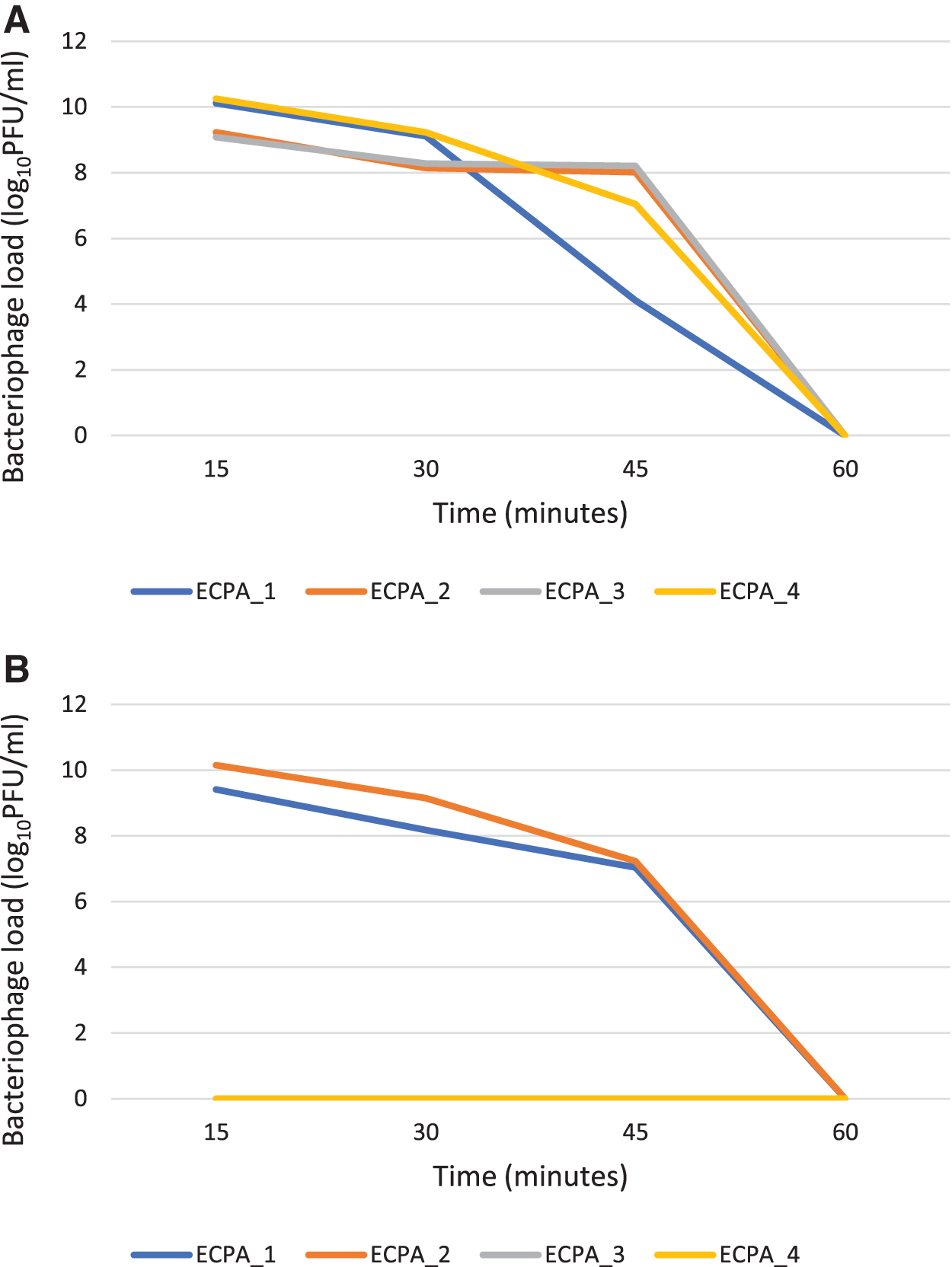
Phage ECPA_2 shows a sharp decrease in titer from the working concentration of 12.58 to 9.23 ± 2.00, 8.15 ± 1.00, and 8.02 ± 1.00 log10 PFU/mL at 15, 30, and 45 min, respectively, whereas at 60 min, there was no visible plaque detected. Phage ECPA_3 shows a sharp decrease in titer from the working concentration of 12.58 to 9.08 ± 2.00, 8.28 ± 2.65, and 8.20 ± 3.00 log10 PFU/mL at 15, 30, and 45 min, respectively, whereas at 60 min, there was no visible plaque recorded.
Phage ECPA_4 shows a decrease in the phage titer as the time increased from the working concentration of 12.58 to 10.26 ± 1.00, 9.23 ± 1.00, and 7.04 ± 1.00 log10 PFU/mL at 15, 30, and 45 min, respectively, whereas at 60 min, there was no visible plaque observed.
