
Abstract
Introduction
Human gingival fibroblasts have been stimulated by LLLT with different wavelengths (660, 670, 692, 780, 786, 809 nm) with an energy density of 2 J/cm2. 18,21 –23 LLLT-irradiated gingival fibroblasts have exhibited prominent rough endoplasmic reticulum, Golgi complex, and exuberant mitochondria. 23
Human dermal (skin) and scar-derived fibroblasts have also been studied after irradiation with red wavelengths of 632.8, 660, and 670 nm, respectively, and energy densities ranging from 0.5 to 180 J/cm2. These findings suggest that LLLT doses in the range of 2–6 J/cm2 enhanced cell proliferation, whereas high doses decreased cell proliferation by showing fewer cells in the S phase of the cell cycle. 24 –26
However, the literature is not unanimous in showing that fibroblast cell activity is enhanced after laser irradiation. Some researchers found no significant difference in fibroblast proliferation with wavelengths of 632.8, 647, 790, and 830 nm in collagen proteoglycan biosynthesis and cell migration and chemotaxis. 27 –29
The seemingly conflicting results may be explained by either nonoptimal LLLT doses or a physiological state in the fibroblast cell where they are not responsive to LLLT.
In in vitro studies, appropriate LLLT dosing depends on numerous factors, such as wavelength, power output, continuous or pulsed emission, power density, irradiation time, dose in J/cm2, total energy delivered, application technique, and intervals between sessions. 30 We had previously reviewed the literature and found a distinct dose–response pattern with a beneficial threshold for power densities started at 5 mW/cm2 and doses above 0.2 J/cm2, whereas power density above 3 mW/cm2 and doses above 4 J/cm2 seemed to inhibit fibroblast activity and fibrosis formation. 31
Research data also point out that the physiological condition of cells may influence fibroblast response to laser therapy. A single irradiation on human embryonic fibroblast results in neither a functional nor micromorphological alteration in the cell surface. 32
Wound-derived fibroblasts have enhanced cell viability and proliferation and higher mitotic activity and are more active in the migration process than normal skin fibroblasts. 24 Irradiated glucose-deprived fibroblast in cultures showed a higher proliferation when compared to nonirradiated controls. 33
The present study was designed to investigate LLLT effects in two metabolically different fibroblast cell lines using identical irradiation parameters and culture media.
Materials and Methods
Laser irradiation procedure
An InGaAlP diode laser (Quasar Medical–Dentoflex, São Paulo, Brazil) with 660-nm wavelength was used for the experiment. For laser irradiation, a support device held the LLLT probe perpendicular to the wells. The emission tip was placed a 2-mm distance from the 96 wells in order to illuminate the entire area of the wells. The laser parameters are given in Table 1.
Cell culture
The murine fibroblast 3T3 cell line was chosen in this study because this lineage has a high metabolic rate and is an immortal cell culture. It was obtained from American Type Cell Culture (Rockville, MD). Murine fibroblast cells were cultured in DMEM (Dulbecco's minimal essential medium) supplemented with 10% fetal bovine serum (FBS), 2 mM glutamine, 100 U/mL of penicillin, and 100 μg/mL of streptomycin sulfate.
Primary human keloid fibroblast cell cultures were obtained from operative biopsy specimens and cultured in DMEM supplemented with 10% fetal bovine serum (FBS), 2 mM glutamine, 100 U/mL of penicillin, and 100 μg/mL streptomycin sulfate.
All cells were maintained at 37°C in a humidified atmosphere containing 5% CO2. 34 These cells were chosen because of their low metabolic rate and limited capacity for growing.
Cell viability (MTT assay)
Cells were plated at a density of 3 × 104 cells/well of both murine fibroblast and keloid primary fibroblast. Ninety-six-well tissue culture plates (Corning Inc., Corning, NY) were used. Cell proliferation and viability were determined by the MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide] method. 1 At the end of treatment, 0.5 mg/mL of MTT in fresh medium was added. After 4 h of incubation, the blue formazan crystals were dissolved with 150 μL of acid-isopropanol, and the absorbance was measured at 570 nm with an ELISA reader (BioTek Instruments, Winooski, VT).
The MTT assay is a laboratory test and a standard colorimetric assay (an assay that measures changes in color) for measuring cellular proliferation (cell growth). This assay is based on the ability of a mitochondrial dehydrogenase enzyme from viable cells to cleave the tetrazolium rings of the pale yellow MTT and form dark blue formazan crystals that are largely impermeable to cell membranes, thus resulting in their accumulation within healthy cells. Solubilization of the cells by the addition of a detergent results in the liberation of the crystals that are solubilized. The number of surviving cells is directly proportional to the level of the formazan product created. When the formazan produced by cells treated with an agent is compared with the amount of formazan produced by untreated control cells, the effectiveness of the agent in causing the death of cells can be deduced through the production of a dose–response curve.
