
Abstract
Introduction
Aggregatibacter actinomycetemcomitans is one of the main causing agents of aggressive periodontitis, in a particularly aggressive manner in adolescents. The disease is characterized by the rapid destruction of the tooth-supporting tissues. 1,4 It has also been isolated in cases of bacterial endocarditis and from cerebral and subcutaneous abscesses as an opportunist pathogen. 4
Periodontitis is an inflammatory illness of the tooth-supporting tissues caused by the formation of biofilm by bacteria on the surface of teeth and in the subgingival region. Inflammation leads to the formation of a pocket in the gingival tissues, attachment loss, bone destruction, and finally tooth loss. 5 –7
There is no firmly established protocol or therapy against this aggressive periodontal disease that can consistently stop its advance and keep patients in a stable situation. The most commonly adopted therapy is mechanical removal to break the biofilm and reduce bacterial adherence. This approach is normally associated with the use of systemic antibiotic therapy so as to stop disease development. 8
Although the literature describes a series of antimicrobial substances for local use in the form of gels, biodegradable tablets, and irrigation liquids, none of them have demonstrated relevant clinical results, and no effective products are available today. 9 Also, bacterial resistance to antibiotic therapy is an increasing phenomenon. Development of this resistance can be a consequence of the inadequate use of antibiotics in bacterial infections and their excessive use in the production of meat, garden fertilizers, and cleansing products. In addition, systemic drug administration can have many effects, such as gastrointestinal disturbances, allergies, adsorption of tetracycline by teeth and bones, and alcohol intolerance, among others. 6
Due to increasing difficulties regarding the use of antibiotic therapies, alternative approaches that lead to the removal of the periodontal pathogen from the surface of hard tissue (teeth) have been investigated. 6 There is increasing evidence that photodynamic therapy (PDT) can be a useful tool for antibacterial treatment of aggressive periodontal disease. 7 PDT is based on the principle that a photosensitive substance containing a cellular chromophore can be activated by light and will produce free radicals, ions, singlet oxygen, and other reactive oxygen species (ROS) that have a toxic effect on the cell, leading to cell death. 6,7,10 –12 Many dyes and photosensitive agents, such as methylene blue, toluidine blue, malachite green, crystal violet, and phthalocyanines, among others, have been used in PDT. 4,6,7,10 –12 Many studies have described the use of PDT to inactivate A. actinomycetemcomitans using different dyes, but mainly with laser light devices. For example, Prates and colleagues, 2 in a recent study using a low-power red laser combined with 0.01% (w/v) of malachite green, showed a 99.9% reduction of A. actinomycetemcomitans with a 9-J/cm2 energy dose and 97.2% reduction using a 5.4- J/cm2 dose.
Wilson and colleagues 13 reported that 25 μg/mL of toluidine blue O (TBO) with irradiation from a He–Ne laser (22 J/cm2) promoted a substantial A. actinomycetemcomitans reduction. On the other hand, using 25 μg/mL of methylene blue (MB) and 22 J/cm2, a 64% bacterial reduction was obtained.
Also, Chan and Lai 14 have demonstrated that 267.4 μmol L−1 of MB promoted death in approximately 95% of A. actinomycetemcomitans when using a dose of about 21.2 J/cm2.
Rose bengal (RB), which absorbs light in the range of 500 nm, has been shown to be an excellent singlet oxygen producer when employed as a photosensitizer agent. 15 –17 Preclinical studies with other dyes have already demonstrated that, in adequate doses of drug and light, RB can be safely used to treat skin disorders. 18
Paulino and colleagues 16,17 found that RB, at low concentrations (up to 0.5 μmol) and associated with visible light, promotes cellular death of S. mutans without causing damage to fibroblast cells. This reinforces the choice of using this dye as an active compound in topical PDT protocol. 1
The aim of the present study was to show, for the first time, that the same concept of PDT using RB as a dye for ROS generation under visible light, associated with a classic dental photopolymerizer, could be safely and effectively used as an efficient treatment against A. actinomycetemcomitans colonization.
Materials and Methods
Photosensitizer and light source
The chosen photosensitive dye was rose engal (Sigma, St. Louis, MO). A stock solution (2 mmol L−1) RB was prepared in phosphate-buffered saline (PBS) (0.13 mol NaCl, 2.7 mmol L−1 KCl, 8.1 mmol L−1 Na2HPO4, and 1.47 mmol L−1 KH2PO4, final pH 7.4). This solution was filter-sterilized (0.22 μm) and stored at −20°C.