At 55°C, ECPA_1 shows a sharp decrease in the titer from the working concentration of 12.58 to 9.41 ± 1.00, 8.18 ± 1.00, and 7.04 ± 1.00 log10 PFU/mL at 15, 30, and 45 min, respectively, having no visible plaque at 60 min. ECPA_2 shows a decrease in the phage titer from the working concentration of 12.58 to 10.15 ± 1.00, 9.15 ± 1.00, and 7.23 ± 2.00 log10 PFU/mL at 15, 30, and 45 min, respectively, whereas at 60 min, there was no visible plaque observed. At 15, 30, 45, and 60 min, ECPA_3, and ECPA_4 exhibit no apparent plaque. At 65°C and 75°C, there are no visible plaques observed for phages ECPA_1, ECPA_2, ECPA_3, and ECPA_4 at 15, 30, 45, and 60 min.
pH stability test
Figure 5 shows the pH stability of the phages. At pH 1, 2, and 11, ECPA_1 reveals no apparent plaque. There is a sharp decrease in the phage titer from the working concentration of 10.79 to 5.40 ± 3.00, 5.64 ± 1.00, and 3.447158 ± 8.89 log10 PFU/mL at pH 5, 7, and 9, respectively. ECPA_2 shows a decrease from the working concentration of 10.79 to 5.16 ± 2.00 log10 PFU/mL at pH 7 whereas there is no visible plaque at pH 1, 2, 5, 9, and 11.
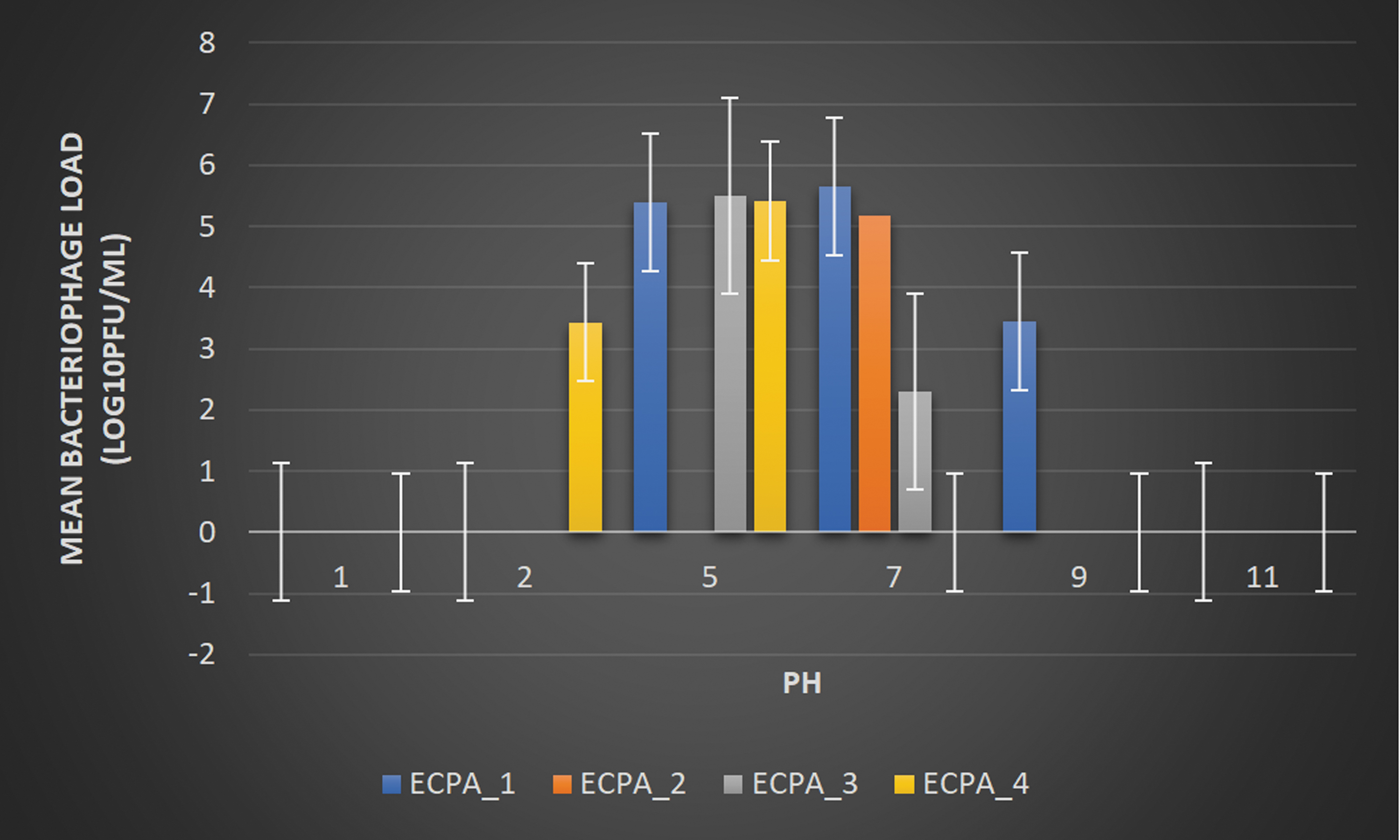
pH Stability test. Working concentration = 10.79 log10 PFU/mL.
