
Abstract
Introduction
The mechanisms of action of infrared and visible low-level laser on cells differ [4], but their clinical results are quite similar. The LPLI-triggered photobiomodulated cell response depends on a combination of parameters, such as wavelength, energy density, and power density. 2,4,5,6 These factors may contribute to an increase in collagen synthesis, cell proliferation, and mitochondrial respiration.
In vitro and in vivo studies have demonstrated that laser irradiation exerts significant influence over cell function (cytoskeleton organization, mitochondrial activity, and plasma membrane stimulation). 7 The laser photobiomodulatory effect on mitochondria triggers the activation of ATP production through an increase in the mitotic process due to excitation of cell respiration and endogenous porphyrins. However, the increase in mitochondrial ATP produced by the variation in the energy absorbed by the cell also affects cell metabolism (for example, mitochondrial oxide reduction), resulting in a cascade of biochemical reactions. 8,9 The irradiation of isolated mitochondria also affects the transcription and transduction of mitochondrial proteins, thereby increasing the reaction cascade, as well as the number of components in the respiratory chain (cytochrome oxidase and flavin dehydrogenase). 10
The MTT assay [3-(4.5-dimethylthiazol-2-yl)-2.5 diphenyltetrazolium bromide] is one of the most frequently used assay methods for measuring cell proliferation and cytotoxicity. In living cells, tetrazolium salt MTT is reduced to formazan [1-(4.5-dimethylthiazol-2-yl)-3,5 diphenylformazan], which cannot pass through the cell membrane and therefore accumulates in healthy cells and can be colorimetrically analyzed. A reduction in MTT is generally attributed to mitochondrial activity, but it has also been related to non-mitochondrial enzymes as well as endosomes and lysosomes. If the MTT assay were to reflect mitochondrial metabolic activity precisely, it could be useful for the evaluation of cell viability. This assay has been tested for validity in various cell lines. 11,12,13,14,15
The aim of the present study was to determine the action of different levels of laser photobiomodulation on L929 fibroblast cells at 904 nm and energy densities of 6 J/cm2 and 50 mJ/cm2.
Materials and Methods
L929 cells (mouse conjunctive tissue - ATCC CCL-1 NCTC) (Cell Culture Laboratory, Instituto Adolfo Lutz, Sao Paolo, Brazil) were used in in vitro toxicity tests, as recommended by ISO 10993-5 (Biological evaluation of medical devices-Part 5-Tests for in vitro cytotoxicity). The cells were routinely cultivated in 25 cm2 flasks (TPP, Trasadingen, Switzerland) in Minimal Essential Medium (MEM) (Gibco™-Invitrogen Corporation, Grand Island, New York) supplemented with 10% (vol/vol) bovine fetal serum (BFS) (Cultilab Materiais para Cultura de Células Ltda, Campinas, Brazil), 100 U/mL penicillin, 100 mM/mL streptomycin, and 0.25 μg/mL fungizone™ (GibcoBRL, Gibco-Invitrogen, Grand Island, NY). The cells were cultured in an incubator (Thermo Forma Scientific, Waltham, MA) at 37°C in a humidified 5% CO2 atmosphere. The medium was changed every two days. When the cells became confluent, the medium was removed and the cell layer was washed with phosphate-buffered saline. 0.25% Trypsin™ (Gibco BRL) in buffered ethylenediaminetetraacetic acid (EDTA) (Carlos Erba, ABC Lab, São Paulo, Brazil) was then added and incubation continued for 2–4 min. A solution with an approximate concentration of 1 × 104 cells/mL was prepared and poured into each well of 96-well culture plates (TPP™, Trasadingen, Switzerland). The protocol was approved by the Research Ethics Committee of Univap (process No. A061/CEP/2006).
Irradiation
A gallium arsenide (GaAs) diode laser (KLD™ Biosistemas Equipamentos Eletrônicos Ltda, São Paulo, Brazil) emitting radiation at λ = 904 nm; with a repetition rate of 10 KHz; output power of 50 mW; pulse width of 100 ns; peak power of 50 W; beam area of 0.01 cm2; divergence of 8° × 25°, and 0.1% active cycle; was used to irradiate three groups of fibroblast cells sub-cultured in 96-well culture plates (TPP™, Trasadingen, Switzerland) at a density of 1 × 104 cells/mL. The distance of the beam from the cell cultures was 1.5 cm. Three groups were established: Group 1, non-irradiated (control); Group 2, irradiated at 6 J/cm2; and Group 3, irradiated at 50 mJ/cm2. The cell cultures were cultivated in a well with 0.3 cm2 of cross section receiving radiation perpendicularly to the plate at 24-h, 48-h, and 72-h intervals for 36 sec at 6 J/cm2 and 2 sec at 50 mJ/cm2. Control cells (non-irradiated) were submitted to the same condition as the laser-irradiated cells. Prior to irradiation, the laser was calibrated according to manufacturer's instructions, using an Agilent oscilloscope (Agilent Technologies, Inc., Colorado Springs, CO).