An odontological resin photopolymerizer was used as the light source, since its drug-absorption spectrum coincides with the irradiated light emission (manufacturer's original filter, 400–500 nm), which allows dye photosensibilization with the available light, as described by Paulino and colleagues. 16,17
Bacterial culture
The A. actinomycetemcomitans JP2 was kindly supplied by Dr. José Francisco Hofling (Faculdade de Odonologia, Universidade de Campinas). The lineage was stored at −20°C in 40% (v/v) glycerol (Sigma, St. Louis, MO), harvested, fractioned, and used (only primary cultures to avoid possible mutations). The liquid medium used to cultivate and check cell viability was either triptic soy broth (TSB, a soybean–casein digest medium; Acumedia, Lansing, MI) or tryptic soy agar (TSA, a soybean–casein digest agar; Difco, Sparks, MD), prepared according to the manufacturer's instructions and autoclaved at 121°C for 15 min prior to use. All cultivations were accomplished by a candle-jar technique, as described by Paulino and colleagues. 16,17 The initial number of colony-forming units per milliliter (CFU/mL) was estimated by measuring the suspension turbidity with a spectrophotometer and verified by using CFU/mL counts on TSA after growth at 37°C for up to 72 h.
Cytotoxic effects of phototherapy on planktonic growing A. actinomycetemcomitans in the presence and absence of light
Before incubation with different RB concentrations, the cells (A600 nm = 0.5) were serially diluted in TSB medium to obtain 103 CFU/mL and distributed (1 mL) in assay tubes (125 × 15 mm). One set of tubes was submitted to a control-cell experiment to evaluate the dye toxicity per se in the dark and the light toxicity per se without any dye. The other set was submitted to a cell-killing experiment with variable RB concentrations ranging from 10 nmol up to 0.5 μmol L−1 under light exposure and from 0 to 50.0 μmol L−1 in the dark.
Light exposure was performed by holding the hand held photopolymerizer (HHP) tip at a 11-cm distance from the cell suspension. Light toxicity per se was first evaluated by irradiating a cell suspension (1 mL) without incubation with the dye, using a light dose of 0.65 J/cm2 (equivalent to a 1-min pulse of light). After that, dye-treated cells (concentration range from 0 to 0.50 μmol L−1) were submitted to different periods of light exposure (from 10 s to 3 min) under a stable irradiation power to promote bacterial inactivation. The contact time with the RB dye before irradiation (from 10 to 30 min) was also studied. Dye toxicity per se (in the dark) was investigated in a similar way, the only difference being that there was no light exposure. The whole process was performed under gentle magnetic stirring.
Cells were grown by spreading 50 μL of the previously diluted cell suspension directly onto TSA, followed by incubation (protected from light) in a candle jar for 72 h at 37°C. After this period, the CFU/mL value was calculated.
Cytotoxic effects of phototherapy on biofilm growing A. actinomycetemcomitans in the presence and absence of light
The biofilm was grown after inoculation in a 24-well cell culture cluster (Corning Costar, 3524, flat bottom, Corning Glass, Corning, NY) (1 mL/well) with 15 μL of a 109-CFU/mL cell suspension and incubation for 24 h in a candle jar. After this period, the medium was removed and the wells were washed with PBS pH 7.4. Each well containing biofilm was then incubated for 30 min with 1 mL of RB solution ranging from 0.01 to 0.5 μmol L−1. After this period, each well was irradiated (1 or 3 min) with a HHP, the dye solution was replaced with 1 mL of TSB medium, and the cell culture cluster was incubated for 24 h in a candle jar. Each well was then washed with PBS, dried at room temperature, and photographed with a Leica DC 300 F digital camera associated with a Leica DMLB light microscope (Leica, Bensheim, Germany) (100 × magnification).
The remaining cells present in the biofilm after the PDT process (or without PDT, as a control) were determined using crystal violet, as described by Kaplan and colleagues, 19 by measuring the optical density at 550 nm of the ethanol–dye solution in each well.
Glycolytic pH-drop assay
The drop in pH caused by A. actinomycetemcomitans biofilm, treated or not with PDT, was monitored as described by Thedei and colleagues. 20
Cytotoxic effects of phototherapy on gingival fibroblasts in the presence and absence of light
The fibroblast's strains were obtained from American Type Culture Collection (ATCC) (CCL-110) as a stock cell line and kept frozen in liquid nitrogen with fetal bovine serum (FBS) (GIBCO, Grand Island, NY) and dimethylsulfoxide (DMSO) (9:1 v/v). After the defrosting process, the fibroblasts were grown in a 24-well cell culture cluster with a Dulbecco's Modified Eagle/Medium F-12 Nutrient medium (DMEM/F12) (GIBCO, Grand Island, NY), prepared according to manufacturer's instructions and filtered through a 0.22-μm membrane. The final medium was enriched with 10% FBS, 100 U/mL penicillin (Sigma, St Louis, MO), 100 μg/mL streptomycin (Sigma), and 0.25 μg/mL amphotericin (Sigma).
