
Abstract
Heat-shock proteins (HSPs) are indicative of stressing conditions that may affect cell viability. In Streptococcus mutans, acid stress induces high levels of GroEL, an HSP, in addition to metabolic alterations, as shown by proteomic analysis.
Introduction
A number of methods have been developed to control bacterial proliferation in mouth environments, including (a) vaccine production 15 ; (b) fluoride therapy, 16 and (c) antibiotic therapy. 17 Studies using PDT have been routinely used in bacteria, 18 yeasts, 19 and viruses, 20 including HIV 21 inactivation, to search for new strategies against microorganisms.
PDT is based on the principle that a photosensitive substance can be activated by light and produce free radicals, ions, and singlet oxygen, among other substances that have a toxic effect on the cell, leading to cell death. 22 –26 These toxic substances might be acting in different cell locations, causing cell wall/plasma membrane damage, leading to cytoplasm leakage and nucleic acid strand breakage, among others. 27,28 Rose bengal (RB) has been shown to be a good singlet oxygen producer when used as a model photosensitizer agent, presenting a light absorption in the range of 500 nm region. 29 –31 Preclinic studies with other dyes have already demonstrated that, in adequate doses of drug and light, it can be safely used to treat skin disorders. 32 It was found that RB, at ideal concentrations and associated with the use of light, promotes cellular death of the S. mutans bacteria without causing damage to the fibroblast cells. 30,31
Previous work of Dahl et al. 33 reported a comparison of PDT effects (by using a tungsten lamp) and RB (5 μmol/L), resulting in death of gram-positive species about 200 times more quickly (99% inactivation) than with gram-negative species. Also, Goulart et al. 34 showed the use of odontologic resin photopolymerizer and RB (0.1 μmol/L), resulting in death of Aggregatibacter actinomycetemcomitans, in both planktonic and biofilm cultures, not causing damage to fibroblast cells under the same conditions.
Heat-shock proteins (HSPs), also called stress proteins, were discovered after thermal-shock studies and are present in different organisms. 35 The better-known ones are HSP-60 (also called GroEL, with ∼60 kDa molecular mass) and HSP-70 (also called DnaK, with ∼70 kDa molecular mass). Previous studies showed that S. mutans responds to stress with the induction of general and stress-specific proteins. 36 In this organism, the expression of HSP may be stimulated by acid exposure 37 and inhibited by xylitol in xylitol-sensitive strains, whereas xylitol-resistant mutans showed no alteration in HSP synthesis after exposure to this polyol. 35 Taken together, these results show that, in S. mutans, the expression of HSP is evidence that indicates the existence of a condition that can compromise cell viability, allowing the supposition of metabolic alterations in the bacteria.
In the present study, we evaluated the PDT effect on S. mutans with RB as a photosensitive dye and a hand-held photopolymerizer (HHP) as a light source in a condition to induce partial bacteria inactivation to study whether this treatment results in the expression of GroEL protein and DNA damage.
Materials and Methods
Photosensitizer and light source
A stock solution of rose bengal (RB) from Sigma Chemical Co. (St. Louis, MO) in 2 mmol/L saline phosphate buffer (PBS), pH 7.4 (Na2HPO4 8.1 mmol/L; KH2PO4, 1.47 mmol/L; NaCl, 0.13 mol/L; and KCl, 2.7 mmol/L) was prepared. This solution was filter sterilized (0.22 μm) and stored at −20°C. An odontologic resin photopolymerizer (HHP) from Dabi Atlante SA (Ribeirão Preto, SP, Brazil) was used as a light source with the following characteristics: 350–500 mJ/cm2 of light emitted at a wavelength range of 400–500 nm, as described by Paulino et al. 30,31
Strain and bacterial culture
