
Abstract
Introduction
Photobiomodulation is a nonlinear process 5,6 that is well described by the Arndt-Schultz (A.S.) curve. 7 This curve shows that weak stimuli excite physiological activity, whereas strong stimuli have the opposite effect. Visible light can be absorbed by cellular photosensitizers, such as cytochromes, flavins/riboflavins, 8 and NADP. Wavelengths above 500 nm probably stimulate cells by accelerating mitochondrial respiratory electron transfer, as suggested by Karu (2003). 9 A growing body of evidence suggests that cytochrome c oxidase is a key photoacceptor in the red and near-infrared (IR) spectral region. For example, 660–680-nm irradiation has been shown to increase electron transfer in purified cytochrome oxidase, 10 to increase mitochondrial respiration and ATP synthesis in isolated mitochondria, 11 and to upregulate cytochrome oxidase activity in cultured neuronal cells. 12 Although photobiological effects in the near-IR are ascribed to light absorption by mitochondrial cytochrome c oxidase, the low absorbance 13 in this region makes us question it. Lubart et al. (2006) proposed that other mechanisms have to be considered. 14 Santana-Blank et al. (2000) describe that the infrared pulsed laser device (IPDL) has an effect on the water dynamics of the tissues monitored by transverse relaxation times (T 2, I/T 2) obtained through nuclear magnetic resonance (NMR) and this effect might to originate profound effects on biological systems. 15 As is well-known, the solids content of most biological tissues is predominantly protein, and electrostatics affects virtually all aspects of protein structure and activity and is particularly important in proteins whose primary function is to stabilize charge (Rodrigues-Santana et al. 2003). 16
Enzymes undergo structural changes when exposed to light. A low-frequency electric field produces a similar response in enzymes with no absorbent atom in their structures, thereby suggesting that electrical phenomena may stimulate the same mechanism as light. 17 Dian et al. (2002) in their research about conformational dynamics in a dipeptide concluded that the isomerizations induced by the infrared radiation (IR) involve large structural changes, breaking or forming an intramolecular hydrogen bond, or reconfiguring the indole side chain relative to the peptide backbone. 18 However, the molecular mechanism by which LLLT promotes changes in cell functions and structures remains unclear. 19 Although the irradiation dose is one of the most important aspects in LLLT, it is also one that is controversial. The ideal dose is determined based on the literature. The estimated dose depends on the type of tissue to be irradiated and is adjusted for tissue irradiation time and irradiated area as a function of the absorbed energy. 20
The speed of an enzyme-catalyzed chemical reaction depends on four variables: the enzyme concentration, substrate concentration, H+ concentration (pH), and temperature. The presence or absence of cofactors and/or inhibitors will also influence the reaction speed. The possible effects of types of energy other than heat (temperature), such as light, on the activity of the enzymes involved in human metabolic reactions have been scarcely studied. The purpose of the present study was to assess the effect of gallium-aluminum-arsenide (GaAlAs) laser irradiation on α-amylase activity. α-Amylase was chosen mainly because it is easily obtainable and does not require activation by cofactors.
Materials and Methods
Reagents
The following reagents were used: Liquid CNPG amylase (Labtest Diagnostica S.A., Lagoa Santa, Minas Gerais, Brazil) – a system for the determination of α-amylase in biological samples; Control serum, Qualitrol 1H (Labtest Diagnostica S.A., Lagoa Santa, Minas Gerais, Brazil) – a lyophilized preparation in human protein matrix containing various components, including α-amylase.
According to the proposed test, α-amylase hydrolyzes the substrate α-(2-chlorine-4-nitrophenyl)-β-1-4-galactopiranosil maltoside (Gal-G2-α-CNP), releasing 2-chlorine-4-nytro-phenol (CNP) and 1-4-galactopiranosilmaltoside (Gal-G2). The enzyme acts directly on the substrate, releasing more than 90% of chromogen CNP. The speed of formation of CNP was measured by photometry (405 nm) in kinetic mode, and the speed of formation of 2-chlorine-4-nytro-phenol afforded a direct measure of the activity of α-amylase in the sample.