The medium was changed twice a week, and fibroblasts were transferred when they reached a confluent population of cells with approximately 1 × 106 cells/mL. This amount of cells was isolated by centrifugation and washed twice with a Hank's Balanced Salt Solution (HBSS) (GIBCO, Grand Island, NY). The initial cell concentration used in all experiments was set up to 0.7 × 106 cell/mL in HBSS. One milliliter of this cell suspension was distributed in assay tubes (125 × 15 mm) to evaluate light toxicity at variable intensities, with or without dye, as previously described for the bacteria.
After the experiments, the cells were isolated by centrifugation, washed twice with HBSS, and placed in DMEM/F-12 growth medium. Afterward, the culture was incubated for 24 h at 37°C in a CO2 incubator. Cell viability was calculated using a Neubauer's chamber based on the trypan blue (Sigma, St. Louis, MO) exclusion test. 21
Statistical analysis
Data are reported as the mean of triplicate measurements of three different preparations, and they were considered to be statistically significant at p ≤ 0.05.
Results
Figure 1 shows the gingival fibroblast viability in the presence of variable RB concentrations, with or without light irradiation (0.65 J/cm2). The dark toxicity test demonstrated that there is no toxicity to the cells below 1-μmol L−1 RB, whereas above this concentration toxicity becomes higher. After light exposure, an exponential toxicity is observed for concentrations above 1-μmol L−1.
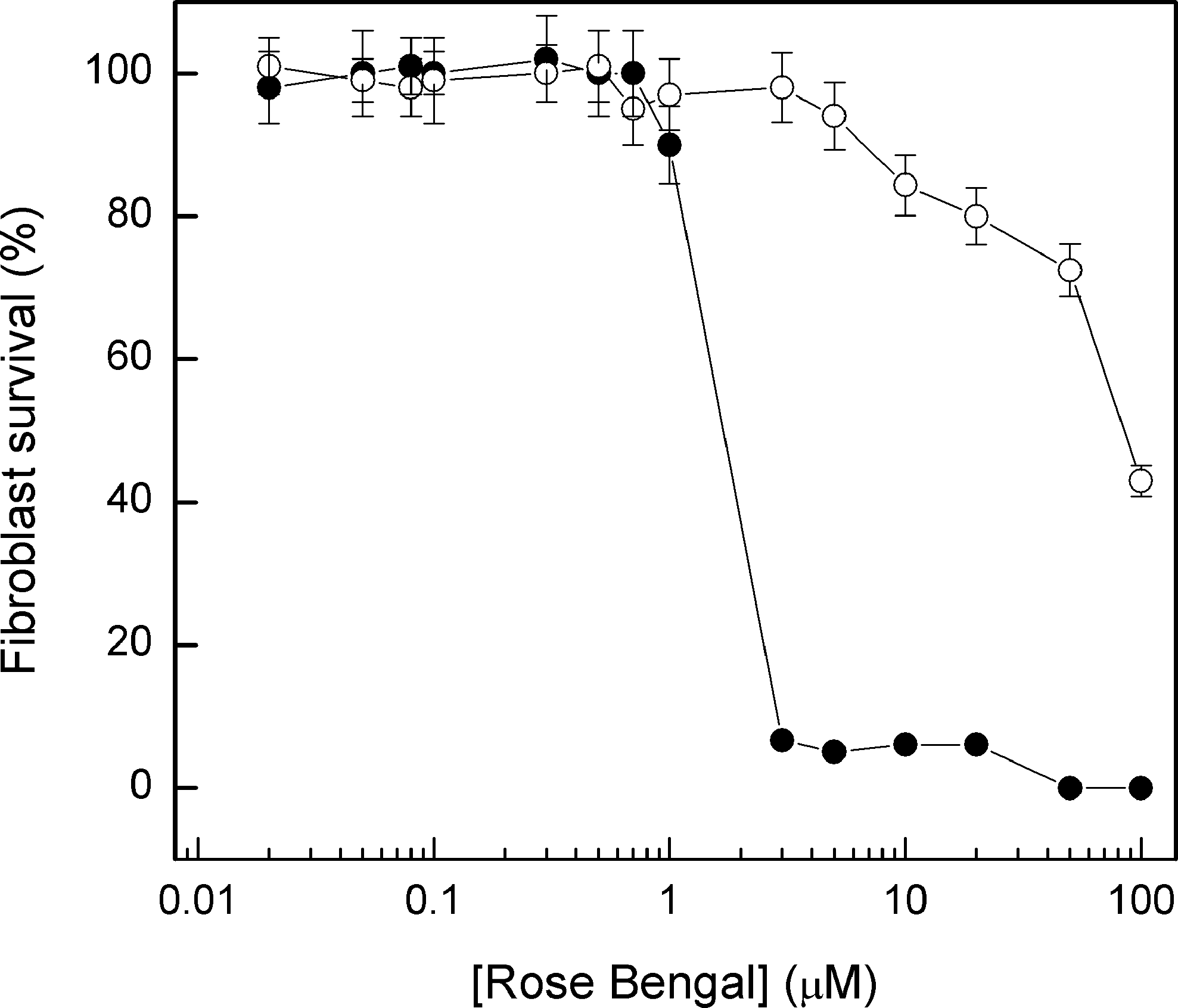
Fibroblast viability in the presence of variable concentrations of rose bengal with (filled circles) or without (open circles) light irradiation (0.65 J/cm2). One milliliter of cell suspension (0.7 × 106 cells) was assayed as indicated, and the cells were isolated by centrifugation, washed twice with HBSS, and cultured in DMEM/F-12 growth medium. Afterward, the culture was incubated for 24 h at 37°C in a CO2 incubator. Fibroblast survival was calculated using a Neubauer's chamber based on the trypan blue, as described in Materials and Methods.