This reduction takes place only when mitochondrial reductase enzymes are active; therefore, conversion is directly related to the number of viable (living) cells. When the amount of purple formazan produced by cells treated with an agent is compared with the amount of formazan produced by untreated control cells, the effectiveness of the agent in causing death of cells can be deduced, through the production of a dose–response curve.
Cell-cycle analysis
The cell cycle, or cell-division cycle, is a series of events that takes place in a cell, leading to its division and duplication (replication). Cell cycle was analyzed by flow cytometry. Adherent cells (105 mL−1) were trypsinized, washed with cold phosphate buffer saline (PBS), and fixed in 70% ethanol at −20°C for at least 1 h. Cells were washed twice with cold PBS, incubated in 0.5 mL of 0.5% Triton X-100/PBS
Fluorescence emitted from the propidium–DNA complex after excitation of the dye was quantitated by FACScan flow cytometry. 23 For each sample, at least 10,000 events were acquired, and the data were analyzed using appropriate software (CELLQuest, Becton Dickinson, San Jose, CA). Cells in S/G2/M and G0/G1 phases and a number of hypodiploid cells were analyzed.
Statistical analysis
One-way ANOVA and Bonferroni's Multiple Comparison Test were used for statistical analyses. A p-value of <0.05 was set for the level of significant difference.
Results
Immortal cultured murine fibroblast
Cell viability
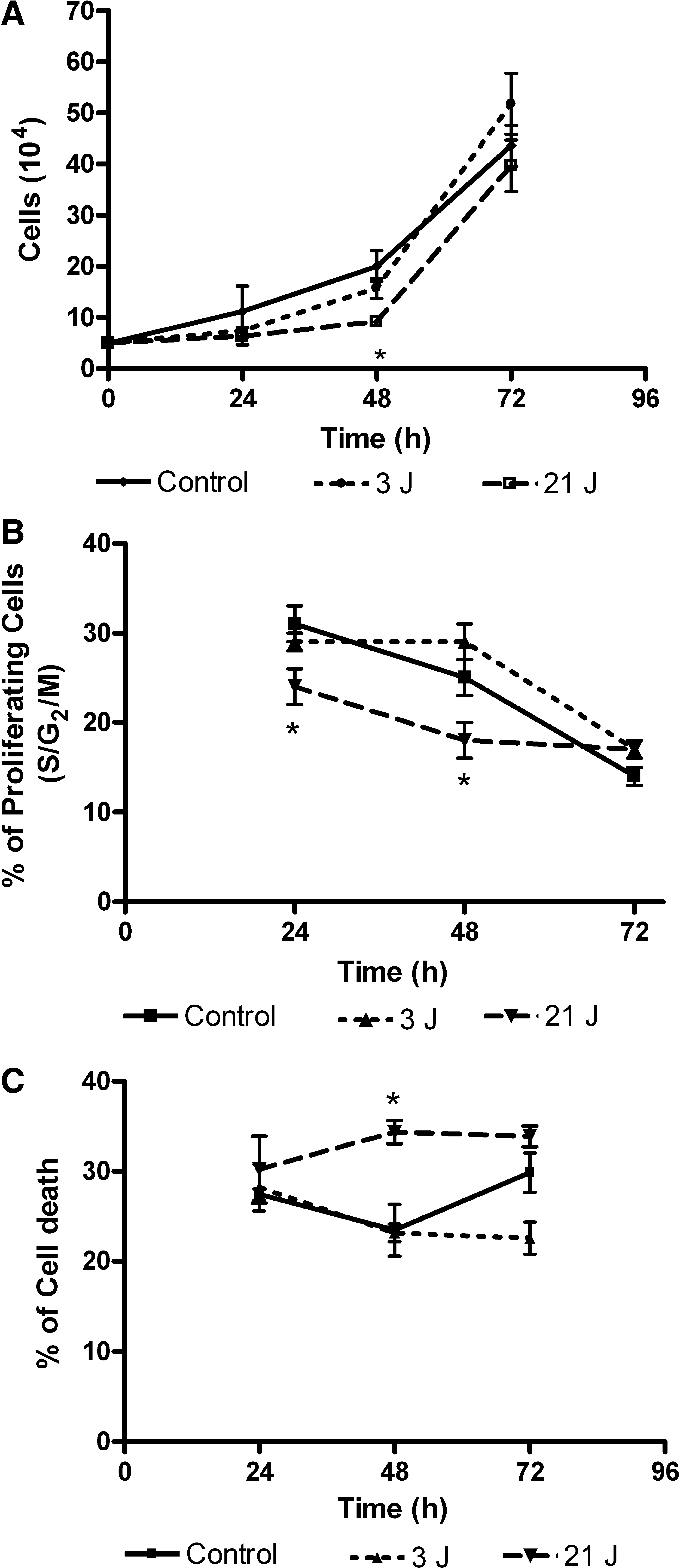
We found no significant differences between irradiated groups (3 and 21 J) or the control group in the trypan blue exclusion test 3T3 for the immortal fibroblast cell line (Fig. 1A), with the exception of the group that received 21 J of energy at 48 h that presented a delay in cell growth after irradiation when compared to the control group.