Laser
A GaAlAs laser (λ 904 nm, 50 mW, Ø 0.035 cm2 Endophoton LLT 0107 KLD Biosistemas Equipamentos Eletrônicos Ltda, Amparo, São Paulo, Brazil) was used at energy densities (ED) of 0.01, 0.1, 0.5, and 1.0 J/cm2 in pulsed mode (2 Hz, 100 ns) with sample irradiation time automatically adjusted based on the ED used. All experiments were performed at the Biochemical Laboratory of UNOCHAPECÓ (Chapecó-SC) on a smooth, light work surface at room temperature.
Experimental groups
Analysis of α-amylase was performed in two sample groups: (1) Enzyme (Qualitrol 1H) + substrate (Liquid CNPG) (E + S) − using 0.5 mL substrate and 0.01 mL enzyme for a final volume of 0.510 mL (510 μL); (2) Enzyme +substrate + laser (E + S + L) − using 0.01 mL enzyme, 0.5 mL substrate, and GaAlAs irradiation based on the stipulated parameters.
Group 2 was submitted to four GaAlAs laser-irradiation treatments with increasing doses (ED 0.01, 0.1, 0.5, and 1.0 J/cm2) in six repetitions for each ED. The irradiation time (1, 4, 20, and 12 s) was varied, resulting in irradiation doses of 0.01, 0.1, 0.5, and 1 J/cm2, respectively. A 0.5 mL substrate and 0.01 mL of enzyme were added to a 10 × 100 mm glass tube. The irradiation was performed on the top of the tube that had been placed inside a dark polyethylene tube in order to prevent light scattering during irradiation.
Spectrophotometry
Following the procedures described above, spectrophotometric measurements were performed in a BTS-310 Bio System equipped with a thermostatic chamber capable of precise absorbance measurement at 405 nm.
Statistical analysis
The enzyme percent activity was determined and the Student's t-test at 5% significance (Microsoft® Office Excel 2003) was used in order to compare the experimental and control results and to establish the reliability of the proposed method.
Results
Table 1 displays the mean α-amylase activity for irradiated and non-irradiated samples, the percentage difference for enzyme percent activity (%), and the Student's t-test results.
Values followed by a are over 5.3% and are significantly different from the control.
tcalculated values followed by * denote a significant difference (5%) between the treatments and the control, as determined by Student's t-test mean comparison. There is a mean of six repetitions for each control and energy densities.
The photomodulation of α-amylase activity by GaAlAs laser irradiation (0.01, 0.1, 0.5, and 1 J/cm2) in the conditions described above was analyzed. Figures 1 and 2 illustrate the mean enzyme activity in 95% confidence intervals (Student's t-test) for each irradiation dose analyzed. Variations in enzyme activity with the applied irradiation doses are as follows: 0.01 J/cm2 (0.10%), 0.1 J/cm2 (13.44%), 0.5 J/cm2 (12.57%), and 1 J/cm2 (−6.10%).

Mean enzyme activity for each irradiation dose, followed by the percentage in relation to activity in the control treatment.