The dark toxicity test using planktonic growing A. actinomycetemcomitans cells showed that RB becomes toxic to this organism in concentrations above 0.1 μmol L−1 and is dose dependent up to 50 μmol L−1, when the cell death induced by the dye itself is close to 100% (Fig. 2). Light exposure, in the absence of RB, does not affect the viability of planktonic growing A. actinomycetemcomitans or fibroblasts in doses up to 1.8 J/cm2.
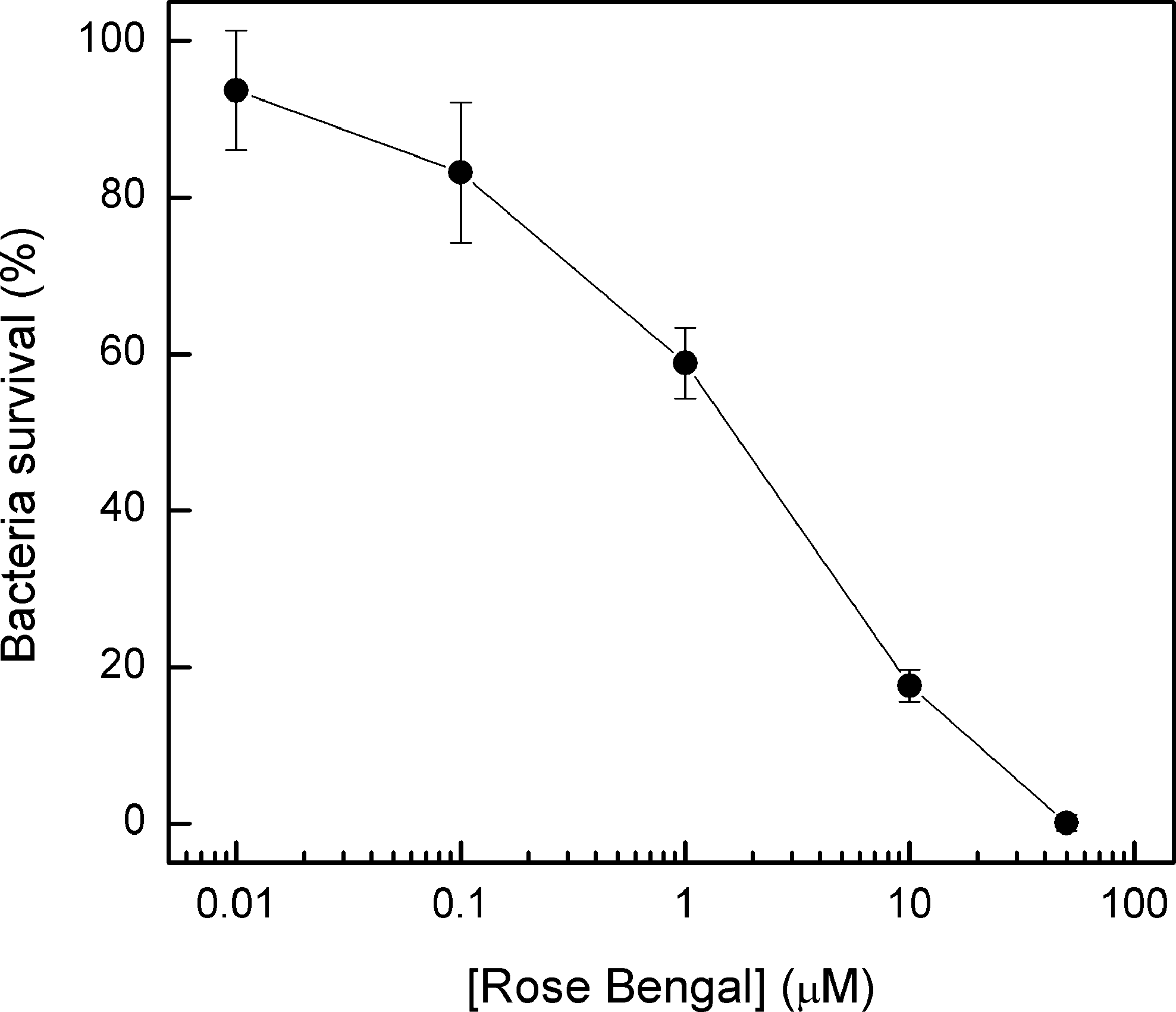
The effect of rose bengal concentration (in the dark) on A. actinomycetemcomitans cell survival. One milliliter (103 CFU) of planktonic growth bacterial cells was preincubated with different concentrations of rose bengal for 10 min and then 50 μL of the suspension was inoculated in TSA medium. The plates were incubated at 37°C in a candle jar for 72 h to determine cell survival.
When RB incubation and light exposure were associated, a dose-dependent effect of dye concentration on A. actinomycetemcomitans death was observed in planktonic cultures. Concentrations as low as 0.01 μmol L−1 were enough to cause 20% cell death, whereas 0.1 μmol L−1 (which presents low dark toxicity) caused 50% cell death after PDT. It was also observed that higher cell death occurred in the first minute of light exposure, with prolonged times (2 or 3 min) causing little additional cell death, for all dye concentrations tested (Fig. 3).

The effect of irradiation time on A. actinomycetemcomitans cell survival incubated with different concentrations of rose bengal. One milliliter (103 CFU) of planktonic growth bacterial cells was preincubated for 10 min with rose bengal, 0.01 (circles), 0.1 (squares), and 0.5 (triangles) μmol L−1, and light irradiated as indicated. Then 50 μL of the suspension was inoculated in TSA medium. The plates were incubated at 37°C in a candle jar for 72 h to determine cell survival.