Cell viability by MTT assay
For the MTT assay, 1 × 104 cells/mL L929 cells were plated in each well of a 96-well plate. After each incubation period with 24-h intervals, the cytotoxic effect of laser irradiation on cell activity was analyzed by MTT assay. This assay measures the percentage of cell survival in comparison with untreated controls. It is widely used for analyzing cytotoxicity, cell viability, and proliferation. 15 Each well received 20 μL of MTT to a final concentration of 0.5 mg/mL and the plate was incubated for 1 h at 37°C in a 5% CO2 atmosphere. An aliquot of 100 μL of dimethyl sulfoxide (DMSO) was then added to each well and the plate was agitated for 30 min for solubilization of the formazan crystals. Crystal concentration was spectroscopically quantified using a microplate reader (ELISA Reader - Spectra Count, Packard Instrument, USA) at an excitation wavelength of 570 nm.
Fluorescence Microscopy Procedures
All dyes used in the fluorescence studies were purchased from Molecular Probes (Eugene, OR). Just prior to imaging, the cells were stained by incubation in a fluorescent dye-containing growth medium at 37°C as follows:
MitoTracker Orange was used as a probe for evaluation of mitochondria membrane potential changes (Δψm). After each incubation interval (24, 48, and 72 h), the cells were stained with MitoTracker Orange Dye (CMTMros™, Molecular Probes, Eugene, OR) [10] (150 nM), incubated for 20 min in the dark, immediately washed with PHEM buffer (60 mM PIPES, 20 mM HEPES, 10 mM EGTA, 5 mM MgCl2) and fixed in 3% PA (paraformaldehyde) (Sigma-Aldrich Chemie GmBh, Steinheim, Germany) in 0.1 M PHEM for 10 min. DioC6 (3,3′- dihexyloxacarbocyanine iodide) was used to study endoplasmic reticulum changes. The cover slips were rinsed with PBS, incubated with 10 μg/mL DioC6 for 30 min, immediately rinsed twice with PBS, fixed with 4% PA in PHEM buffer (0.I M) for 10 min and rinsed again to eliminate excess dye. Cover slips were mounted on slides with n-propyl gallate and observed through fluorescence microscopy. Observations and photographs were made using a Leica microscope (DLMB, Leica Microsystems, Bannockburn). For fluorescence analysis, approximately 100 cells were observed on each plate. Plates were observed and photographed in triplicate for the increase in fluorescence intensity of organelles. Excitation (554 nm) and emission (576 nm) filters were used.
Statistical Analysis
Descriptive statistics were used to describe the results expressed as mean ± Standard Error of the Mean (± SEM). Comparisons between groups were performed with two-way analysis of variance for repeated measures (ANOVA). Post hoc test Tukey's HSD (Tukey-Kramer method) analysis was used to determine significant differences between groups. Values of p ≤ 0.05 were considered statistically significant (95% confidence interval). Statistical analysis was performed with the GraphPad Prism (version 4.0) package.
Results
The effects induced on L929 cells by laser radiation at energy densities of 6 J/cm2 and 50 mJ/cm2 were determined with the aid of fluorescence staining and MTT assays in Groups 2 and 3 (irradiated) following each 24-h incubation interval. Group 1 (non-irradiated) was submitted to the same analyses.
Fluorescence microscopy demonstrated that the mitochondria of MitoTracker-stained control cells were dispersed in the periphery of the cytoplasm, with evident nucleus localization in the absence of fluorescence (Fig. 1a). The same occurred with DioC6-stained control cells, with the endoplasmic reticulum spread throughout the entire cytoplasm (Fig. 1b). However, MitoTracker was distributed throughout the cell in the groups irradiated with 6 J/cm2 and 50 mJ/cm2. Stain concentration in the perinuclear region suggests intense mitochondrial activity (Fig. 1c-d), as confirmed by the intense reticular activity observed by DioC6-staining stemming from increased protein synthesis (Fig. 1e-f). The fluorescence intensity of all irradiated cells was greater than that of the non-irradiated cells, indicating increased ATP synthesis.