ECPA_3 shows a sharp decrease in titer from the working concentration of 10.79 to 5.51 ± 6.56 and 2.30 ± 1.00 log10 PFU/mL at pH 5 and 7; at pH 1, 2, 9, and 11, there is no apparent development, respectively. ECPA_4 decreases from the working concentration of 10.79 to 3.43 ± 4.36, and 5.41 ± 2.00 log10 PFU/mL at pH 2 and 5, respectively, whereas there is no visible plaque at pH 1, 7, 9, and 11.
Salinity stability test
Figure 6 demonstrates the phage's stability at various NaCl concentrations. ECPA_1 shows a decrease from the working concentration of 10.51 to 10.51 ± 1.00, 10.40 ± 1.00, 10.18 ± 3.00, and 8.08 ± 2.65 log10 PFU/mL with different NaCl concentrations of 0.5, 5.0, 10.0, and 15.0, respectively. ECPA_2 shows a decrease in the phage titer from the working concentration of 9.08 to 9.08 ± 2.65, 10.20 ± 1.73, 10.256 ± 1.00, and 8.00 ± 2.00 log10 PFU/mL at NaCl concentrations of 0.5, 5.0, 10.0, and 15.0, respectively.
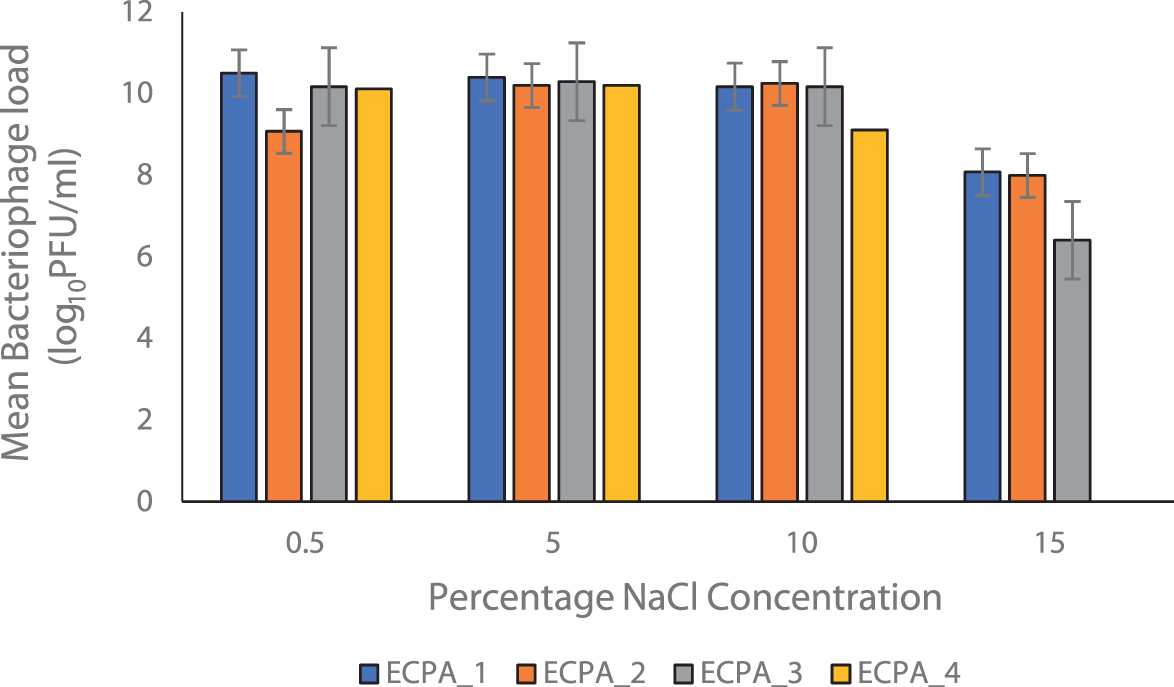
Salinity stability test. Working concentrations; ECPA_1 = 10.51 log10 PFU/mL, ECPA_2 = 9.08 log10 PFU/mL, ECPA_3 = 10.26 log10 PFU/mL, and ECPA_4 = 8.08 log10 PFU/mL.
ECPA_3 shows a decrease from the working concentration of 10.26 to 10.18 ± 4.00, 10.30 ± 1.00, 10.18 ± 3.00, and 6.41 ± 2.65 log10 PFU/mL at of 0.5, 5.0, 10.0, and 15.0 NaCl concentrations, respectively. ECPA_4 show a slight increase from the working concentration of 8.08 to 10.11 ± 1.00, 10.20 ± 1.00, and 9.11 ± 3.00 log10 PFU/mL at NaCl concentrations of 0.5, 5.0, and 10.0, respectively, but at an NaCl concentration of 15.00, there is no apparent plaque.
