
Abstract
Introduction
According to Shi et al., 6 lipopolysaccharide (LPS) of Gram-negative bacteria causes a time-dependent increase of MIP-2 mRNA and protein expression in rat alveolar macrophages. Another study presented by the same research group provided direct evidence that oxidative stress generated by H2O2 can induce mRNA expression of a chemokine, MIP-1α, in a rat alveolar macrophage. 7 Moreover, some authors have demonstrated that the regulation of MIP-2 gene expression by oxidative stress in rat alveolar macrophages (AM) can be modulated by transcriptional factors, such as the nuclear factor-κB (NF-κB). 8 –10
NF-κB is a protein that is required for transcription of many pro-inflammatory molecules, which are considered highly important factors in inflammation. Among those molecules, it is relevant to mention the chemokine (IL-8). 11 Shi et al. 6 reported that AM stimulated with LPS produced a significant activation of NF-κB that was attenuated by a pretreatment with the antioxidant N-acetylcysteine (NAC), then suggesting a role for reactive oxygen species (ROS) in the regulation of gene transcription. Therefore, studies focusing on the comprehension of new therapies capable of relieving the symptoms provoked by those cellular events are urgently needed, particularly those that present fewer side effects.
Treatment of ALI includes a variety of pharmacological corticosteroids, methotrexate, and other disease-modifying agents such as cyclosporine and intravenous immunoglobulin, as well as newer treatments like tumor necrosis factor-α (TNF-α) and interleukin-1β (IL-1β) inhibitors. 12,13
Despite those treatments presenting a convenient efficacy, they are associated with several side effects, precluding the employment of those therapies for long periods.
Low-level laser therapy (LLLT) has been used for inflammatory pathologies as an anti-inflammatory therapy, and does not seem to be associated with any side effects. 14 –16 Indeed, LLLT has been employed in the treatment of patients with pleurisy 17 and asthma. 18,19 Furthermore, some authors have demonstrated that the therapy is also beneficial in experimental models of ALI. 20,21 It has been reported in the literature that LLLT can interfere positively in order to relieve the late and early symptoms of airway and lung inflammation by reducing the mRNA expression for inflammatory mediators, such as TNF-α and IL-1β. 22,23 Although it is well established that AM are important cells in the development of ALI, the in vitro effect of LLLT on those cells after stimulation by LPS or oxidative stress has not been completely explored.
Therefore, the objective of this study was to test the hypothesis of a synergistic action involving LLLT and antioxidant agents in modulation of the MIP-2 and ROS mRNA expression in AM, stimulated by oxidative stress and LPS.
Materials and Methods
Cells and culture conditions
The rat AM cell line AMJ2-C11 was acquired from the Rio de Janeiro Cell Bank, Rio de Janeiro Federal University, Clementino Fraga Filho University Hospital. These cells behave similarly to primary AM with respect to the respiratory burst and cytokine release.
24
–26
The cells were cultured in RPMI-1640 supplemented with 5% equine serum, 2 mM
Cell treatment and experimental design
Culture plates (100 mm) were precoated with polyhydroxymethylacrylate to prevent cell adherence. 27 The AM were plated at a concentration of 2.5 × 106 cell mL−1, final volume of 200 μL, in serum-free RPMI-1640 for 4 h prior to the addition of LPS (150 μL) or H2O2 (100 μL), and they were incubated with LPS or H2O2 for a period of 4 h. The NF-κB inhibitor or antioxidant agents were added to culture plates 1 h before LPS or H2O2. For antioxidant treatment, cells were incubated with 20 mM NAC or 1% dimethyl sulphoxide (DMSO). An excessive activation of NF-κB results in an intense inflammatory response of lungs that, consequently, leads to ALI. In order to investigate if the effect of LLLT on MIP-2 mRNA expression involves a cellular signaling through NF-κB, the AM were bathed with BMS 205820 (4 μM), which is an inhibitor of NF-κB, 1 h before LPS or H2O2 exposure. 28 In the same context, the effect of LLLT on NF-κB protein was tested and it was measured in the AM by the enzyme linked immunoassay (ELISA) method 4 h after LPS or H2O2 exposure.