Figure 4 shows no difference in cell death for both 0.1- and 0.5-μmol L−1 RB after 10 or 30 min of dye preincubation with cells. However, when 1 μmol L−1 of RB is used with 10 min of preincubation, PDT induces 55% death of planktonic cells. At this same RB concentration, but with 30-min contact time with the drug before irradiation, about 90% of the cells die after 1-min irradiation.

The effect of preincubation time of rose bengal on PDT of A. actinomycetemcomitan cell survival. One milliliter (103 CFU) of planktonic growth bacterial cells was preincubated for 10 (open bars) or 30 min (filled bars) with different concentrations of rose bengal and a light dose of 0.65 J/cm2. Then 50 μL of the suspension was inoculated in TSA medium. The plates were incubated at 37°C in a candle jar for 72 h to determine cell survival.
Figure 5 shows that A. actinomycetemcomitans forms a continuous biofilm at the bottom of the well. This biofilm is not affected by RB or light alone. Following PDT, there is a significant reduction in the biofilm, which depends on RB concentration and irradiation time using this dye as the ROS generator. The higher the RB concentration used for PDT, the more affected was the biofilm, leading to its detachment from the bottom of the well. PDT strongly reduces the acidogenic capacity of A. actinomycetemcomitans biofilm (ΔpH = 0.1 ± 0.05) compared with biofilm treated with RB or light alone (ΔpH = 0.75 ± 0.05). In addition, a reduction of crystal violet incorporation (dose and RB dependent) was observed, which reflects a reduction in the biofilm extension of approximately 45% (Fig. 6). This result confirms the stereomicroscopic analysis shown in Fig. 5.

