
Abstract
Introduction
Several investigators have suggested that the use of low-level laser therapy (LLLT) can promote faster epithelization and greater vascularization and collagen synthesis of surgical wounds. 7 –17 In addition, studies regarding the effects of laser therapy on cell cultures obtained satisfactory results of cellular proliferation and modulation of growth factors. 18 –24
However, there is a lack of scientific evidence demonstrating the influence of LLLT on physiologic and pathologic processes of angiogenesis, including the expression of VEGF in animal models. Therefore, the aim of the present study was to investigate the kinetic expression of VEGF-A165 messenger ribonucleic acid (mRNA) during wound healing of tongues of Wistar rats after laser therapy using an infrared laser (gallium-aluminum-arsenide (GaAlAs), 780 nm) associated or not with a visible red laser (aluminum-gallium-indium-phosphide (AlGaInP), 660 nm).
Materials and Methods
Animals and characterization of experimental groups
The Ethics Committee for Animal Research of the Bauru School of Dentistry, University of São Paulo approved the present study (process # 02/2005). Forty-five healthy rats (Rattus norvegicus, albinus, Wistar) weighing between 240 and 340 g were used for the study. All rats were fed a standard rat diet and given untreated water ad libitum.
The animals were randomly divided into three experimental groups: I, 5 rats used as normal control animals and not subjected to any surgical procedures; II, 20 rats that were operated on; and III, 20 rats that were operated on and subjected to laser irradiation. The weight of all animals was used to control their patterns of growth and development.
Surgical procedure
After anesthesia with pentobarbital sodium (60 mg/kg, intraperitoneal injection), full-thickness, 12.4-mm2 excision wounds were made using a 15C scalpel blade on the right side of the midline of the tongue in the 40 rats of groups II and III (Fig. 1).

Illustration representing the surgical wound performed on the tongue of rats. Measurements are in millimeters.
Laser therapy
Two types of nonpulsed, low-intensity lasers were used in the therapeutic procedures: an infrared diode laser (GaAlAs; λ780 nm; power, 70 mW; spot size, 0.04cm2) and a visible red diode laser (AlGaInP; λ660 nm; power, 40 mW; spot size, 0.04cm2). the manufacturer had previously calibrated both lasers (MMOptics, São Carlos, Brazil).
Two sessions of laser irradiation were accomplished. In the first, infrared laser was used immediately after the surgical procedure (energy density, 35 J/cm2; total time, 40 s; total energy, 2.8 J). In the second, 48 h later, the animals were anesthetized similarly to the first session, and the visible red laser was used (energy density, 5 J/cm2; total time, 10 s; total energy, 0.4 J). The laser doses were chosen in accordance with recommendations of the manufacturer. Both lasers, positioned 1 mm away from the surgical wound, were used perpendicularly at two distinct points: on the center of the dorsal and ventral surfaces.
Sacrifice of animals and collection of tissues
Five animals each from groups II and III were sacrificed using an ether overdose 1, 3, 5, and 7 days postoperatively. All animals from group I were weighed for 7 consecutive days and sacrificed subsequently. A tongue sample was removed from the animals after sacrifice and placed into microcentrifuge tubes containing TRIzol (Gibco BRL Life Technologies, Frederick, MD), weighed previously on a high-precision balance. The tubes were then stored at −80°C until use for reverse transcription polymerase chain reaction (RT-PCR).
Isolation of RNA
The procedures to isolate total RNA were performed as described by Santos and colleagues. 25 Total RNA was isolated using guanidinium isothiocyanate-phenol-chloroform extraction. The tubes were thawed and weighed again. Briefly, the tissues were homogenized in TRIzol (0.1 g of tissue/1.0 mL of Trizol) using a tissue homogenizer (PowerGen 700, Fischer Scientific, Pittsburgh, PA); then chloroform extraction, isopropanol precipitation, and 70% (v/v) ethanol washing of precipitated RNA were performed. RNA was resuspended in diethyl pyrocarbonate–treated water and stored at −80°C until use. RNA concentration was measured spectrophotometrically at 260 nm, and the quality of RNA was verified by the 260/280 nm ratio.