Chloroform stability test
Figure 7 illustrates the phages' stability at a concentration of 10% v/v chloroform. The phages show a decrease from a working concentration of 12.80 to 9.76 ± 2.51, 9.79 ± 1.00, 9.75 ± 6.56, and 9.78 ± 4.36 log10 PFU/mL for the phages ECPA_1, ECPA_2, ECPA_3, and ECPA_4, respectively.
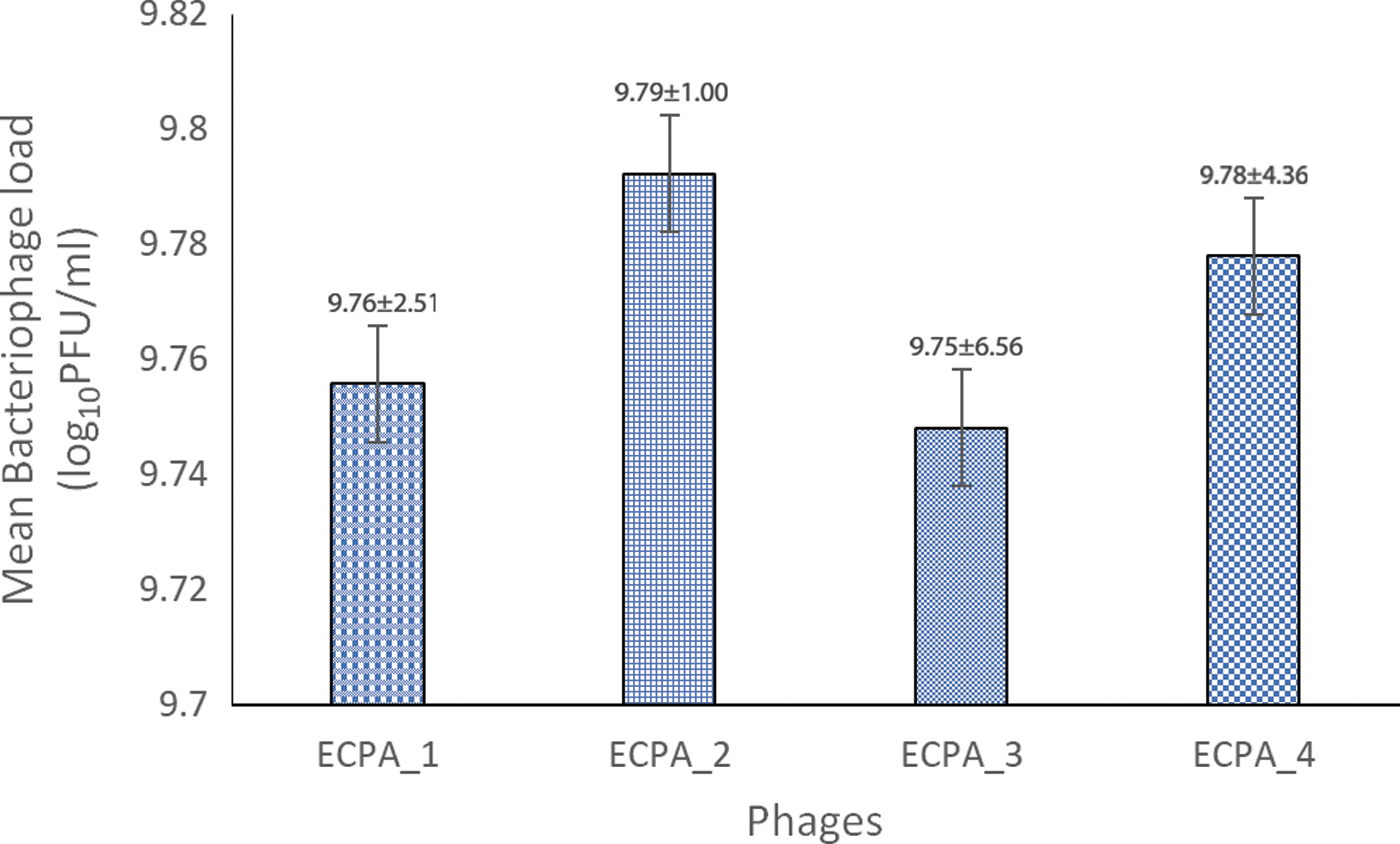
Chloroform stability test. Working concentration = 12.80 log10 PFU/mL.
Host range of the test organism and some selected pathogenic organisms
Table 5 depicts that except for Salmonella Typhi_2, Staphylococcus capitis, Salmonella pullorum_2, Escherichia coli_026, and Pseudomonas aeruginosa_2, ECPA_1 has activity against selected pathogenic organisms, with a moderate activity against Pseudomonas aeruginosa_1. ECPA_2 has strong activity against Salmonella pullorum_1, Salmonella Typhi_1, Salmonella Typhi_3, Klebsiella pneumonia, S. Aureus, and Escherichia coli O157:H7; no activity against Salmonella Typhi_2, Salmonella pullorum_2, Salmonella pullorum_1, S. capitis, Pseudomonas aeruginosa_2, and Escherichia coli 026; and a little activity against Pseudomonas aeruginosa_1.