Laser irradiation
A diode laser with an output power of 30 mW and operating at 660 nm (MM Optics, São Carlos, Brazil) irradiated an area of 0.785 cm2 for 252 s, which resulted in an energy dose of 4.5 J/cm2. The optical power was calibrated utilizing a multifunction optical meter (Model 1835C; Newport Corporation, Ivine, CA). The stability of the laser output during the experiment was monitored by collecting light with a partial reflecting surface (4%). For irradiation, the cover plate was removed, and the AM were irradiated for 1 h after the addition of LPS or H2O2.
Involvement of intracellular ROS generation
To prove that the LLLT modulates the intracellular ROS generation, the 2′,7′-dichlorofluorescin diacetate (DCFH-DA; MoBiTec, Göttingen, Germany) was used for ROS detection. DCFH-DA diffuses into the cell and is hydrolyzed by intracellular esterases to polar 2′,7′-dichlorofluorescin. This non-fluorescent fluorescin can be oxidized to the highly fluorescent 2′,7′-dichlorofluorescein by intracellular oxidants. 29 The AM were cultured attached to plates and incubated with 10 μM DCFH-DA for 30 min. Next, the cultures were washed twice with RPMI 1640 and subsequently treated as described before. Fluorescence baseline was measured with a fluorimeter (FLUOstar, BMG LabTechnologies, Offenburg, Germany) immediately after wood dusts were added, as described by Imrich and Kobzik. 29 After 4 h of incubation at 37°C and in 5% CO2 and 21% O2, the fluorescence was measured again. The results are given as the percentage change from baseline values. The addition of H2O2 served as a positive control. NAC is a synthetic antioxidant, which can replenish intracellular glutathione levels. 30 Likely, DMSO has also been widely used as an antioxidant. 31
Measurement of NF-κB protein
For measuring NF-κB protein, the AM were plated at a density of 2.5 × 105 cell mL−1 and allowed to adhere for 2 h. After washing off the non-adherent cells, cultures were incubated in 0.25% FCS RPMI 1640 and treated for 4 h with LPS (10 μg mL−1) or H2O2 (0.5 mM). Cultures were washed again to remove the LPS or H2O2 and incubated in serum-free RPMI 1640 in the absence or presence of NAC (5 ng mL−1) for 1 h. Culture supernatants were aspirated and the cells were lysed by incubation for 20 min with 0.1M HCl (22°C), followed by disruption using a cell scraper. Then, the intracellular content of NF-κB protein was determined using NF-κB ELISA kit by R&D System following the manufacturer's recommendations.
Real-time PCR
PCR of AM (2.5 × 106 cells) was performed on a 7000 Sequence Detection System (Applied Biosystems, CA), using the SYBRGreen core reaction kit (Applied Biosystems). RT-PCR was performed with specific primers for rat MIP-2 195-305 (GenBankTM accession number X66539) forward primer 5 ′-ATCCAGAGCTTGAGTGTGACGC-3′ and reverse primer 5′-AGGCAAACTTTTTGACCGCC-3′; rat Exon (GenBankTM accession number D00475) and rat β-actin-3474-3570 (GenBankTM accession number J00691) forward primer 5′-CATCGCCTTCCGTGTTCCTA-3′ and reverse primer 5′-GCGGCACGTCAGATCCA-3′ as the control. One microliter of RT reaction was used for Real-Time PCR. The PCR primer efficiencies were calculated using standard curves and, the relative expression levels of MIP-2 in real time were analyzed using the 2CT method, presented as the ratio to the expression of the housekeeping gene, actin.
Statistical analysis
Results are expressed as mean ± SEM values. Statistical differences were evaluated by analysis of variance (ANOVA) and the Tukey–Kramer test. The multiple comparisons test was elaborated in order to determine differences between groups, and results were considered significant when p < 0.05.