Detection of VEGF-A165 mRNA using RT-PCR
All samples of total RNA were treated with DNAse, following the manufacturer's instructions (RQ1 RNase – Free DNAse, Promega Corporation, Madison, WI), to avoid possible genomic deoxyribonucleic acid (DNA) contamination. RT-PCR procedures were performed as described by Santos and colleagues. 25,26 First-strand complementary (cDNA) synthesis was performed in a 33-μL reaction volume using 2 μg of total RNA, 0.2 μg of random hexadeoxynucleotides, reverse transcription buffer (45mM tromethamine hydrochloric acid (HCl), pH 8.3; 68mM potassium chloride (KCl); 9mM magnesium chloride (MgCl2)), 0.08 mg/mL of bovine serum albumin, 15mM dithiothreitol (DTT), 1.8mM deoxyribonu-cleotide triphosphate (dNTP), and 150 U of murine leukemia virus reverse transcription, as provided in the first-strand cDNA synthesis kit (Amersham Pharmacia, Piscataway, NJ). cDNA was synthesized during a 60-min incubation at 37°C, and the reaction was terminated by heating at 90°C for 5 min. Reverse transcription products (3 μL) served as the template for PCR amplification, using primers synthesized by Research Genetics (Huntsville, AL): sense 5′-ATC ATG CGG ATC AAA CCT CAC C-3′ and anti-sense 5′- GGT CTG CAT TCA CAT CTG CTA TGC-3′ (VEGF-A165; final PCR product ∼ 80 bp) and sense 5′- GAA GGG TGG GGC CAA AAG-3′ and anti-sense 5′- GGA TGC AGG GAT GAT GTT CT-3′(glyceraldehyde 3-phosphate dehydrogenase (GAPDH); final PCR product ∼318 bp). All PCR reactions were performed in a total volume of 50 μL composed of 20 pmol (0.4μM) of each primer, PCR buffer (20mM tromethamine HCl, pH 8.4; 50mM KCl; 1.5mM MgCl2), 0.2mM dNTP, and 2.5 U of recombinant Taq DNA polymerase (Go Taq DNA Polymerase, Promega). Temperature cycling consisted of an initial denaturation step (2 min at 94°C, VEGF-A165; 2 min at 95°C, GAPDH) followed by 45 cycles of amplification. Each round consisted of denaturation (1 min at 94°C, VEGF-A165; 30 s at 95°C, GAPDH), annealing (30 s at 65°C, VEGF-A165; 30 s at 60°C, GAPDH), and extension (1 min at 72°C, VEGF-A165; 1 min 30 s at 72°C, GAPDH). Samples were incubated for an additional 10 min at 72°C (terminal elongation) after completion of the final cycle. GAPDH was used as a positive control. For each set of primers, RT-PCR was performed on sterile water to check for genomic DNA contamination. No RT was performed for all samples as an additional negative control. A 10-μL aliquot of each sample was electrophoretically size fractionated on a 2.0% agarose gel containing ethidium bromide (0.64 μg/mL). DNA was visualized under ultraviolet light to detect the presence of PCR amplification products at the anticipated sizes. The molecular weight of the PCR products was determined by comparison with the 100-base pair ladder (Amersham Pharmacia). Following the manufacturer's instructions, Scion Image software (Scion Corporation, Frederick, MD) was used to determine the relative expression of VEGF-A165 and GAPDH. The same examiner determined the systematic error according to re-assessment of all images 1 month after the first measurement. The data are presented as the ratio of specific gene VEGF-A165 to the housekeeping gene GAPDH.