The ATCC 25175 strain of S. mutans was purchased from Fundação André Tosello (Campinas, SP, Brazil). Bacterial cells were kept stocked at −20°C in water with 40% glycerol (vol/vol) or in a candle jar for frequent use with weekly subculturing on Tryptic Soy Agar (TSA-Soybean-casein digest agar) from Difco (Sparks, MD). Large amounts of cells were obtained from 1-L cultures of complete medium incubated in a candle jar (obtained after lighting a candle inside the jar to consume part of the oxygen) under magnetic stirring, as described by Magalhaes et al. 11 and grown until the pH reached 4.2. The liquid media used to cultivate and check cell viability was a Tryptic Soy Broth (TSB-Soybean-casein digest medium) from Oxoid or Tryptic Soy Agar (TSA) medium, prepared according to the manufacturer's instructions and autoclaved at 121°C for 15 min before use. For microbial assays, the initial turbidity of the bacteria culture was adjusted to A600nm = 0.5 (∼109 cells/mL) by using a spectrophotometer (Milton Roy, Spectronic, Genesys 2). Then 15 μL of the stock was inoculated in “Falcon tubes” (15 mL) containing 4 mL of previously autoclaved TSB medium. The number of colonies forming units per milliliter (CFU/mL) was estimated by measuring the suspension turbidity with a spectrophotometer and verified by using CFU/mL counts on TSA after anaerobic growth at 37°C for 36 h. 30,31
Photosensibilization of S. mutans by rose bengal and stress procedure
S. mutans was grown as described and diluted in TSB medium to get 103 CFU/mL. This bacterial suspension (200 mL) was preincubated with 0.1 μmol/L RB for 10 min and then photoirradiated with HHP (at a 11-cm distance from the light source), with gentle stirring. The irradiation was done during four cycles of 30 s each that correspond to 1.3 J/cm2 or 12.8 mW (no change in temperature was observed after irradiation). 30,31 After 5 min, 4 mL of 1 mmol/L chloramphenicol ethanolic solution was added to all samples. Then samples were centrifuged for 30 min at 4,000 g to remove the culture medium, and the pellet was used for protein or DNA extraction. Controls were done in the absence of light, in the absence of dye, and without treatments. For the stress procedure, a sample was preincubated for 10 min with 11.7 g of NaCl and 20 μL of H2O2 30% (vol/vol) with gentle stirring and processed in the same way.
Cell viability
The number of viable cells in both controls and PDT-treated samples was determined with the standard pour-plate method. Sequential dilutions of cell aliquots in sterile 0.9% NaCl (wt/vol) were mixed in brain/heart infusion agar, poured into Petri dishes, and incubated in candle jars at 37°C for 48 h. The number of colonies of S. mutans was then estimated with a light stereomicroscope.
Glycolytic pH-drop assay
Suspension of S. mutans (with or without the PDT process) were harvested, washed (50 mmol/L KCl and 1 mmol/L MgCl2) once by centrifugation, and resuspended in the same solution. Suspensions were titrated with KOH to a pH of ∼7.2. Then glucose was added to an initial concentration of 50 mmol/L, and the resulting decrease in pH was monitored with a thin-bulb glass pH electrode, reflecting metabolic activities of both samples. 38
Protein extraction
The pellet obtained after different treatments was washed 3 times with PBS (pH 7.4) containing 0.1 mg/mL of chloramphenicol. To the final pellet, 2.0 mL of cleavage buffer (8 mol/L urea, 62 mmol/L DTT, and 2% (vol/vol) Igepal), and 1.0 g of glass beads for each gram of cells were added. Finally, the samples were sonicated by using a tip sonicator for 5 cycles of 30 s, with 1 min of interval (37% amplitude, 220 watts) in an ice bath and vortexed for 10 min. The cell lysate was centrifuged for 15 min at 6,400 g, the protein concentration estimated as described by Read and Northcote, 39 and the supernatant used to assay HSP expression by Western blot.
SDS-PAGE and Western blot
SDS-PAGE of proteins extracts (30 μg) was done by using a method described by Laemmli. 40 Five percent polyacrylamide was used for the stacking gel, and 7%, for the running gel (100 × 105 × 1 mm). The gels were silver stained and β-galactosidase (116 kDa), phosphorylase B (97.4 kDa), bovine serum albumin (66 kDa), egg albumin (45 kDa), and carbonic anhydrase (29 kDa) were used as molecular markers.
After SDS-PAGE, proteins were electrophoretically transferred to a nitrocellulose membrane by using 15 mA, 200 V, for 2 h. The membrane was blocked with a blocking buffer [40 mmol/L Tris-HCl, pH 7.4, containing 5% (wt/vol) nonfat milk, 1% (wt/vol) BSA, and 0.1% (vol/vol) Tween 20], reacted (2 h) with the antibody against GroEL (Sigma) diluted 1:1,000 (vol/vol) in the blocking buffer and washed with a washing buffer (40 mmol/L Tris-HCl, pH 7.4, containing 0.3 mol/L NaCl) for 30 min with three buffer changes. Then the membrane was incubated with peroxidase-labeled goat anti-rabbit IgG conjugate (GAR, Sigma) diluted 1:1,000 (vol/vol) in the washing buffer for 1.5 h. The substrate (0.7 mg/mL) 3,3'-diaminobenzidine (Sigma) was used to reveal protein–antibody reactions.