Micrography showing the architectures of the biofilm formed by A. actinomycetemcomitans (100 ×) after growth during 24 h in a candle jar, as described in Materials and Methods, undergoing different treatments. The relative intensity of the images is shown: (

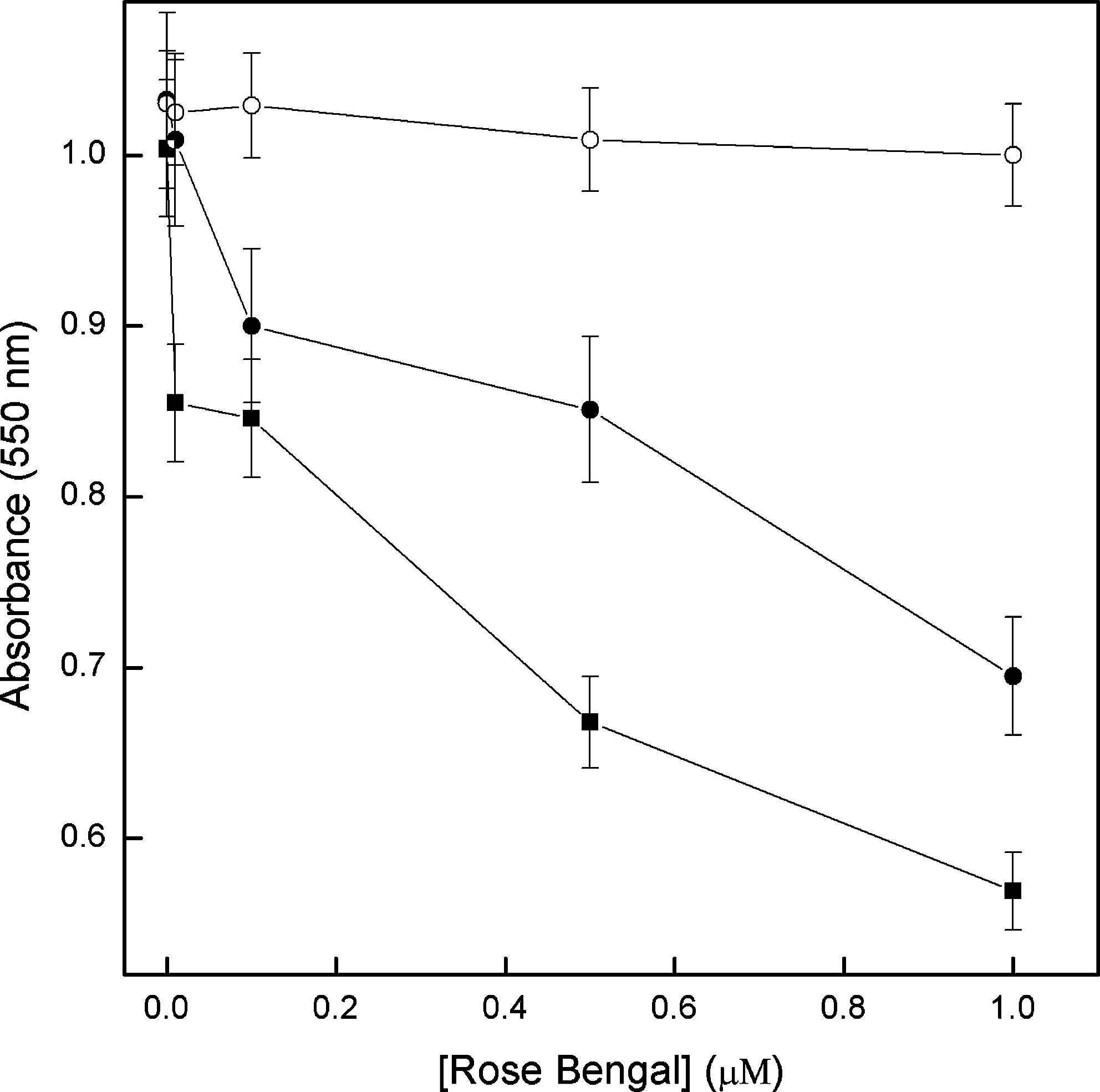
The effect of PDT on A. actinomycetemcomitans biofilms after growth during 24 h in a candle jar. The previously formed biofim was incubated with different concentrations of rose bengal during 30 min, as described in Materials and Methods: (open circle) without photoirradiation, (closed circle) 1 min, and (closed square) 3 min of photoirradiation. The plates were incubated at 37°C in a candle jar for 24 h to quantify the remaining cells in biofilm by crystal violet methodology, as described in Materials and Methods.