Statistical analysis
Most assays were carried out in triplicate. Statistica 5.0 software (StatSoft Inc., Tulsa, OK) was used to assess the data obtained. Differences of expression of VEGF-A165 mRNA between groups I, II, and III were analyzed using two-way analysis of variance (ANOVA) and the Tukey test. The systematic error of the method was evaluated using dependent t-tests. P < 0.05 was considered statistically significant.
Results
The rats were weighed regularly; all grew normally with regard to weight and size. Clinical situations that could delay wound healing, such as infectious or systemic diseases or accidental lacerations of tissue during the surgical procedure, 27 were not observed in the study.
The method used to assess the relative expression of VEGF-A165 mRNA was considered valid after statistical analysis. No statistically significant systematic error was found (p > 0.05), and any errors were considered acceptable and within normal range.
Densitometric analysis of the ethidium bromide–stained bands of VEGF-A165 mRNA was performed, and their intensities were compared with those of the housekeeping gene GAPDH (Fig. 2).
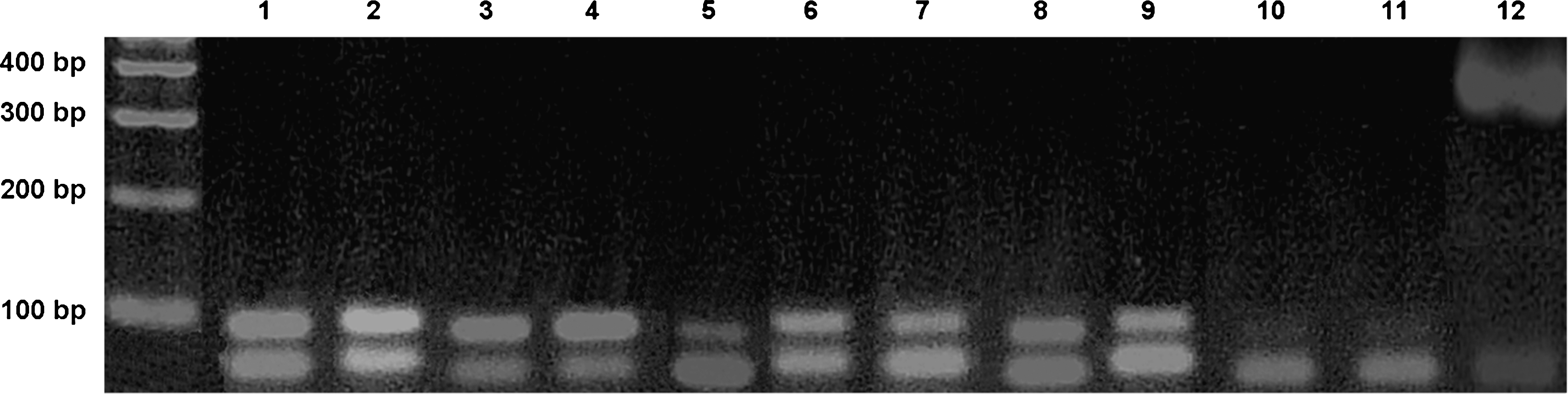
Ethidium bromide–stained agarose gel of reverse transcription polymerase chain reaction (RT-PCR) products to detect and determine the relative expression of vascular endothelial growth factor A165 (VEGF-A165) messenger ribonucleic acid (mRNA) in the tongue of Wistar rats. Lane 1, VEGF-A165 mRNA detected in the tongue of rats operated on and not irradiated (group II), killed on day 1; lane 2, VEGF-A165 mRNA detected in the tongue of rats operated on and irradiated (group III), killed on day 1; lane 3, VEGF-A165 mRNA detected in the tongue of rats operated on and not irradiated (group II), killed on day 3; lane 4, VEGF-A165 mRNA detected in the tongue of rats operated on and irradiated (group III), killed on day 3; lane 5, VEGF-A165 mRNA detected in the tongue of rats operated on and not irradiated (group II), killed on day 5; lane 6, VEGF-A165 mRNA detected in the tongue of rats operated on and irradiated (group III), killed on day 5; lane 7, VEGFA165 mRNA detected in the tongue of rats operated on and not irradiated (group II), killed on day 7; lane 8, VEGF-A165 mRNA detected in the tongue of rats operated on and irradiated (group III), killed on day 7; lane 9, VEGF-A165 mRNA detected in the tongue of rats not operated on (group I); lane 10, water control for VEGF-A165 mRNA; lane 11, PCR of RNA (no RT); lane 12, glyceraldehyde 3-phosphate dehydrogenase (GAPDH) mRNA detected in the tongue of all rats – housekeeping gene.