DNA extraction
DNA was extracted by using the method described by Pitcher et al. 41 S. mutans cultures, previously submitted to different treatments, were harvested by centrifugation at l,000 g for 15 min. The cell pellet was resuspended in fresh lysozyme (50 mg/mL) in Tris EDTA buffer (10 mmol/L Tris-HC1; 1 mmol/L EDTA, pH 8.0), and the suspension was incubated at 37°C for 30 min. Cells were lysed with 0.5 mL 5 mol/L guanidium thiocyanate (Sigma), 100 mmol/L EDTA, and 0.5% vol/vol sarkosyl. After cooling, 5 ml of 10% vol/vol sarkosyl was added, filtered through a 0.45-μm Nalgene filter (BDH, Ltd.), and vortexed. The lysates were cooled on ice, and 0.25 mL cold 7.5 mol/L ammonium acetate was added with mixing, held on ice for a further 10 min, and then a 0.5 mL chloroform and 2-pentanol (24:1) mixture was added. The phases were mixed thoroughly, transferred with a wide-bore Pasteur pipette to a 1.5-mL Eppendorf tube, and centrifuged (25,000 g) for 10 min. Supernatants were transferred to Eppendorf tubes, and 0.54 volumes of cold 2-propanol were added. The tubes were inverted for 1 min to mix the solutions, and the fibrous DNA precipitate was deposited by centrifugation at 6,500 g for 20 min. Pellets of DNA were washed 5 times in 70% ethanol and dried under vacuum. DNA samples were redissolved overnight at 4°C in a small volume (usually 100 μL) of sterile, deionized water. The integrity of the DNA (10-μL samples) was checked by horizontal gel electrophoresis in 0.5% agarose in Tris Borate EDTA buffer (89 mmol/L Tris, 89 mmol/L boric acid, 2 mmol/L EDTA, pH 8.3) containing ethidium bromide (0.5 mg/mL).
Densitometry
The intensity of the protein bands of stained gels and of the Western blot were determined by using the equipment and program from Spectronics Corporation ImageAide, file version: 3.06.04; the background of each sample was discounted. Three proteins present in the extract were used as control because their intensity did not change >3%.
Statistical analysis
Data were reported as the mean (±SD) of five measurements of three different experiments and were submitted to analysis of variance in a completely randomized design. Differences between means were verified by the Tukey test (p < 0.05).
Results
The pH variation profile of a cell suspension with and without PDT (0.5 J/cm2) treatment is shown in Fig. 1. Results show that a cell suspension of S. mutans is able to acidify the external environment after a glucose pulse. This acidification begins just after glucose addition, and after 10 min, a ΔpH of ∼0.5 pH units is observed. Figure 1 also shows that, after PDT with 0.1 μmol/L of RB, the acidogenic capacity was almost totally suppressed and resulted in ∼25% cell death, as measured by pour-plate method. The same ΔpH of acidification is observed when light is used alone and when dye was used without light irradiation (data not shown). After PDT with 1 μmol/L of RB, 100% cell death was observed, and the cell suspension totally lost the acidification capacity. Other sublethal conditions (<0.1 μmol/L of RB) did not show significant GroEL expression.

pH of cell suspension of Streptococcus mutans: (•) control (without PDT process) and (○) after PDT process. Each point corresponds to an average of five independent experiments done with different bacterial cultures.
Figure 2A shows the electrophoretic profile of S. mutans cell extracts submitted to different treatments. The silver-staining procedure used did not reveal any detectable difference in protein profiles of extracts obtained from cells submitted to PDT as compared with control experiments. Conversely, Western blot using antiGroEL antibodies showed that S. mutans expresses this protein in all conditions tested with different intensity, because of the identification of a 70-kDa protein band (Fig. 2B).