Discussion
Rose bengal is a photosensitive agent that is routinely used as a model compound for ROS generation in PDT studies, but it has not been employed in clinical trials. However, if used in lower concentrations for topical treatment, it could be feasible as a protocol to treat some significant dentistry diseases. 16,17 This agent locates itself in the plasmatic membrane of mammals, forming high quantities of singlet oxygen (φ1O2 = 0.70) after visible light irradiation, thus leading to cell death. Since the effective and toxic concentrations are close, therapeutical use has been avoided. 18,22 In addition, RB presents a light absorption band in the 500- to 600-nm range, which is not compatible with the therapeutic windows used in clinical trials (between 600 and 800 nm). 23
In PDT, the dye should not cause cell damage, that is, without light exposure; an ideal dye has no toxic effect on the cells, and the conditions used for the photoinactivation of any pathogen with PDT (light dose and dye concentration) should not affect the neighboring human tissues either.
The results shown in Fig. 1 clearly demonstrate that gingival fibroblasts are not affected in the conditions used to photoinactivate A. actinomycetemcomitans (up to 0.5- μmol L−1 RB, 10 min of preincubation before PDT, and 0.65 J/cm2), as also observed by Mousavi and colleagues, 4 who demonstrated that low RB concentrations (5–10 μmol L−1) per se do not kill either epithelial cells or gingival fibroblasts. On the other hand, verification of RB toxicity per se for the A. actinomycetemcomitans bacterial lineages reveals high toxicity for concentrations over 1 μmol L−1 (Fig. 2). Hence, we have chosen to work with the 0.01-, 0.1- and 0.5-μmol L−1 concentrations (non-, slightly, and moderately toxic to A. actinomycetemcomitans, respectively). In all the studied conditions, an increase in light quantity results in a decrease of bacterial viability (Fig. 3), which clearly indicates that the compounds produced by RB photoactivation are responsible for planktonic cell death.
Similar results have been reported by other authors, who used different dyes, light sources, and organisms. Bhatti and colleagues 25 used toluidine blue and a He–Ne laser (0.88 J/cm2) to photoinactivate Porphyromonas gingivalis, which together with A. actinomycetemcomitans is one of the periodontal pathogens. Paulino and colleagues 16,17 achieved photoinactivation of S. mutans using a similar light source (about 0.5 J/cm2) and rose bengal as the photosensitive dye.
Paulino and colleagues 16,17 showed that death of S. mutans induced after PDT was also obtained without previous incubation with the dye, demonstrating that toxicity to these cells is probably due to the reactive species produced in the extracellular medium and not exclusively by those generated in the intracellular medium. But this may not occur with A. actinomycetemcomitans, which is a Gram negative bacterium presenting a different surface and membrane structure. For this organism, some contact time with the dye, before irradiation, is required (Fig. 4), probably to enable interaction of RB with the cell wall and even to reach the periplasmic space, plasmatic membrane, or the accumulation in the cytoplasm. In fact, 30 min of contact with the dye before irradiation is the most efficient period for A. actinomycetemcomitans inactivation for all RB concentrations used.
In Fig. 4, it can also be observed that, when 1.0 μmol L−1 of RB is used, there is significant variation in the bacterial viability after PDT of cells pre incubated for 10 or 30 min, because high concentrations of dye can form aggregates, decreasing dye efficiency and requiring additional time to reach the place of effective action. The photochemical and photophysical characteristics of rose bengal (singlet oxygen quantic yield, fluorescence emission, transient absorption, triplet decay, phosphorescence emission, and quantic yield for ROS production) are well described, 26,27 showing that, in specific experimental conditions, this dye could form a dimer or another superior aggregate, which results in a broad absorption spectrum and changes in the fluorescence properties, thus reducing ROS production significantly. 28 Today, some studies using the dye-aggregate delivery system could improve the response and increase the photoactivity in PDT studies. 29
Many works have demonstrated that bacteria of the oral cavity grown in planktonic media are sensitized by PDT. Microorganisms that cause oral disease are organized in biofilm, which presents some different characteristics from those observed in planktonic growth, such as the presence of extracellular polymeric substances (EPS), as well as different cell wall composition, growth, metabolic activity, and gene expression. 30 The bacteria grown in the biofilm are more resistant to antimicrobial agents, and the use of PDT has provided the significant film reduction.