Host Range of the Test Organism and Some Selected Pathogenic Organisms
Key: −, no reaction; +, turbid; ++, slight turbid; +++, clear.
K. pneumonia, Salmonella pullorum_1, Salmonella Typhi_3, S. aureus, and Escherichia coli O157:H7 had moderate activity, whereas Salmonella Typhi_1, Salmonella Typhi_2, Salmonella pullorum_2, Pseudomonas aeruginosa_1, Pseudomonas aeruginosa_2, and Escherichia coli_026 recorded no activity. ECPA_4 has a high level of activity against Salmonella Typhi_1, Salmonella Typhi_3, Salmonella pullorum_1, K. pneumonia, S. aureus, and Escherichia coli O157:H7; no activity against Salmonella Typhi_2, Salmonella pullorum_2, S. capitis, Pseudomonas aeruginosa_2, and Escherichia coli 026; and slight activity against Pseudomonas aeruginosa_1 (Figs.8 and 9).
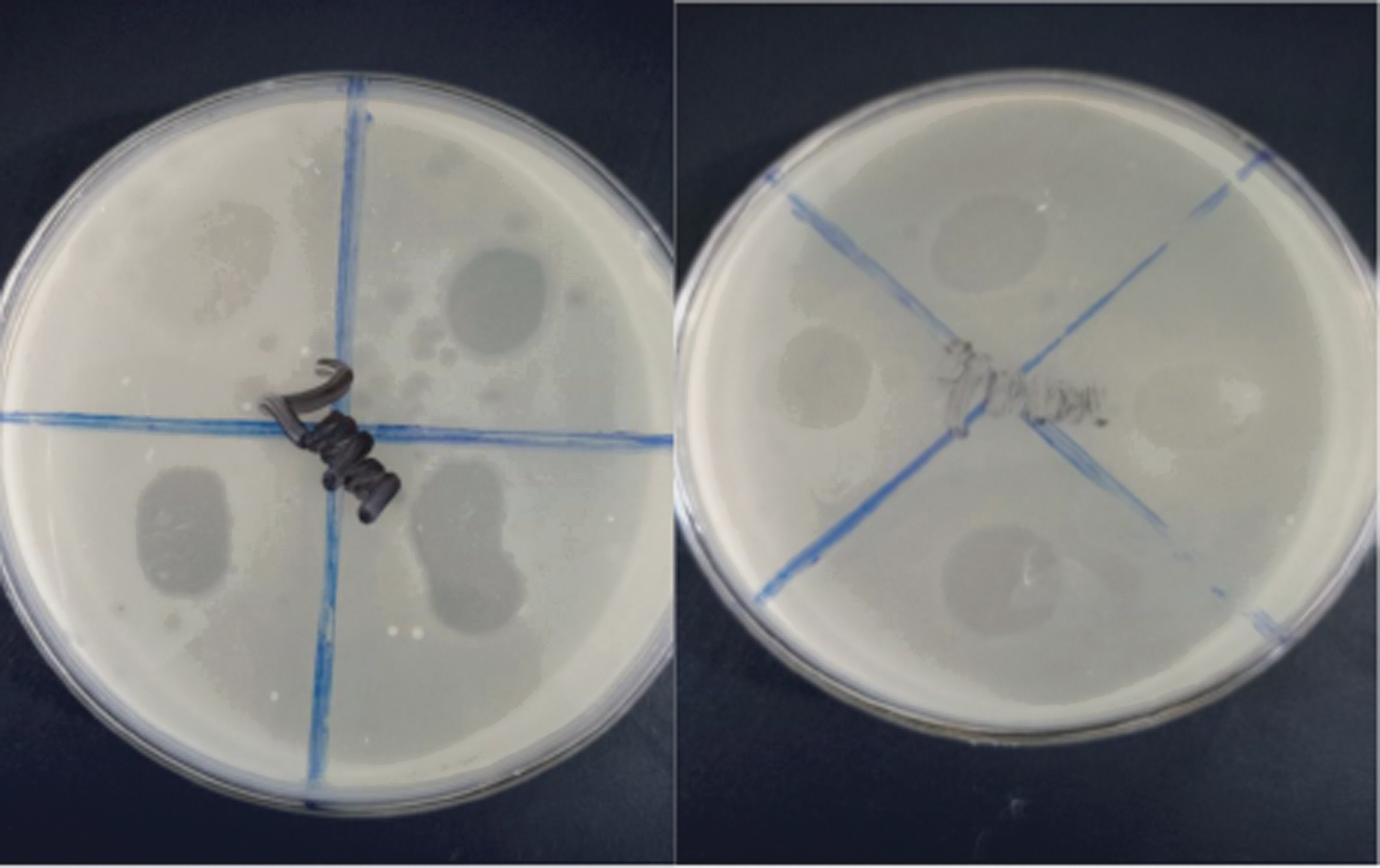
Spot assay showing clearing/lysis zones. The clearing zones represent the virulent activity of bacteriophages, which are susceptible to the host bacterium (Escherichia coli O157:H7).