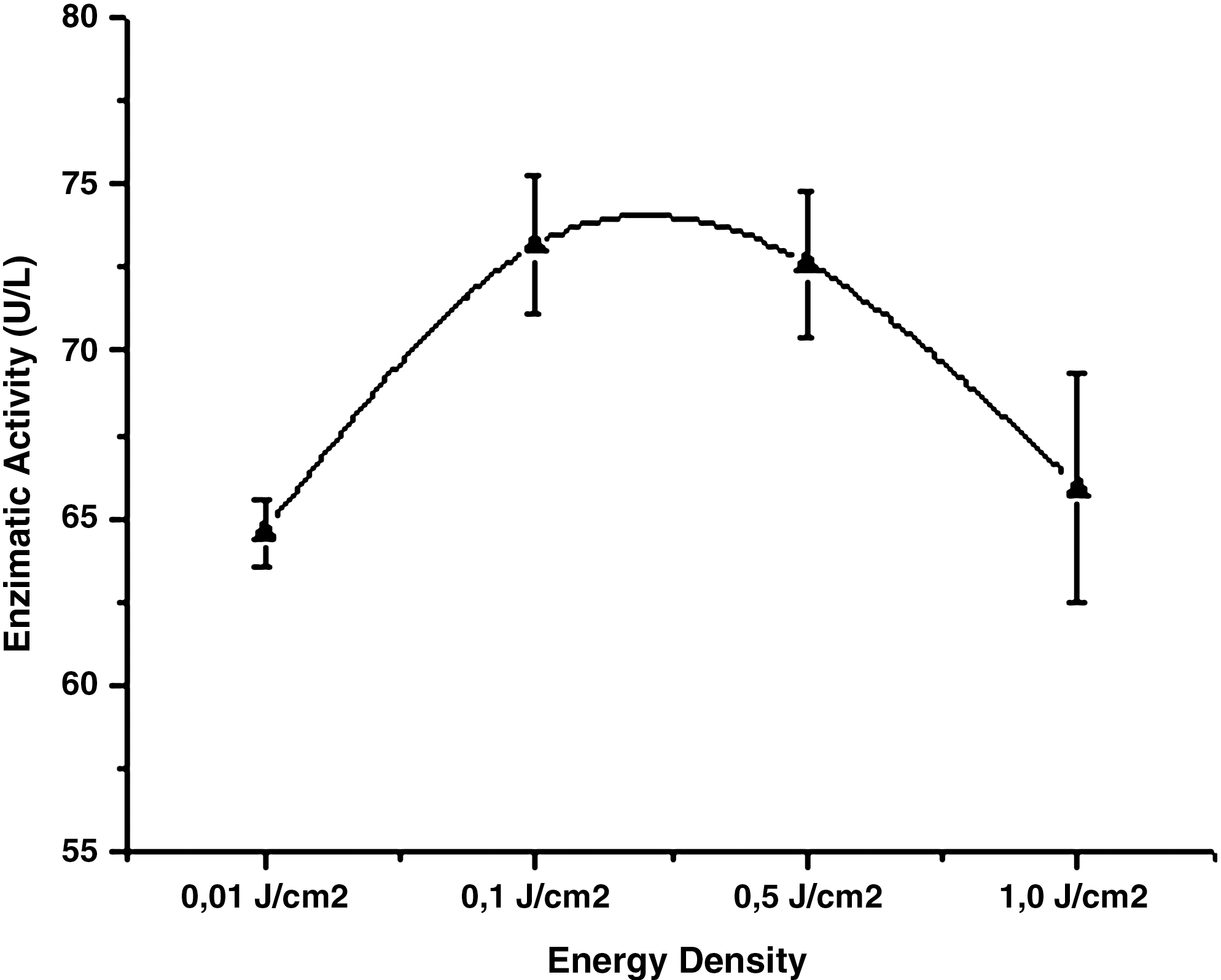
Mean enzyme activity and respective 95% confidence intervals (Student's t-test) for each irradiation dose analyzed.
The results revealed a significant increase for two applied doses, 0.1 and 0.5 J/cm2, as confirmed by the Student's t-test.
Discussion
The use of LLLT has been investigated for decades. Studies have demonstrated that it can stimulate biological processes, particularly in physiotherapy, medicine, and dentistry. Few studies in the literature have addressed the effects of LLLT on enzyme activity.
In the present study, α-amylase was irradiated with GaAlAs laser (904 nm) in order to determine whether it induces any variation in its activity. Based on the data obtained, α-amylase activity increased significantly at 0.1 J/cm2 (13.44%) and 0.5 J/cm2 (12.57%). A similar finding was described by Lombard (1999), who assessed the in vitro effect of He–Ne laser irradiation on isocitrate dehydrogenase activity. 21 Following irradiation at doses of 0.1, 0.2, 0.3, and 0.4 J/cm2, the activity of isocitrate dehydrogenase underwent a statistically significant increase. At 0.3 J/cm2, it increased by 10.02% in relation to the control, a result similar to the findings of the present investigation–the enzyme activity increased and peaked at a specific dose and then gradually declined.