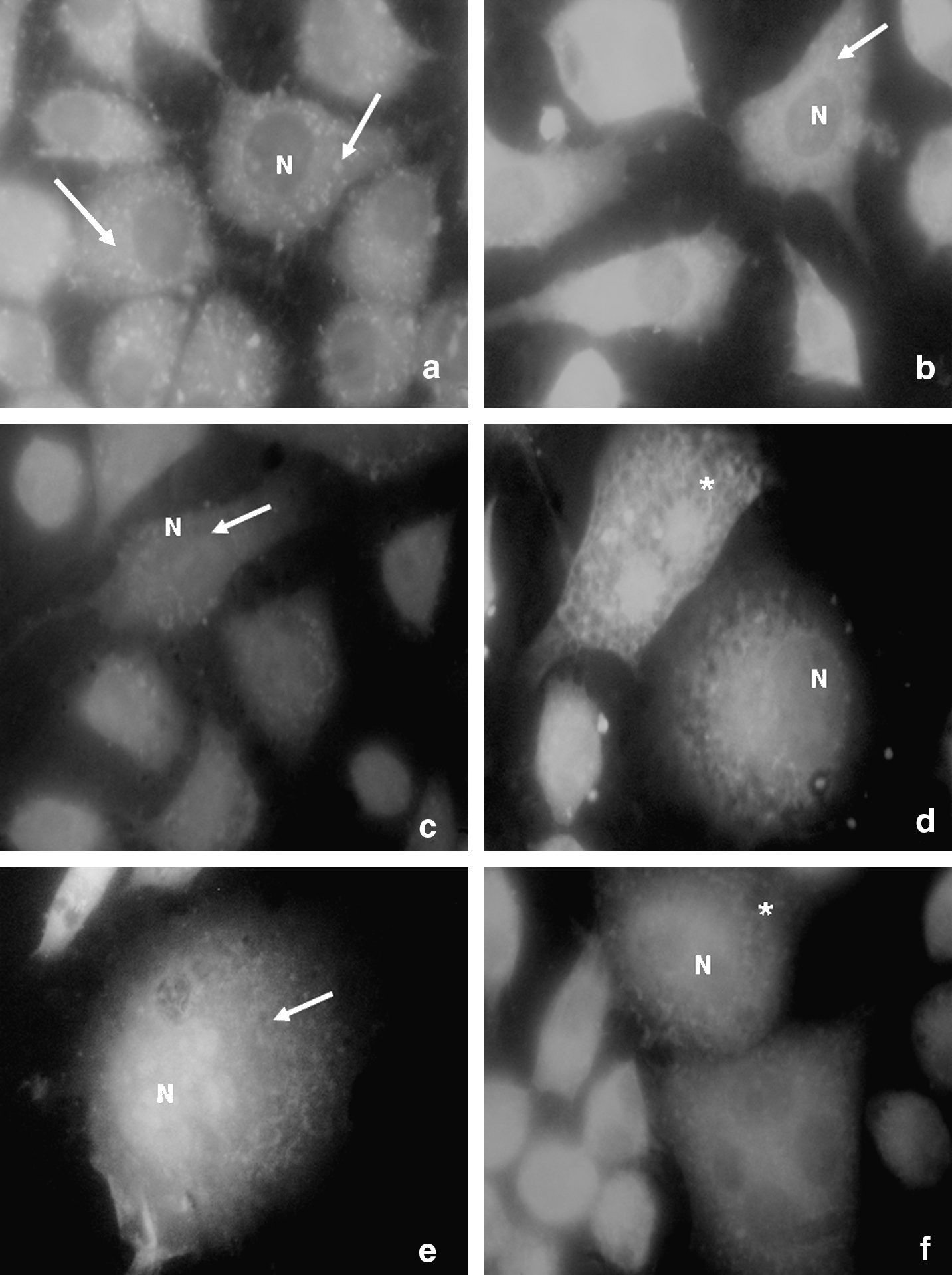
MitoTracker Orange™ and DioC6-stained L929 cell death after 72 h. (
These results corroborate those of other authors, who report that mitochondria are sensitive to monochromatic visible and near-infrared (IR) light irradiation. 16,17 Mitochondria are believed to be the primary targets when whole cells are irradiated with visible and near-infrared light and an increase in ATP synthesis is observed. As the mitochondrial membrane potential (Δψm) is directly related to ATP production and synthesis stimulation, cell metabolism may be significantly affected.
Simultaneous comparison by MTT assays revealed that Groups 2 and 3 exhibited an increase in cell viability at the two different energy densities with the increase in post-irradiation time in comparison to the control (p ≤ 0.05) (Table 1). Concerning temporal evolution (24, 48, and 72 h), cell growth occurred with similar intensity in both irradiated groups, regardless of the energy density used when comparing 24-h and 72-h post-irradiation within a single group (p = 0.01) (Table 2).
Mean.