Two-way ANOVA demonstrated an interaction between the variables “group” (I, II, and III) and “days after surgery” (1, 3, 5, and 7) (F = 8,1505). Therefore, subgroups were formed [I, II (1), II (3), II (5), II (7), III (1), III (3), III (5), and III (7)], and the results of relative expression of VEGF-A165 mRNA took them into account (Table 1, Fig. 3). Significant differences were found between different days after surgery (p < 0.01) and distinct subgroups (p < 0.01), althoughwhen the variable “days after surgery” was not considered, no significant differences were noted between groups I, II, and III (p > 0.05).
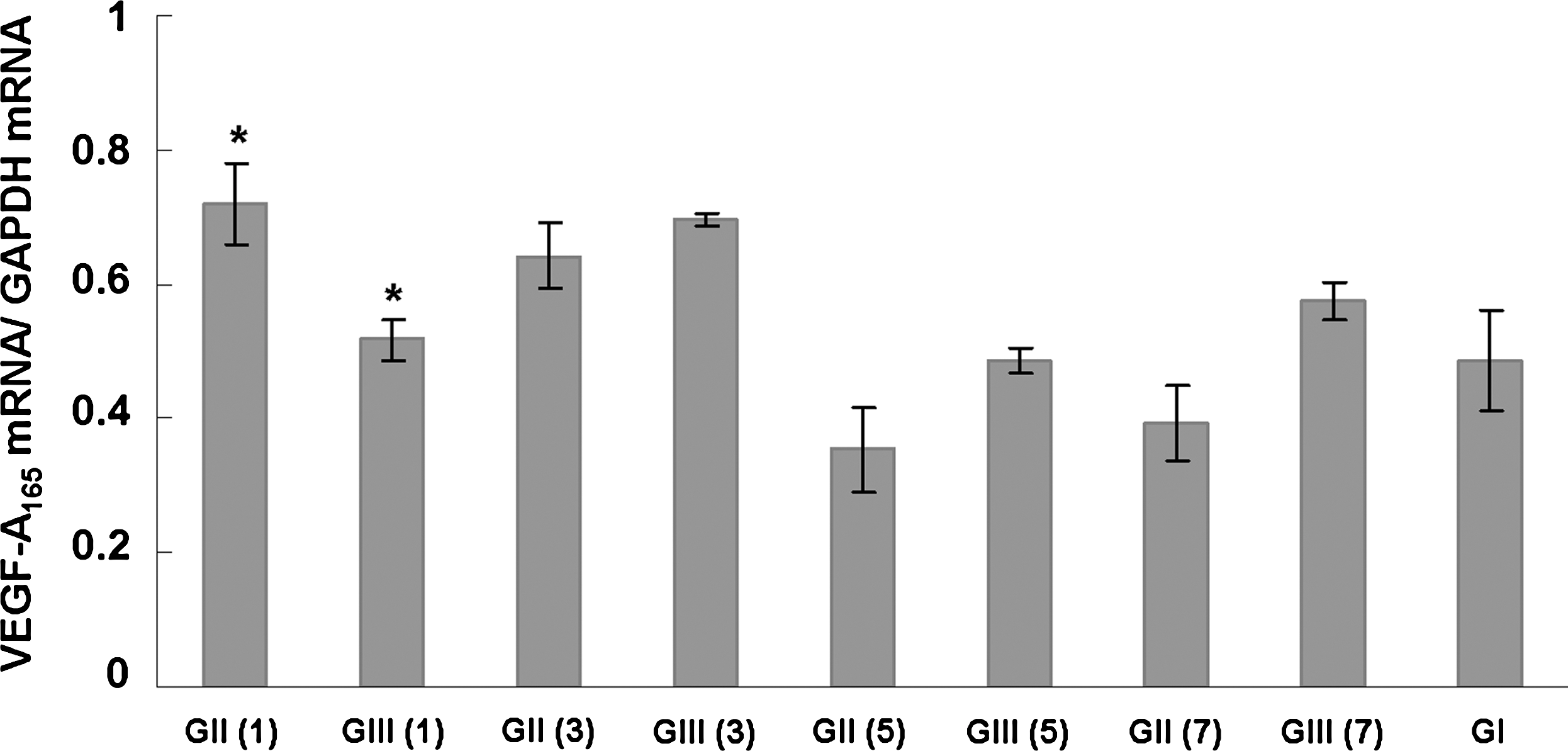
Means of the ratio between the expression of VEGF-A165 mRNA and the expression of GAPDH mRNA in groups I, II, and III and respective days after surgery in parentheses. Error bars show the standard error of each subgroup. *P < 0.05, comparison between the groups I, II, and III on the same day after surgery.
Means followed by the same superscript letters indicate statistically significant differences.
The Tukey test determined significant differences in the relative expression of VEGF-A165 mRNA between the subgroups as follows: I and II (1) (p < 0.05), II (1) and II (5) (p < 0.001), II (1) and II (7) (p < 0.001), II (3) and II (5) (p < 0.01), II (3) and II (7) (p < 0.01), III (3) and III (5) (p < 0.05), and II (1) and III (1) (p < 0.05) (Table 1, Fig. 3).
Discussion
The main finding of the present study is that LLLT influences expression of VEGF-A165 mRNA in vivo. Previous studies demonstrated the capacity of LLLT to produce nucleic acids. 28,29 Angiogenesis is essential to wound healing. 30 VEGF is responsible for the stimulus of proliferation and differentiation of endothelial cells and, therefore, for the sprouting of new blood vessels. 4,31 Most specifically, VEGF-A is considered to be a molecule that is important to inducing angiogenesis and vasculogenesis, 32 and VEGF-A165 is the most abundant isoform. 33 In vivo study of biomodulation of VEGF-A165 mRNA using LLLT can contribute to knowledge about the influence of photonic therapies on tissue repair and wound healing.