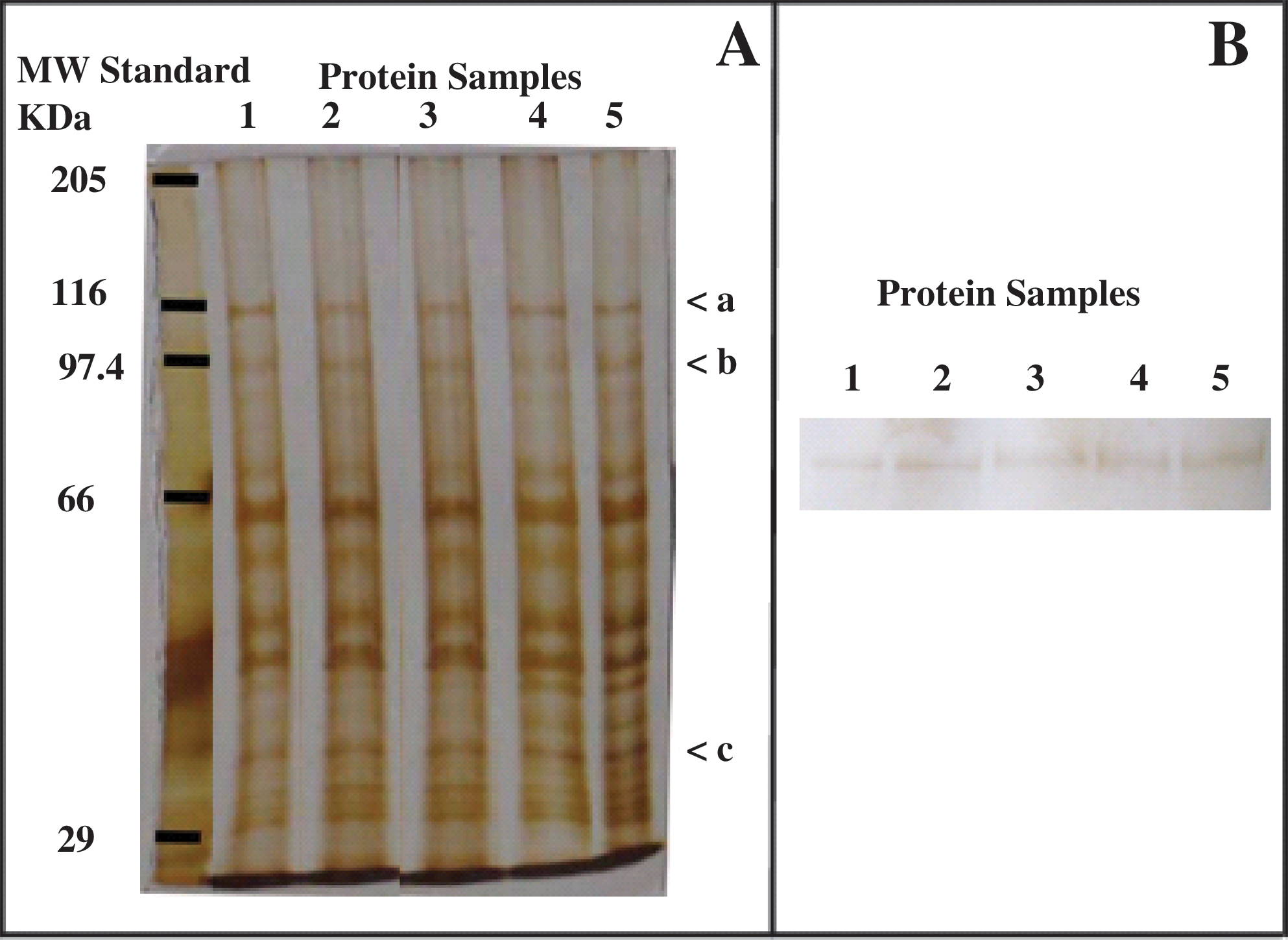
Comparison of the protein profiles of cellular extracts from Streptococcus mutans submitted to different treatments: (1) negative control, bacteria without any treatment; (2) bacteria treated with 0.1 μmol/L of RB kept in the dark; (3) bacteria submitted to photoirradiation (without RB); (4) PDT process; and (5) positive control, bacteria submitted to saline stress. (
The quantitative analysis with Western blot by densitometry shows that PDT induces GroEL expression in equivalent amounts as the stress procedure with salt plus hydrogen peroxide. Both stress conditions induced a GroEL expression of twofold relative to control treatment (Fig. 3). The dye or light applied alone did not induce significant GroEL expression, as compared with control experiments. As densitometry control, the quantification of bands indicated by a, b, and c in Fig. 2A (116, 97.4, and 35 kDa, respectively) showed the same intensity in all conditions tested, confirming that the same amount of protein was applied to the gel.