Standard error mean.
p ≤ 0.05.
Mean.
Standard error mean.
p ≤ 0.05.
-Nd, not defined: values which have variance and standard deviation equal to zero.
Discussion
In the present study, L929 cells submitted to proliferation and MitoTracker-staining exhibited mitochondria distributed throughout the cell, including the perinuclear region, thereby suggesting intense mitochondrial activity. Mitochondria in the perinuclear region may be associated with the endoplasmic reticulum, with bi-directional calcium flow as well as acidic vesicles, which may play an important role in proton gradient regulation across the mitochondrial membrane. Fluorescence analysis by DioC6 staining revealed intense reticular activity resulting from post-irradiation protein synthesis. 18
When the concentration of MTT in the cytoplasm is increased, more formazan products are formed and higher irradiation absorption by the cells is detected, indicating greater cell viability. MTT is not permeable to the lipid plasma membrane and it has been suggested that MTT enters the cells through endocytosis. A reduction in MTT has been associated with mitochondrial activity as well as endosome and lysosome enzymes. 13 MTT can also be reduced by the receiving electrons in mitochondria and other cellular organelles, indicating that it is dependent on cellular redox activity. Therefore, a reduction in MTT reflects mitochondrial function and may be used as an indicator of cellular oxidative metabolic activity. 12
Photobiomodulation of L929 fibroblast cells with near-infrared radiation from GaAs laser (904 nm) produces biological effects that are directly dependent on dosimetry. 2,4,7,18 The pulsed GaAs laser was used to administer different energy densities to human embryo fibroblast cell cultures and a significant increase in collagen levels was observed in irradiated cells. 19
Both energy densities used in the present study (6 J/cm2 and 50 mJ/cm2) resulted in cell growth dependent on post-irradiation time. An inhibitory effect has been observed for high cumulative doses with energy densities above 10 J/cm2 over long periods. 3,20 The cumulative doses used in previous studies using 0.15 J/cm2 and 18 J/cm2 were not found to be inhibitory, even at 18 J/cm. 2,3,20 Furthermore, different cell proliferation responses have been recorded at different wavelengths (665 and 810 nm), along with inhibition following fibroblast cell irradiation in the infrared band (810 nm). 3 Another aspect to be considered is the different wavelengths used, as the present study used 904 nm.
Cell irradiation with visible light has both positive effects (acceleration of cell division) and negative effects (damage to the intracellular system and apoptosis). Moreover, the occurrence of cell inhibition is related to the cumulative effect of the dosage accompanied by growth inhibition, cell death, and the destruction of photoreceptors. There is accelerated electron transfer in the redox pairs in some sections of the respiratory chain as well as transfer of energy from oxygen excitation, which explains the photobiomodulatory effect of low-intensity radiation. 4
A cascade of events has been proposed, beginning with the absorption of light by a photoreceptor that conducts photo-activated enzymes in the interior of mitochondria and the resulting photo-response. Light absorption by components of the respiratory chain leads to changes in the mitochondria and cytoplasm. The magnitude of the laser photobiomodulatory effect depends on the physiological state of the cell at the moment of irradiation. This may explain why photobiomodulation is not always detectable. It may also explain the variability of results reported in the literature. 4
Laser therapy is commonly associated with cell stimulation by both visible and near-infrared radiation through the membrane potential or mitochondria. Comparing HeLa and TK6 cells irradiated with continuous 808 nm and pulsed 905 nm laser doses of 1-60 J/cm2, better results and dose dependence have been found at 905 nm. 9 In the present study, irradiation at the 904 nm wavelength increased cell proliferation at both 50 mJ/cm2 and 6 J/cm2 when compared with the control group. The same type of pulsed laser on fibroblast cell cultures has also resulted in better cell proliferation at an energy density of 5 J/cm2. 2 Our results corroborate those of another study that compared the effect of low-level ultrasound and low-level laser with the same laser parameters as the present study and demonstrated an effect on cell growth at 6 J/cm2 and 50 mJ/cm2, with a higher effect at 50 mJ/cm2. 21
Irradiation of L929 cells with pulsed laser in the near infrared region (904 nm) at two quite distinct energy densities allowed the observation of increased cell proliferation in both cases. The result was slightly higher for an energy density of 50 mJ/cm2 than for 6 J/cm2, as demonstrated by fluorescence microscopy. Cell response was also found to be better 72 h after irradiation.
Footnotes
Acknowledgments
We thank CAPES, FAPESP for grant No. 01/07380-6, KLD™ Biosistemas Equipamentos Eletrônicos Ltda and Nelson Fuirini Junior, Brazil for supporting this study.
Author Disclosure Statement
No competing financial interests exist.