(
Cell-cycle analysis
Cell-cycle analyses demonstrate that 21 J increased the percentage of hypodiploid cells, probably due to apoptosis, or decreased the percentage of cells under division, whereas the 3-J group showed no significant differences when compared to the control group (Fig. 1B).
Cell-death analysis
The cell-death analyses (hypodiploid cells) after 3 irradiations at 24-h intervals revealed after 72 h that 21 J presented a high number of cell-deaths similar to the control group. However, 3 J of energy was responsible for the smallest number of cell deaths when compared to the control and 21-J groups (Fig. 1C).
Human keloid fibroblasts
Cell viability
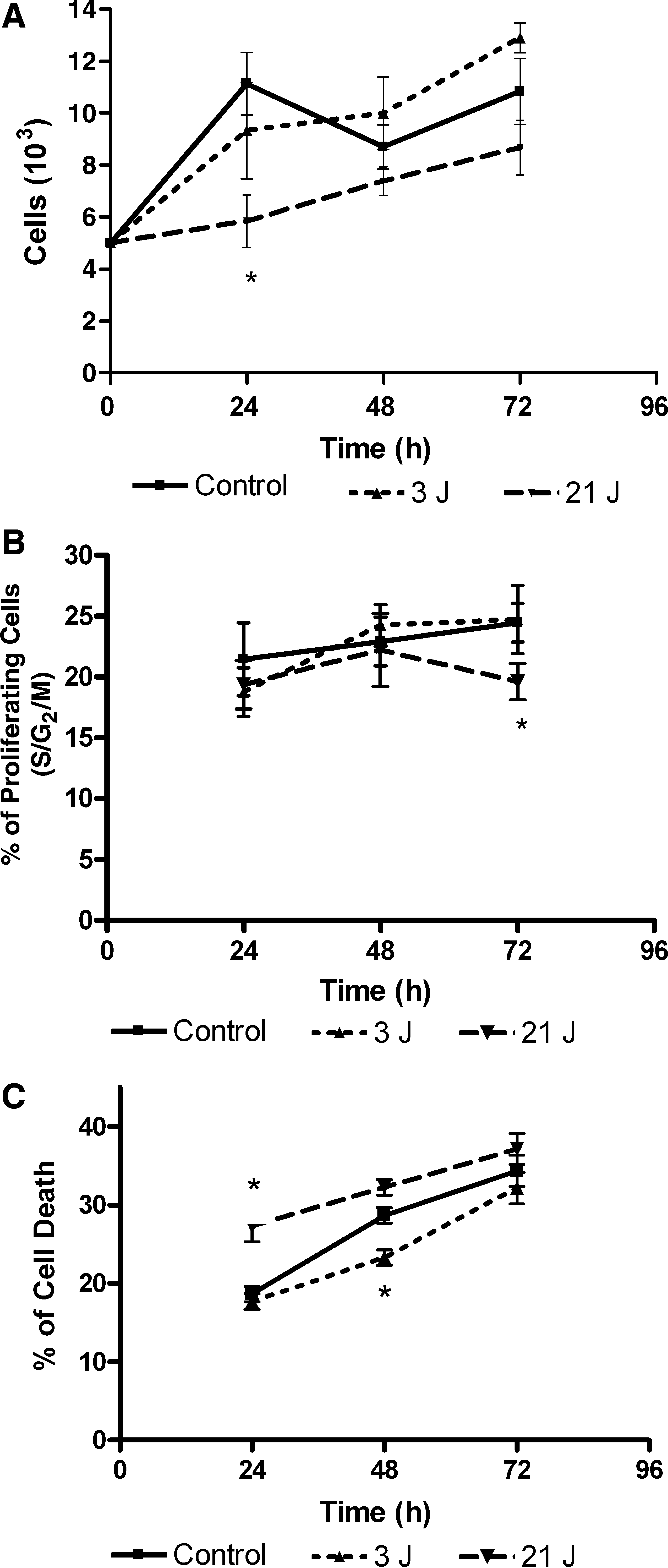
In the primary fibroblast cell culture from surgery keloid tissue, after 3 irradiations in 72 h, the 3-J group showed significantly higher proliferation by MTT test when compared to the other two groups. On the other hand, 21 J showed a smaller number of cells even when compared to the control group (Fig. 2A).