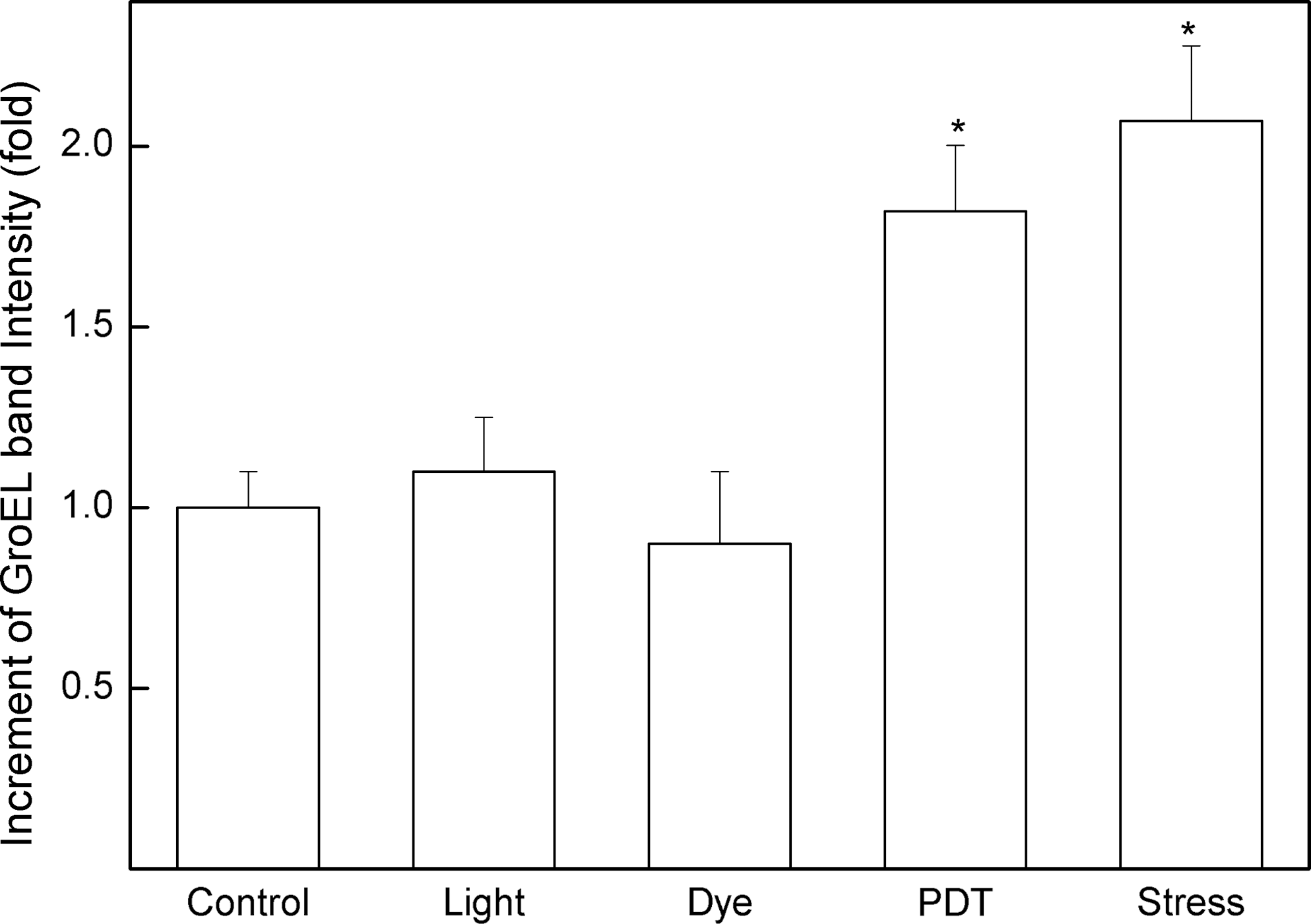
Densitometry of the relative amount of the GroEL band in different protein extracts determined from Western blots of Fig. 2B. Bars indicated by Control, Light, and Dye correspond, respectively, to the bacteria without treatment, light, and RB treatment, separately. (Bars: Control, Light, Dye, PDT, and Stress correspond to bands 1, 2, 3, 4, and 5 of Fig. 2B, respectively). Each bar corresponds to an average of five independent experiments. *p < 0.05.