Results
Effect of LLLT on MIP-2 mRNA expression in AM induced by LPS or H2O2
To test the potential roles of ROS in H2O2 or LPS-induced MIP-2 mRNA expression, the AM were exposed to LPS or H2O2 in the presence of NAC or DMSO or laser (Fig. 1). Four hours after LPS, the AM presented an increase in MIP-2 mRNA expression in comparison with the control cells. Both NAC and DMSO significantly decreased this induction. When the AM were stimulated with LPS and irradiated with laser, the MIP-2 mRNA expression was diminished but less than the treatment with antioxidants (Fig. 1A). To assess the role of oxidants on MIP-2 mRNA expression, the AM were exposed to H2O2 for a period of 4 h, and MIP-2 mRNA was measured by RT-PCR. Levels of MIP-2 mRNA were rapidly induced as early as 1 h following exposure to 0.5 mM H2O2, presenting a maximum after 4 h when compared to the control group. The induction of MIP-2 mRNA expression on AM following 0.5 mM H2O2 exposure was partially attenuated by pretreatment with NAC, as well as DMSO, in comparison to cells exposed to H2O2. The treatment with laser was also efficient in reducing MIP-2 mRNA expression when compared to non-irradiated AM exposed to H2O2 (Fig. 1B).
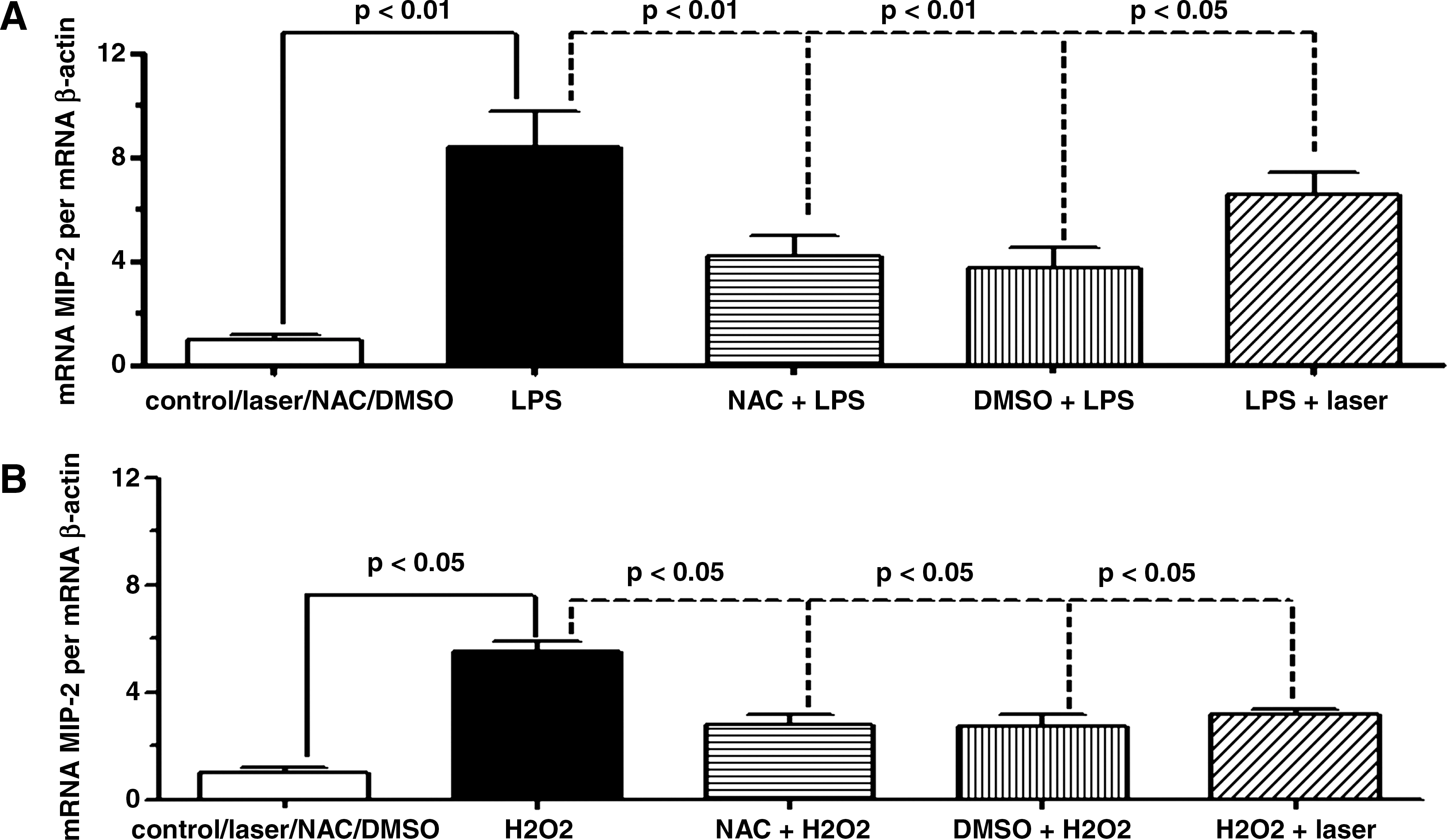
Effect of LLLT on MIP-2 mRNA expression in AM induced by LPS or H2O2. The AM were irradiated 1 h after addition of LPS (
LLLT acts synergistically with NAC on MIP-2 mRNA expression from AM in response to LPS or H2O2
The AM were pretreated with 20 mM NAC 1 h before the addition of LPS or H2O2 (Fig. 2). One hour after LPS addition, the expression of MIP-2 was increased when compared to the control group. When cells were pretreated with NAC, the effect of LLLT was significantly enhanced in comparison to the AM that were irradiated only (Fig. 2A). AM exposure to H2O2 also increased MIP-2 expression when compared to the control group. The