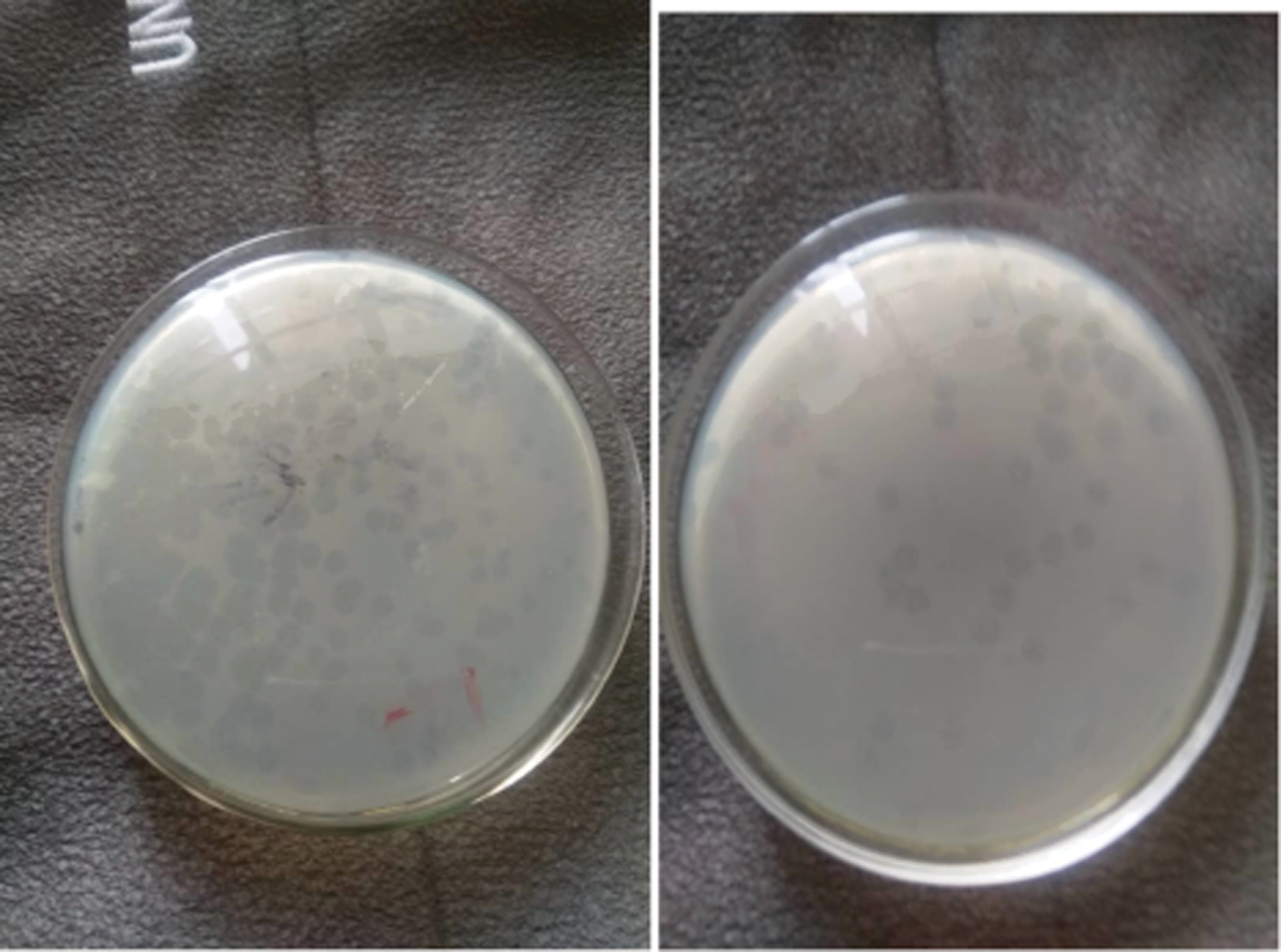
Plaque assay showing the presence of individual plaques. The plaques are considered as bacteriophages and are counted to determine the bacteriophage titer (PFU/mL).
Discussion
According to the findings of this investigation, bacteriophages isolated from effluent water and soil have lytic activity against Escherichia coli O157:H7 and a vast community of bacteria. The incidence of Escherichia coli O157:H7, which has mostly emerged in recent years as the causative agent of hemorrhagic colitis in humans, was a motivation for this investigation. 35 This illness, with characteristic symptoms of bloody diarrhea and abdominal cramps, can progress into a more severe, life-threatening complication known as a hemolytic uremic syndrome. 37
In the current scenario, the serious social and medical problem has resulted from the increasing antibiotic resistance of bacterial strains.38,39 Pharmaceutical companies are retreating from the research and development of new antibiotics due to the unprofitability of the endeavor and the possibility of bacterial resistance developing, 40 which has sparked interest in alternate microbial control systems to antibiotic therapy. 41 Antibiotics may be replaced by lytic phages to treat bacterial illnesses that do not respond to traditional antibiotic therapy.42,43
Total viable count for environmental isolates
Angwa Rukuba_1, Old JUTH_1, Student Village Hostel_2, and Old JUTH_2 had the highest bacterial count, whereas Angwa Rukuba_2 and Student Village Hostel_1 had the lowest bacterial load count. This confirms that pathogenic bacteria can be isolated from the environment, which supports the findings of Obayiuwana et al. 44 carried out in wastewater obtained from pharmaceutical facilities; a significant colony of bacteria was discovered in Nigeria's Lagos and Ogun States.