Nucleic acid electrophoresis after PDT treatment of S. mutans revealed no DNA damage induced by this procedure (Fig. 4), having DNA integrity similar to that of control groups.

DNA extract of Streptococcus mutans: (1) control; (2) submitted to PDT; (3) incubated with 0.1 μmol/L of rose bengal; and (4) photoirradiated with light in the absence of rose bengal. This gel is representative of five independent experiments.
Discussion
The acidogenic capacity, due to formation of acid end-products generated mainly by carbohydrate fermentation, is a characteristic of S. mutans. This acidogenic capacity is a measure of cell's metabolism, and, by extension, of its viability, because no acidification is observed after PDT in a condition that causes 100% cell death. 30,31,38
Figure 1 shows that PDT abolishes the acidogenic capacity of a S. mutans cell suspension, sustaining the effect of this therapy on bacterial metabolism. 25,38 In addition, when the per se toxicity of RB or of the light applied to a non–dye-treated suspension was evaluated, results were similar to those of the controls. That indicates that the decrease of the acidogenic capacity of S. mutans is due to the combination of the dye treatment followed by irradiation (which generates reactive oxygen species) and not to dye or light applied alone. In a previous work, Paulino et al. 30,31 showed that, under the conditions used here, ∼20% of cells were killed. It is a good condition for PDT, because higher concentrations increased cell death, making protein synthesis not feasible, including GroEL protein.
Prinsze et al. 42 describe PDT action in enzymatic systems, causing a decrease in specific activity for some enzymes. Therefore, the acidogenic capacity decrease can be the result of many factors, not mutually exclusive, such as inactivation of enzymes; cell wall/plasma membrane damage; and transport systems among other mechanisms, directly or indirectly involved in the bacterial metabolism and consequent acid extrusion through plasma membrane.
The biochemical and metabolic effects of PDT against microorganisms are diverse. 27 Toluidine blue (TBO) is known to be membrane active on Escherichia coli, because it causes increased permeability, 28 whereas methylene blue causes strand breaks in this organism's nucleic acid. 43 In another gram-negative organism, Proteus mirabilis, methylene blue causes photodamage to the cell envelope, as proved by enhanced cell lysis. 44 Photodynamic microbial damage at molecular level may involve damage to cytoplasmic membrane and also chemical alteration in amino acids, such as crosslinking and methionine sulfoxide formation. Such alterations strongly affect the protein structure and, by extension, its function. 28,43 –45 According to Bevilacqua et al., 25 TBO can diffuse through the bacterial cell membrane, concentrating itself in the interior of bacterial cell, causing cell death after light irradiation, because of reactive oxygen species (ROS) generated by PDT. Whichever the system involved in cellular death induced by PDT in S. mutans, regardless of the existence of a specific target, it is capable of inducing the expression of GroEL, possibly in an attempt to reverse the damage caused by the ROS.
PDT may also cause DNA damage through chemical alteration of nucleotides, mainly guanine, as well as DNA photodamage. 27,45 This was not the case in the present study, because no DNA degradation was observed after PDT. It is noted that methylene blue acts as a photosensitizer to E. coli mainly on DNA, whereas TBO is membrane active and causes no DNA damage in this organism. 46 Associated with that fact, no need of previous incubation exists for cell death induced by PDT, 30,31 which leads us to propose that PDT with RB acts mainly on the cell surface (plasma membrane/cytosol) and not in DNA, as is the case with methylene blue and TBO. 42
Studies on growth-temperature changes led to the discovery of HSP, also called stress proteins. 35 The upregulation of stress proteins is well characterized in S. mutans, 37 like the increase in DnaK levels during thermal stress. 47 With acid stress, an upregulation for isoforms of GroEL chaperone was observed. 48 Our results show, for the first time, an increase in GroEL expression after PDT of S. mutans cells, indicating that PDT induces cell stress.
Conclusion
Data presented show that PDT induces a typical situation of cell stress and no observable DNA damage when applied to a S. mutans suspension. The chaperone system induction is not, however, enough to avoid cell death, because PDT may involve many action sites.
Footnotes
Acknowledgments
We thank Priscila Cerviglieri for linguistic advice. We also thank FAPESP, CNPq, and UNIUBE (PAPE-FAPEMIG 2007/011 and PAPE 2006/052) for the financial support given to our laboratory. MB and TPP received an M.S. and a Ph.D. from CAPES scholarships, respectively.
Author Disclosure Statement
No conflicting financial interests exist.