effect of the laser on MIP-2 mRNA expression when associated to NAC was also potentiated in relation to the AM that were only irradiated (Fig. 2B).
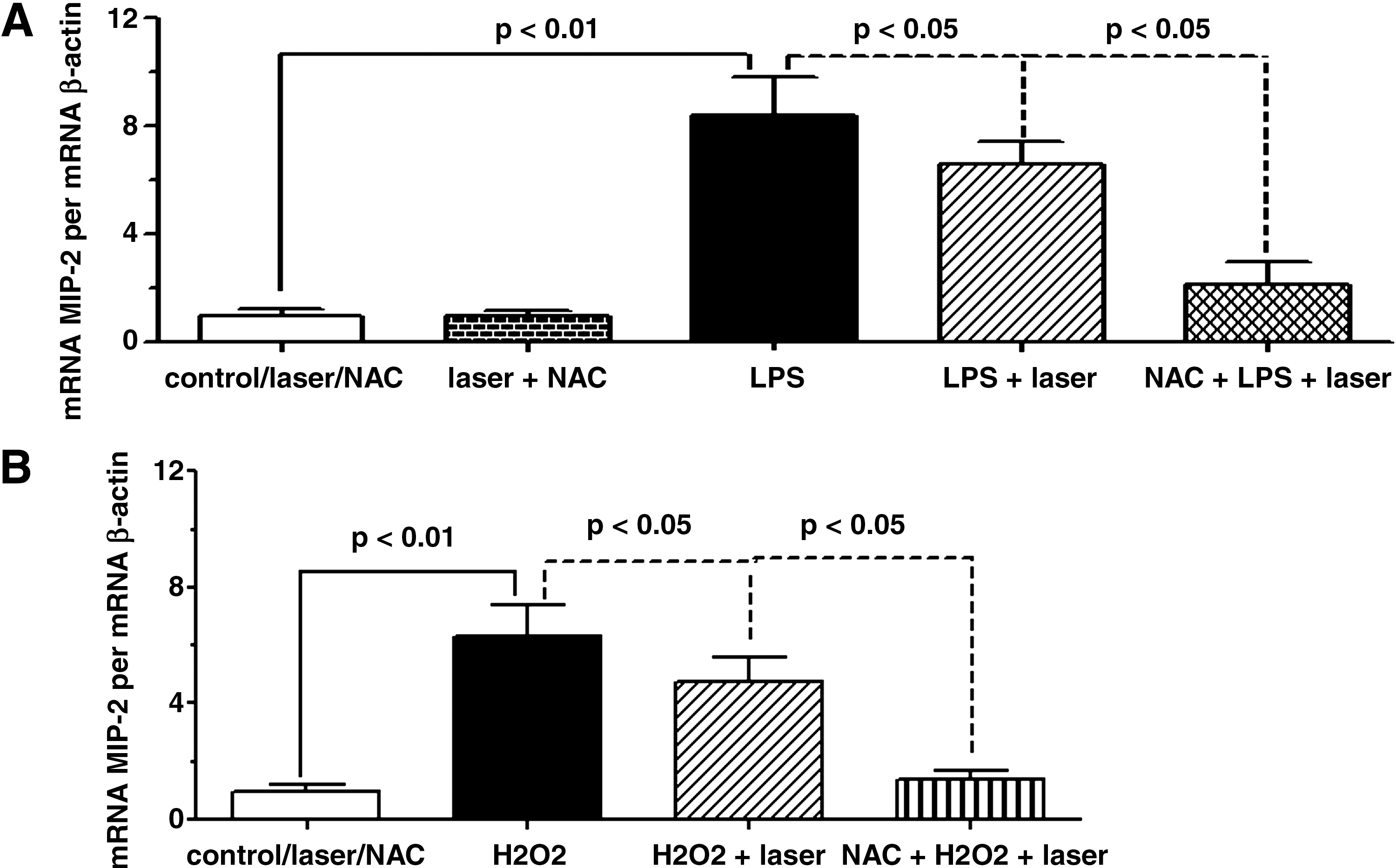
NAC potentiates the effect of LLLT on MIP-2 mRNA expression from AM in response to LPS or H2O2. One hour before addition of LPS (
Effect of NF-κB inhibitor, BMS 205820, in response to NAC or LLLT on MIP-2 mRNA expression induced by LPS or H2O2 in AM
In order to investigate if the laser could mediate the MIP-2 mRNA expression in AM through NF-κB signaling (Fig. 3), the AM were treated with BMS205820 (4 μM) 1 h before the addition of LPS or H2O2. The LPS or H2O2 effects on MIP-2 mRNA expression were not change by pretreatment with BMS 205820. The pretreatment with NF-κB inhibitor reduced the effect of NAC (Fig. 3A) in AM stimulated by LPS or H2O2. Otherwise, the laser (Fig. 3B) effect was suppressed after NF-κB inhibitor in both the LPS or H2O2 condition.