(
Cell-cycle analysis
Cell-cycle analyses of the primary fibroblast cell culture from surgery keloid tissue demonstrate that 21 J increased the percentage of hypodiploid cells, probably due to apoptosis, or decreased the percentage of cells under division, whereas the 3-J group showed no significant differences when compared to the control group (Fig. 2A and B).
Cell-death analysis
The cell-death analyses (hypodiploid cells) after 3 irradiations at 24-h intervals revealed after 24 h that 21 J presented a high number of cell deaths similar to the control group. However, after 72 h, 3 J of energy was responsible for the smallest number of cell deaths when compared to the control and 21-J groups (Fig. 2C).
Discussion
The objective of our study was to compare the effect of low-level laser irradiation (InGaAlP, 660-nm) on different fibroblast cell-line cultures. First we tested the effects of the InGaAIP laser in a cell line of 3T3 immortal fibroblasts with two different doses of LLLT. There was no statistically significant difference in cell numbers between control and the group receiving a dose of 3 J. On the other hand, the group treated with 21 J presented a delay in cell division at 48 h. 24,36,37
Changes in cAMP concentration on fibroblasts have been shown following visible laser irradiation. Recent study has demonstrated that He-Ne laser (632.8 nm) irradiation induces an immediate increase in cAMP of melanoma cell line A2058 cell by enhanced cytochrome c oxidase activity, depending on the energy dose. Pharmacological evidence suggests that activation of A(2) receptors may inhibit collagen synthesis by adenyl cyclase-induced elevation of cellular cAMP. 38,39 This indicates a relationship between growth stimulation of cells and an ability to assist the regulatory mechanism of collagen fibrils in the wound-healing process. This dose dependency can be determined from the dosage used and the time of irradiation. In cell cultures, any changes in the irradiation protocol can induce alterations in the outcome.
The cell cycle, or cell-division cycle, is the series of events that takes place in a cell leading to its division and duplication (replication). Cell cycle was analyzed by flow cytometry. The 3T3 cell cycle analyses by flow cytometry showed a decrease in proliferating cells when cells were irradiated with 21 J after 24 and 48 h. The hypodiploid cells are characteristic for cell death, which in cell cultures probably represents apoptosis, and they increased after 21-J irradiation. Our results are in line with the results of Whang and colleagues, who looked at apoptosis markers and showed that visible red laser with doses of 60 J/cm2 led to cell death by caspase-3. 40
The other fibroblasts evaluated were obtained from human surgery keloids. These cells have a slower growth rate than 3T3 cell lines. The observed effects for the keloid cells were similar, but more apparent than in 3T3. LLLT irradiation with 3 J increased proliferative activity (cell growth) in the MTT test. On the other hand, the propidium iodide (PI) flow cytometric assay revealed a decreased proliferative activity in the group treated with 21 J of energy.
Our results may be explained by the hypothesis that the physiological state of the cell (redox condition) can activate the metabolism of reactive oxygen species (ROS). These are very cytotoxic in higher concentrations, but they can stimulate the cell cycle and protein synthesis in lower concentrations. This hypothesis was discussed by Vladimirov and colleagues, 41 who emphasized that laser light dosimetry is an important part of the phenomenon of cell photostimulation. The authors refer to oxidation–reduction properties as a mechanism that was influenced by light in the cell by chromophores, as a cytochrome–oxidase complex, and consequently changes in the electron transfer rate in the mitochondria. They also suggest the removal of superoxide radicals by superoxide dismutase as another pathway for lipid peroxidation reactions. The concentration of peroxide radicals can damage the cell if the ability to remove these free radicals from the cell is reduced. It can explain the cell death in the high-dose (21 J) group. 41
In clinical studies of chronic tendinopathy, high doses of LLLT have yielded poor results, 42,43 whereas lower LLLT doses have often been associated with positive results. 44 –46 This can be explained by the dose-dependent differences in cellular responses to LLLT found in the current study. In tendinopathy research, an increased focus has been directed toward the recently discovered signs of increased oxidative damage and more apoptosis in fibroblast cells of symptomatic tendons. 47,48 The observed effects in terms of reduced apoptosis in fibroblast cell cultures give an additional rationale for using LLLT in chronic tendinopathy.
Conclusion
In conclusion, our data support the hypothesis that in the cell physiological state, in terms of high metabolic rate and short cell cycle, 3T3 cells are less responsive to LLLT. The biostimulatory effect of LLLT on fibroblast cells is also dose dependent, and a dose of 3 J increased cell proliferation and reduced apoptosis significantly.
Footnotes
Acknowledgment
The authors wish to acknowledge financial support 05/02117-6 provided by FAPESP.
Author Disclosure Statement
No conflicting financial interests exist.