This finding is consistent with the findings of Mulamattathil et al. 45 who conducted research in Mafikeng, North West Province, South Africa, and found that some of these species are indicators of fecal contamination, as evidenced by the presence of heterotrophic bacteria, total coliforms, fecal coliforms, Aeromonas, and Pseudomonas in water samples.
Identification of environmental isolates
A total of 50 bacterium isolates were found from the cultural identity of environmental isolates, with Bacillus spp. having the highest percentage of occurrence, followed by Escherichia coli and P. aeruginosa. Proteus spp. and S. aureus had the least number of occurrences, followed by Klebsiella spp. This agrees with the research findings of Egbere et al. 46 in the area of the isolates but disagrees with the percentage occurrence of the isolates where the most common occurrence is Proteus spp., followed by P. aeruginosa, E. coli, Salmonella spp., S. aureus, and Streptococcus faecalis from hand-washed samples of internally displaced persons in Jos and surroundings had the lowest percentage occurrence.
It also supports the findings of Bolaji et al. 47 on the antibiotic resistance pattern of bacteria spp. isolated from hospital wastewater in Ede South Western, Nigeria, where the following organisms were isolated from wastewater: Enterobacter aerogenes, Pseudomonas putida, Pseudomonas fluorescens, Klebsiella edwardsii, Proteus mirabilis, and P. aeruginosa.
Antibiotics susceptibility profile of the test organism and environmental isolates
The environmental isolates show a low level of antibiotic resistance to Gentamicin, Ciprofloxacin, Nitrofurantoin, and Ofloxacin. Except for a strain of P. aeruginosa, all isolates showed significant levels of antibiotic resistance to Cefuroxime, Ampicillin, and Augmentin, and absolute resistance to Ceftazidime.
This backs the research findings of Adewale et al. 48 on the antibiotic sensitivity of bacteria associated with chosen waste dumpsites in Akure, Nigeria, where Bacillus subtilis, P. aeruginosa, S. aureus, Salmonella Typhi, S. faecalis, S. dysenteriae, E. coli, Serratia marcescens, and Clostridium botulinum are susceptible to antibiotics except Micrococcus luteus, which was resistant to all the antibiotics used, thereby posing a potential threat to the local inhabitants of the area. 48
Egbere et al. 46 disagree with these findings in their study of the hand hygiene status of internally displaced persons in Jos and surroundings, Nigeria., where S. faecalis, S. aureus, E. coli, Salmonella spp., P. aeruginosa, and Proteus spp. were the most resistant organisms to the antibiotics followed by P. aeruginosa. 46 The highly sensitive organisms were S. aureus, supporting these research findings, and S. faecalis respectively.
Phage distribution to sample location
A total of four phages, ECPA_1, ECPA_2, ECPA_3, and ECPA_4, were isolated from six different locations of effluent water and soil in Jos metropolis, confirming the findings of Ezemokwe et al. 31 on the complete genome sequence of pseudomonas phage Zikora, a temperate bacteriophage isolated from sewage water in a hospital environment in Jos, Plateau State, Nigeria. This backs up the findings of Sundar et al., 49 who recovered three bacteriophages from sewage water obtained using sterile dark containers from the Jinke Park Sewage Treatment Plant in Bangalore, India. 49
Stability test for the phages
Due to nucleic acid and protein denaturation, high temperatures have been proven to inactivate phages.50,51 Myoviridae phages decreased phage activity (3.5 logs PFU/mL) after 60 min at 60°C, according to Yamaki et al. 51 However, the four phages in the current study exhibit stability at temperatures ranging from 45 to 60°C, with no apparent plaque at temperatures over 60°C.
This is in line with research by Litt and Jaroni 32 on the isolation and physio-morphological characterization of Escherichia coli O157:H7-infecting bacteriophages recovered from beef cattle operations, where the phages were stable for 24 h at a temperature range of 40–60°C and for 90 days in cold storage.
In comparison to phages in earlier investigations, phages in this study were stable over a wide pH range (2–9).52,53 The titer of the Escherichia coli O157:H7 phage, AKFV33, fell by 1.9 logs PFU/mL after 15 min at pH 3 and was undetectable after 2 h, according to Niu et al. 52 Only phage ECPA_4 and phage ECPA_1 survived at pH 2 and pH 9, respectively, in the current investigation.