The results obtained in this study represent the expression of a growth factor in an animal model, which makes difficult comparison with data obtained in studies that have used other methods. Cells cultivated in vitro are more prone to extrinsic effects of photonic therapies than cells organized in animal tissues, which are naturally protected by physical barriers. 34 Another difference is that most of the in vitro studies analyze the responses of one cellular type to a certain stimulus, which is impossible in studies similar to ours. 21
The type of surgical wound made in this study was chosen based on the descriptions of Davidson, 35 who affirmed that “healing rates are often monitored on the basis of total excisional volume filled with granulation tissue, extent of reepithelization, histological organization of connective tissue, angiogenesis, and biochemical content of collagen or proteoglycans.” The difficulty in developing clinical protocols for the use of LLLT in humans and animals motivated the study of the interaction between two different lasers: an infrared laser and a visible red laser. Previous studies have used only one type of laser on the animals or cells. 7,16,21,22,36
One day after surgery, the use of infrared laser therapy resulted in lower levels of expression of VEGF-A165 mRNA in group III than in group II (infrared and visible red lasers were not used). Furthermore, the levels of expression of VEGF-A165 mRNA observed in groups I and II were similar. The cellular saturation theory is a hypothesis that may explain this finding. Kipshidze et al. 37 demonstrated that high densities of energy were toxic for vascular cells. Normally, high concentrations of VEGF are observed soon after traumatic lesions, such as surgical wounds. VEGF induces the increase of vascular permeability through disorganization of junctional proteins of endothelial cells, 38 and therefore, it contributes to increasing edema and postoperative pain. Infrared laser therapy seems to modulate the expression of VEGF-A165 mRNA to values close to those observed in sound tissues, which may decrease the effects of biological unbalances caused by acute inflammatory process.