Molecules need to be raised to a high energy level to overcome the energy barrier that exists between the substrate (S) and product (P) for a reaction to occur. This barrier represents the energy necessary for the alignment of the reagent chemical groups, the formation of unstable transient charges, the rearrangement of bonds, and other necessary transformations for the reaction to occur in either direction. The peak of the energy slope, the transition state, is the point at which a drop in the status of either S or P energy levels is equally likely (it is always a downhill path). This state is not associated with a significantly stable chemical species, and it must not be mistaken for an intermediate reaction compound. Laser energy can activate enzymes at the transition state or at the peak of the activation energy barrier, although various hypotheses regarding this mechanism have been proposed. 22
The use of a pulsed laser at 2 Hz may also have contributed to increased α-amylase activity in vitro. Multiple reports suggest that pulsed laser light is an important factor in LLLT. Ueda and Shimizu (2003) found better results using an 830-nm laser with 1- and 2-Hz pulses rather than 8 Hz on mouse medulla cells, but the mechanism of this effect remains unclear. 23
Chen et al. (2004) irradiated germinal seeds of Isatis indigotica Fort with a He–Ne laser (5.23 mW/mm2, 5 min) and microwaves (1.26, 5.23 mW/mm2, 8 s) to determine the effects of these treatments on enzyme activity. They reported a statistically significant increase in the activities of α-amylase (73.3%), total amylase (40.8%), GOT (23%), GTP (40.6%), and proteinase (13.1%) and concluded that the He–Ne laser accelerates cell division and growth in Isatis indigotica seeds. The underlying mechanism of action may stem from light and electromagnetic effects. 24
In 1993, Ostuni et al. conducted an irradiation study of glutamate dehydrogenase with a He–Ne laser. The enzyme activity increased, but the mechanism of activation remains controversial. They reported that the identification of the direct target during the irradiation process was unclear, but suggest that the structure and function of the mitochondrial glutamate dehydrogenase enzyme had been altered by the laser irradiation. 25
Bolognani et al. (1992) report that myosin ATPase previously deactivated by CO2 may be partially reactivated following He–Ne (632.8 nm) and diode (904 nm) laser irradiation. Furthermore, under the appropriate conditions, the irradiated enzyme behaves in an inverse way, enhancing the ATP synthesis through ADP and Pi. The effects were non-thermal (redundant) and there was no absorption at the studied wavelengths. 26
In Fig. 2, the enzyme activity (U/L) according to the irradiation doses (J/cm2) reveals similar behavior with regard to the effects of temperature and pH. In proving experimentally the influence of pH on the enzyme reaction speeds, the curves obtained indicate that the enzymes exhibit an optimal activity pH. As protein conformation partially depends on electrical charges, an optimal pH yields the best catalytic activity and even slight variations can lead to denaturation. Enzymes are also sensitive to temperature. At low temperatures, enzymes are very rigid, and above 50°C their activity drops abruptly. In general, an increase in temperature accelerates chemical reactions: every 10°C rise in temperature doubles the reaction speed. Reactions catalyzed by enzymes follow this general rule. However, like proteins, they undergo heat-induced denaturation when the temperature is too high.
The comparison of the effects of temperature and pH on enzyme activity in relation to the laser-irradiation dose presented in this study suggests that the mechanism underlying enzyme maximal activity and its decrease is related to the primary, secondary, tertiary, and quaternary protein structures of enzymes, as they are essential for catalytic activity. This means that any change in these structures can either increase or decrease enzyme activity. The data in Fig. 2 show a curve similar to the effects of temperature and pH on the enzyme activity, which can be monitored and needs to be investigated further.
Conclusion
Based on the methodology employed in the present study, low-level GaAlAs laser irradiation at 904 nm influences α-amylase activity. For doses of 0.1 J/cm2 and 0.5 J/cm2, the E + S + L treatment yielded statistically significant increases in enzyme activity when compared to the control. The similar curves of the effects of temperature and pH on enzymatic activity observed in this study suggest that laser irradiation also possesses an optimum dose to modulate the enzymatic activity. That is, enzymes have an optimum laser dose (or range) at which their activity is maximal, whereas at a higher or lower dose activity decreases.
Footnotes
Author Disclosure Statement
No competing financial interests exist.