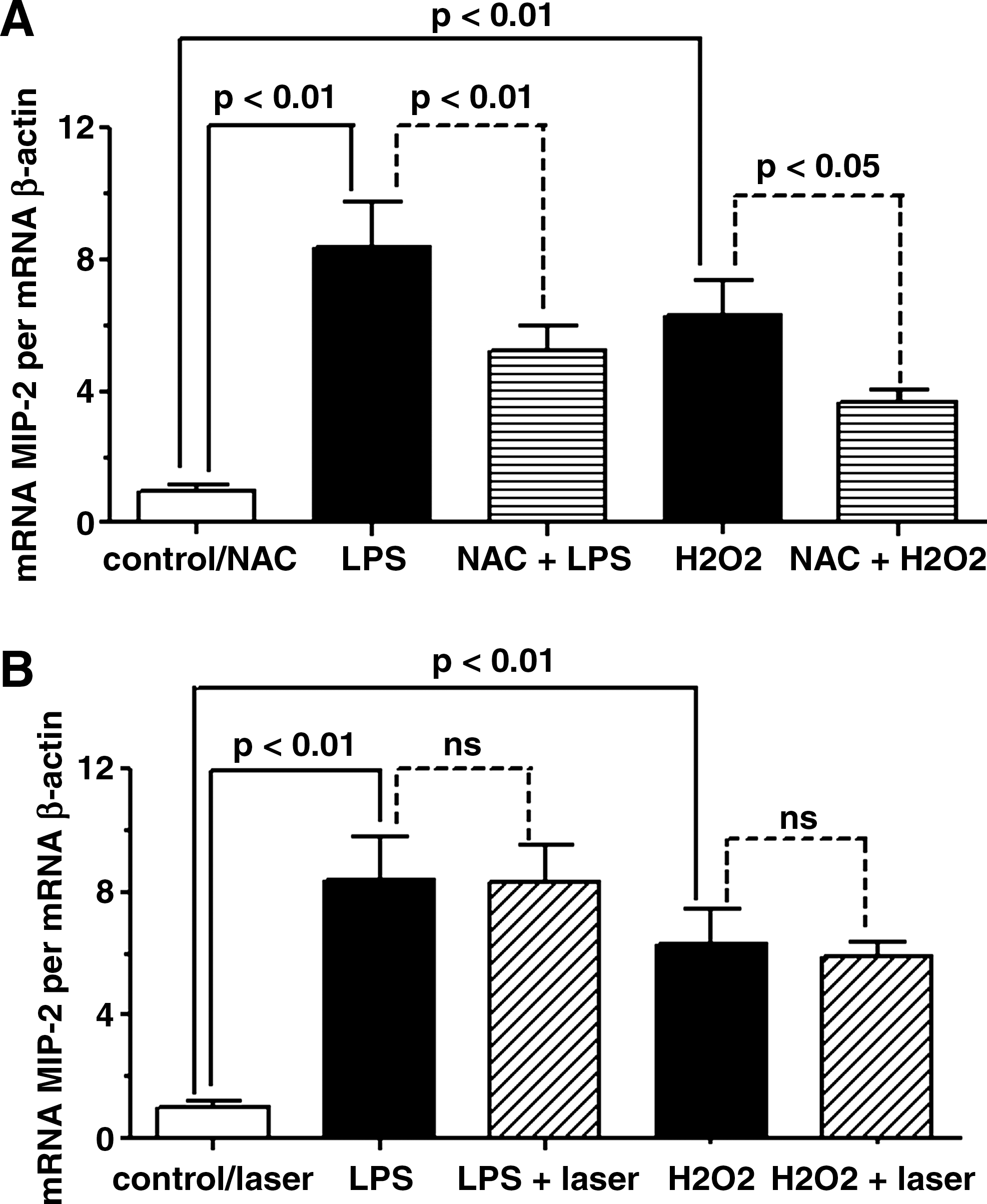
Effect of LLLT or NAC on MIP-2 mRNA expression induced by LPS or H2O2 in AM pretreated with BMS 205820. One hour before addition of LPS or H2O2, the AM were treated with NF-κB inhibitor, and then the effect of NAC (
Effect of NF-κB inhibitor, BMS 205820, on synergistic effect between the LLLT and NAC on MIP-2 mRNA expression induced by LPS or H2O2 in AM
Figure 4 illustrates that BMS 205820 inhibited the synergistic effect of the laser associated with NAC on MIP-2 mRNA expression from AM stimulated by LPS (Fig. 4A) or H2O2 (Fig. 4B). The exposition of AM to BMS 205820 reduces the MIP-2 mRNA expression back to values obtained with NAC without association with the laser.
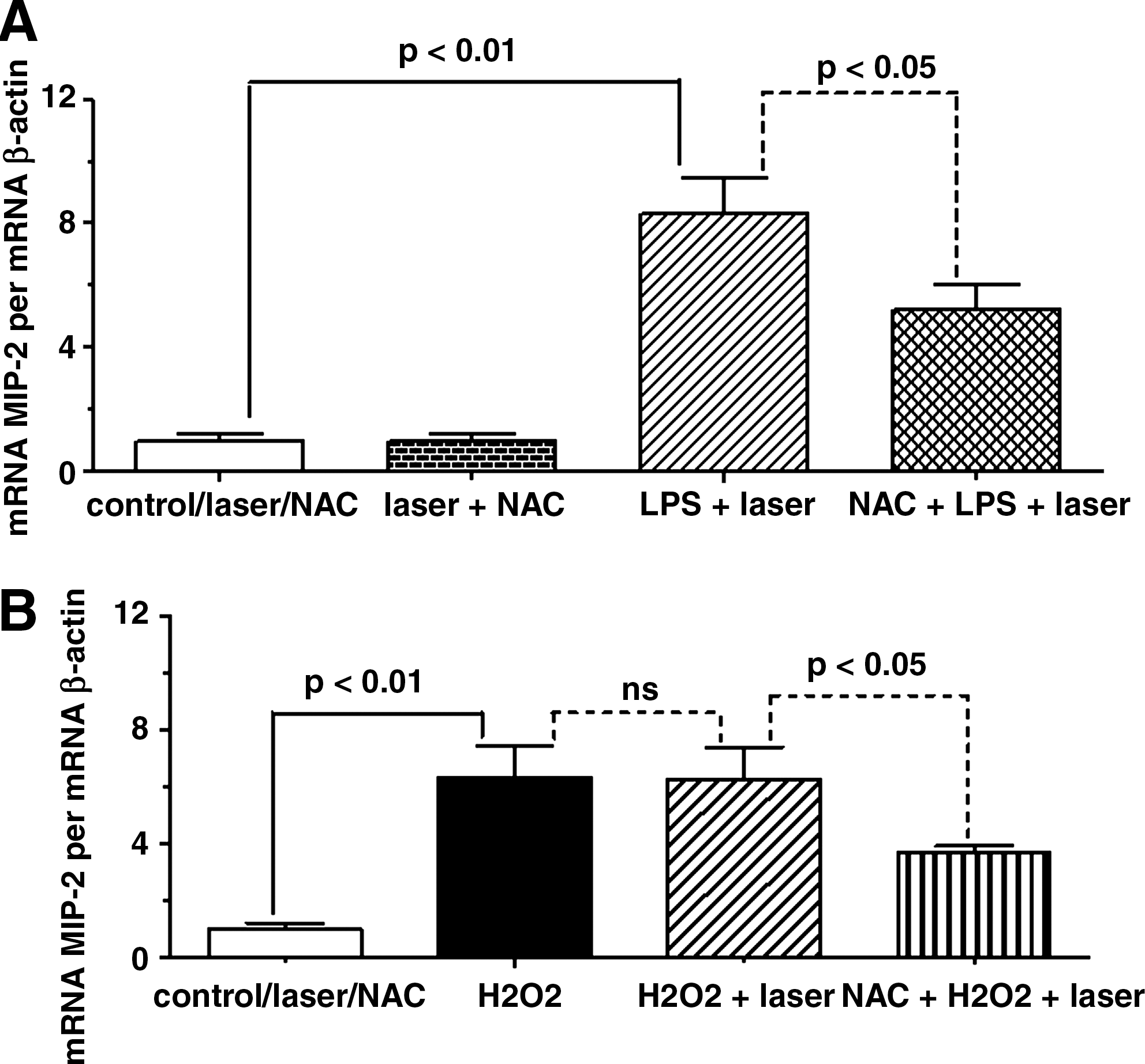
Effect of NF-κB inhibitor, BMS 205820 on the synergistic effect between LLLT and NAC on MIP-2 mRNA expression induced by LPS or H2O2 in AM. One hour before addition of LPS (
Effect of LLLT and NAC and NF-κB inhibitor on ROS intracellular generation in AM in response to LPS
Figure 5 represents the effects of LLLT or/and NAC and NF-κB on ROS intracellular generation in AM in response to LPS. This figure shows an increase in ROS intracellular generation in AM in response to LPS when compared to the control. As can be seen in Fig. 5A, LLLT or NAC strongly reduced the ROS generation in AM stimulated with LPS in comparison to those AM stimulated with LPS but not irradiated or treated with NAC. Figure 5B shows a significant synergistic effect between LLLT and NAC, reducing the ROS generation in comparison to each therapy separately (see Fig. 5A). Figure 5C shows that the effect of LLLT was suppressed when AM were pretreated with BMS 205820, whereas pretreatment alone produced a reduction on the NAC effect in MIP-2 expression. Pretreatment with BMS 205820 reduced the synergistic effect, as can be observed in Fig. 5D.