In this study, the expression of VEGF-A165 mRNA presented a trend of increase after the third day after surgery, when the surgical wounds in the tongue of rats were irradiated with visible red laser. The findings of Agaiby et al., 20 Kipshidze et al. 22 and Young et al., 39 who demonstrated that low densities of energy can increase growth factor release can explain this trend. Low densities of energy (5 J/cm2) may produce low concentrations of reactive oxygen species, which are considered positive cellular stimulatory effects. 40,41 The results of previous studies are not unanimous with regard to the stimulatory effects on cell and tissue reactions after the use of visible red or infrared laser. 21,36,39 Fibroblasts, cardiomyocytes, smooth muscle cells, endothelial cells, and keratinocytes release VEGF under physiologic conditions or stimulatory effects. 22,42 –44 The influence of laser irradiation on migration, proliferation, adhesion, and survival of these cellular types has been already demonstrated. 19 –22,39,45,46 All explanations included in this paragraph may justify the results obtained after visible red laser irradiation.
Hypoxia may be an alternative theory to explain the increase of expression of VEGF-A165 mRNA after the third day after surgery. 47 The metabolic necessity of oxygen for adenosine triphosphate production leads to blood vessel formation during wound healing. Kipshidze et al. 22 speculated about the ability of LLLT to induct hypoxia due to heat shock protein production and, therefore, the increase of VEGF mRNA expression.
Physiologic concentrations of growth factors such as transforming growth factor beta, endothelial growth factor, and platelet-derived growth factor can induce an increase or decrease in the expression of the VEGF-A mRNA. Tumor necrosis factor alpha can increase the transcription of VEGF receptor 2 gene in endothelial cells; consequently, it stimulates cellular migration and wound healing. 48 Different densities of energy may promote diverse effects on the modulation of these growth factors. Regulation of autocrine pathways of cellular cytokines may be associated with the modulation of VEGF-A165 mRNA. 18
After visible red laser irradiation, means of expression of VEGF-A165 mRNA in group III were always greater than those in group II. Therefore, LLLT seems to influence the expression of VEGF-A165 mRNA on the third, fifth, and seventh days after surgery. Moreover, statistical analysis demonstrated that the levels of expression of VEGF-A165 mRNA of group III were closer to that of a sound tissue than those of group II.
Conclusion
This study demonstrated that the infrared diode laser (GaAlAs, 780 nm) inhibited the expression of VEGF-A165 mRNA during wound healing 1 day after a surgical procedure on the tongue of Wistar rats. The levels of VEGF-A165 mRNA expression after the associated use of the infrared diode laser (GaAlAs, 780 nm) and a visible red diode laser (AlGaInP, 660 nm) were not significantly different from those observed in the control group (II) 3, 5, and 7 days after surgical procedures. Further in vivo studies are needed to explain the mechanisms of action of LLLT, the effects of interaction between two or more distinct lasers, and the clinical results of this therapeutic method on wound healing.
Footnotes
Acknowledgment
The authors thank MM Optics Ltda (São Carlos/SP, Brazil) for kindly providing the Twin Laser equipment.
Author Disclosure Statement
No competing financial interests exist.
Sponsored by Conselho Nacional de Desenvolvimento Científico e Tecnológico, CNPq, Brazil (process # 132325/2005–5).