Since smooth or rough strains of A. actinomycetemcomitans also grow in biofilm, it is essential to know whether PDT is effective against this pathogen growing in this form. Our results strongly suggest that the biofilm is affected by PDT since, after this treatment, some clear areas without biofilm appear where a homogeneous biofilm appeared prior to PDT (Fig. 5). More evident is the dependence on RB concentration and irradiation time of the biofilm disarticulation. When crystal violet was used to quantify biofilm (Fig. 6), structured cell reduction on the plate (showed on Fig. 5) of around 45% was confirmed (dose and RB concentration dependent). In addition, the acidogenic capacity, due to formation of acid end products generated mainly by carbohydrate fermentation, is characteristic of this bacterium. Also, the acidogenic capacity is a measure of the cell's metabolism and, by extension, of its viability. Since no acidification is observed after PDT in biofilm, it can be suggested that, at least superficially, the bacteria have died. These results, associated with those in Figs. 5 and 6, show that PDT terminates the acidogenic capacity of A. actinomycetemcomitans biofilms, giving support to this therapy of bacterial metabolism. 11,20
Besides, when the per se toxicity of RB or of the light applied to a nondye- treated suspension was evaluated, results similar to the controls were observed, indicating that the decrease of the acidogenic capacity of A. actinomycetemcomitans is due to the combination of the dye treatment followed by irradiation (which generates oxygen reactive species) and not alone by dye or the light applied.
The efficacy of PDT against biofilms has been widely studied. Zanin and colleagues 30 verified that PDT using 100 mg/L toluidine blue with a He–Ne laser or light-emitting diode (LED) (49 and 294 J/cm2) provides 99% reduction in S. mutans biofilm. Wilson and colleagues 31 showed that the biofilm viability of Streptococcus sanguis is reduced by PDT using a galliun aluminium arsenide (GaAlAs) diode laser light (4.1 J/cm2) combined with aluminum dissulfonated phthalocyanine (AlPcS2) as photosensitizer. Wood and colleagues 32 reported that significant killing occurred when biofilm was treated with a cationic Zn(II) phthalocyanine photosensitizer and exposed for 30 min to a 400-W tungsten-filament lamp.
Wood and colleagues 10 compared the use of three different photosensitizing agents: erythrosine (ERY), Photofrin, and methylene blue (MB) at a 22 μmol L−1 for each dye concentration to photosensitize the S. mutans biofilm, using a 400-W tungsten lamp with a light intensity of 22.7, 22.5, and 22.5 mW/cm2 for each dye, respectively. ERY was more effective than Photofrin or MB, reducing S. mutans biofilm in 48%, compared with MB (41%) and Photofrin (just 0.04%).
Finally, Metcalf and colleagues 33 have also verified the PDT effect on biofilm formed by S. mutans using 22 μmol L−1 of ERY and a light dose of 6.75 J/cm2: it induced 57% of biofilm cell reduction.
Conclusion
In the present study, we can observe that an important periodontopathogenic bacterium, which causes bacterial endocarditis, can be photoinactivated by the light coming from an important odontological resin photopolymerizer, together with RB as the photosensitizer dye. The RB optimum concentration that efficiently kills A. actinomycetemcomitans using PDT (0.5 μmol L−1) is out of the toxicity range for gingival fibroblasts, which could lead to a clinically safe protocol. At higher concentrations, it presents some dark toxicity to A. actinomycetemcomitans itself.
Other dyes that belong to the new generation of photosensitizer compounds are now under photophysical and photochemical evaluation to be used in clinical trials for the treatment of many other odontological diseases. They have been tested for local antimicrobial therapy in periodontal treatment with satisfactory effects and are a promising therapy for periodontal disease treatment.
The photodynamic technique might be an option for clinical treatment once improvements are made to reduce the side effects on patients, although we are not suggesting it can replace antimicrobial chemotherapy. It is clear that additional experimentation has to be made with the use of different dyes, gel suspensions, and light (LED and laser), in vivo and in vitro, to establish a photochemotherapeutic system by which its effectivity can be studied.
Footnotes
Acknowledgments
We thank Cynthia M. de Campos Prado Manso and Priscila Cerviglieri for linguistic advice. We also thank Fundação de Amparo a Pesquisa do Estado de São Paulo, Conselho Nacional de Pesquisa, and Universidade de Uberaba, for the continuous support given to our laboratories. Rosangela de Goulart and Mayte Bolean are recipients of a doctoral and master fellowship from CAPES-Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, respectively.
Author Disclosure Statement
No competing financial interests exist.