Except for phage ECPA_4, which did not survive at pH 7 after 24 h, all of the phages survived at pH 5 and 7. At low pH, phages can acquire nonreversible mutations, which could explain phage survival in our study. Strack et al. 54 found a linear association between phage mutation rate and low pH incubation, implying that phages can acquire mutations to survive in an acidic environment. Strack et al. 54 discovered a linear link between phage mutation rate and low pH incubation, implying that phages can acquire mutations to survive in an acidic environment.
The phages were also stable at alkaline pH (7–9) in the current investigation, with no titer loss at pH 7 and a maximum titer loss at pH 9. In the current study, the four phages were stable at NaCl concentrations of 0.5–15%, except phage ECPA_4, which shows no visible plaque at 15. This investigation supports the findings of Duyvejonck et al., 55 who found that all phages survived at an NaCl concentration of 0.9 without losing titer.
High stability in low salt concentration (0.5–5%) was shown by the isolated phages. But increasing the concentration (10–15%) led to a greater reduction in phage activity. High quantities of salt, up to 5 M in some situations, did not affect the phage titer, according to investigations conducted by some researchers. 56 The osmotic pressure exerted on phage capsids prevents the ejection of phage DNA, 57 or the aggregating impact of high salt concentration on phage particles with a concurrent reduction in bacterial binding sites.57,35
During the isolation, purification, and preparation of phages, some chemicals such as chloroform are added to the growth medium to accelerate lysis and also included in the phage lysates to prevent bacterial contamination. 58 After 24 h, the separated phages demonstrated a high level of stability against chloroform treatment. For these phages, it is recommended that 10% chloroform be added to the phage medium during isolation and storage.
Chloroform was not employed in this study since the sensitivity to this drug was unclear. Some writers have reported varying results on phage susceptibility to chloroform, implying that each phage's sensitivity to chloroform is validated before chloroform treatment.
The test organism's host range and a few selected pathogenic organisms
Bacteriophages, like all viruses, are extremely species-specific in terms of their hosts and often only infect one or a few strains of one particular bacterial species. 59 In this study, all the isolated phages showed high lytic activity against the test organism Escherichia coli O157:H7 and some of the selected pathogenic bacteria, suggesting that they have a broad host range.
Studies have revealed that phages unique to one Escherichia coli O157:H7 strain can also infect other O157:H7 strains.60–62 Raya et al. 60 found that phage AR1 lysed all O157:H7 strains tested, whereas Goodridge et al. 63 found that T-even bacteriophage (CEV1) infected 17 of the 19 Escherichia coli O157:H7 strains examined. The Results section from the current study is similar to these studies, revealing that isolated phages with high target specificity are virulent against a wide range of Escherichia coli O157:H7 isolates.
The isolated phages could, thus, be used to prevent Escherichia coli O157:H7 transmission and survival in the food chain. However, the growth of phage-resistant bacteria, particularly when phages are used in animals or on farms, is a serious worry with this strategy. By keeping selection pressure on the bacterial host, the use of phage cocktails that are continuously updated with new or different phages could potentially address the problem.64,65
Conclusion
Overall, bacteriophages infecting Escherichia coli O157:H7 were isolated and characterized in this study. They showed a wide range of host specificity along with high lytic activity and optimal physicochemical stability, and, therefore, could be considered promising candidates for the biocontrol of an emerging food-borne pathogen Escherichia coli O157:H7 in the area of the food industry. Future research should be directed to the whole genome of the bacteriophages to ascertain their safety and develop effective phage cocktails.
Footnotes
Acknowledgments
A preprint of this manuscript has previously been published on Research Square (DOI: 10.21203/rs.3.rs-1399481/v1). 66 The authors would like to thank the Westerdijk Fungi Biodiversity Institute (Medical Mycology), Netherlands, for MALDI-TOF MS analysis.
Author's Contributions
A.C.N.: Conceptualization, methodology, data curation, and writing-original draft. J.O.E. and I.A.O.: Supervision, review, and editing. U.U.: Methodology, visualization. G.C.E. and N.G.: Data curation. N.E.N.: Methodology, project administration. J.C.A.: Resources, supervision.
Consent for Publication
The authors hereby declare their interest to submit the above manuscript for your consideration for possible publication. By this, they affirm that this paper is only being considered for publication in this journal alone and has not been submitted to any other journal. They also affirm that the content of this journal is original and has followed the journal template. They, therefore, state that the enlisted co-authors in this manuscript made significant scientific contributions. However, the author whose name is missing in R1 and R2 that initially appeared in the original version of the manuscript voluntarily requested for the removal of his name in the manuscript.
Availability of Data and Material
Data sharing is not applicable to this article, as no datasets were generated or analyzed during the current study.
Author Disclosure Statement
The authors declare that they have no conflict of interest. The authors declare no competing interest.
Funding Information
This work was funded by a grant from the Africa Centre of Excellence in Phytomedicine Research and Development (ACEPRD) at the University of Jos to A.C.N. by the World Bank Group (WBG) grant number ACE033 for the esterblishment of ACEPRD.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.