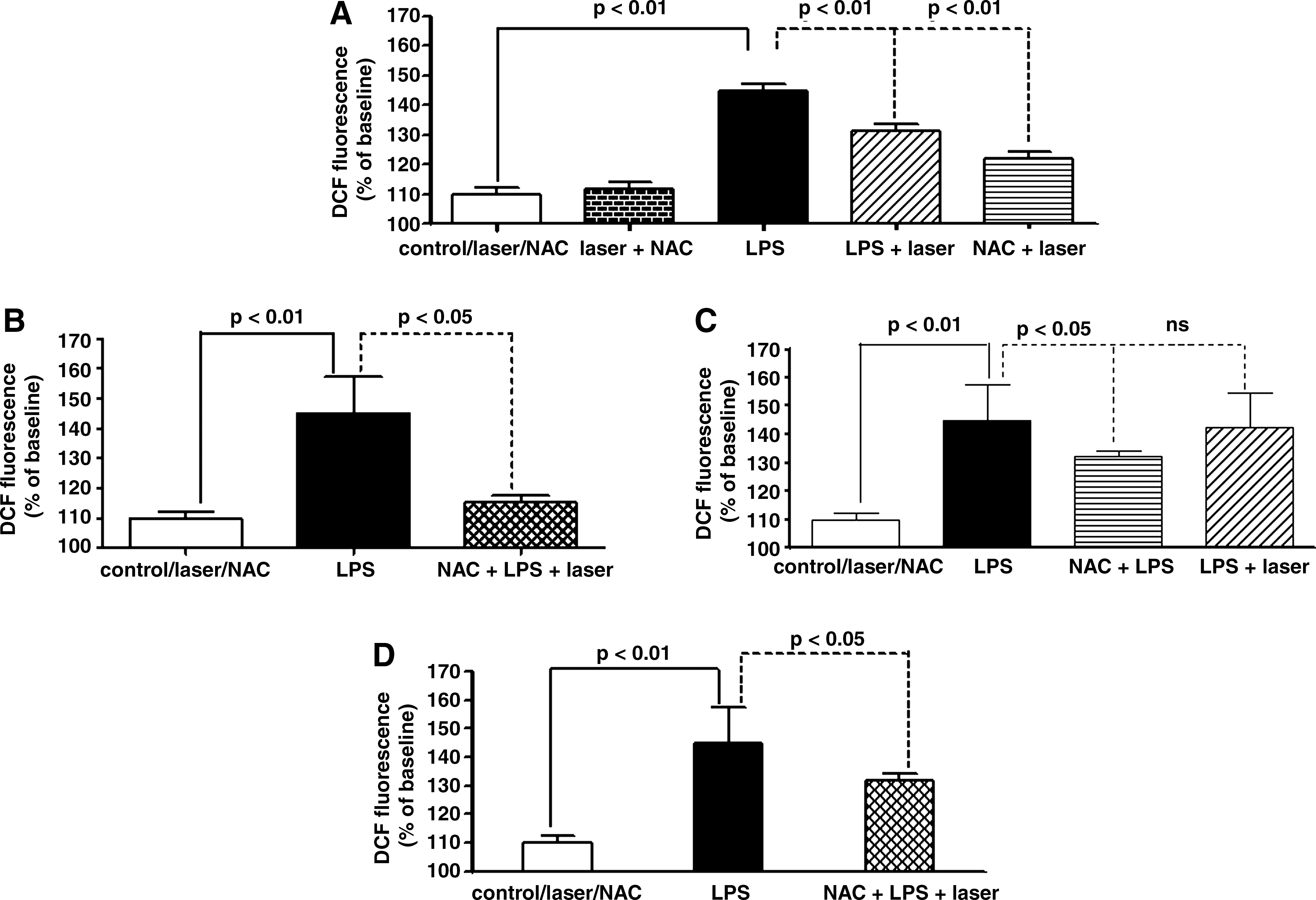
Effect of LLLT, NAC, and NF-κB inhibitor on ROS intracellular generation in AM in response to LPS. (
Effect of LLLT or NAC or NAC associated to LLLT on NF-κB protein induced by LPS or H2O2 in AM
The effect of LLLT or NAC on NF-κB protein after incubation of AM with LPS or H2O2 is shown in Fig. 6. As expected, the exposure of AM to LPS increased the NF-κB protein in comparison to the AM that were not bathed with LPS (see Fig. 6A). The same response, although not so pronounced, was obtained when the AM were exposed to H2O2 instead of LPS. The treatment with NAC strongly reduced the NF-κB protein when compared to the AM bathed with LPS or H2O2 but not treated with the antioxidant. Similar to the NAC case, laser irradiation (Fig. 6B) was also efficient in the reduction of NF-κB protein when the AM were stimulated with LPS or H2O2 in comparison to the AM exposed to LPS or H2O2 but not irradiated. Comparing Fig. 6C to Fig. 6B, it was found that the synergistic effect of NAC associated with LLLT also exists in the NF-κB protein when the AM were stimulated by LPS or H2O2, since there was a stronger reduction of this protein when the two therapies were combined.
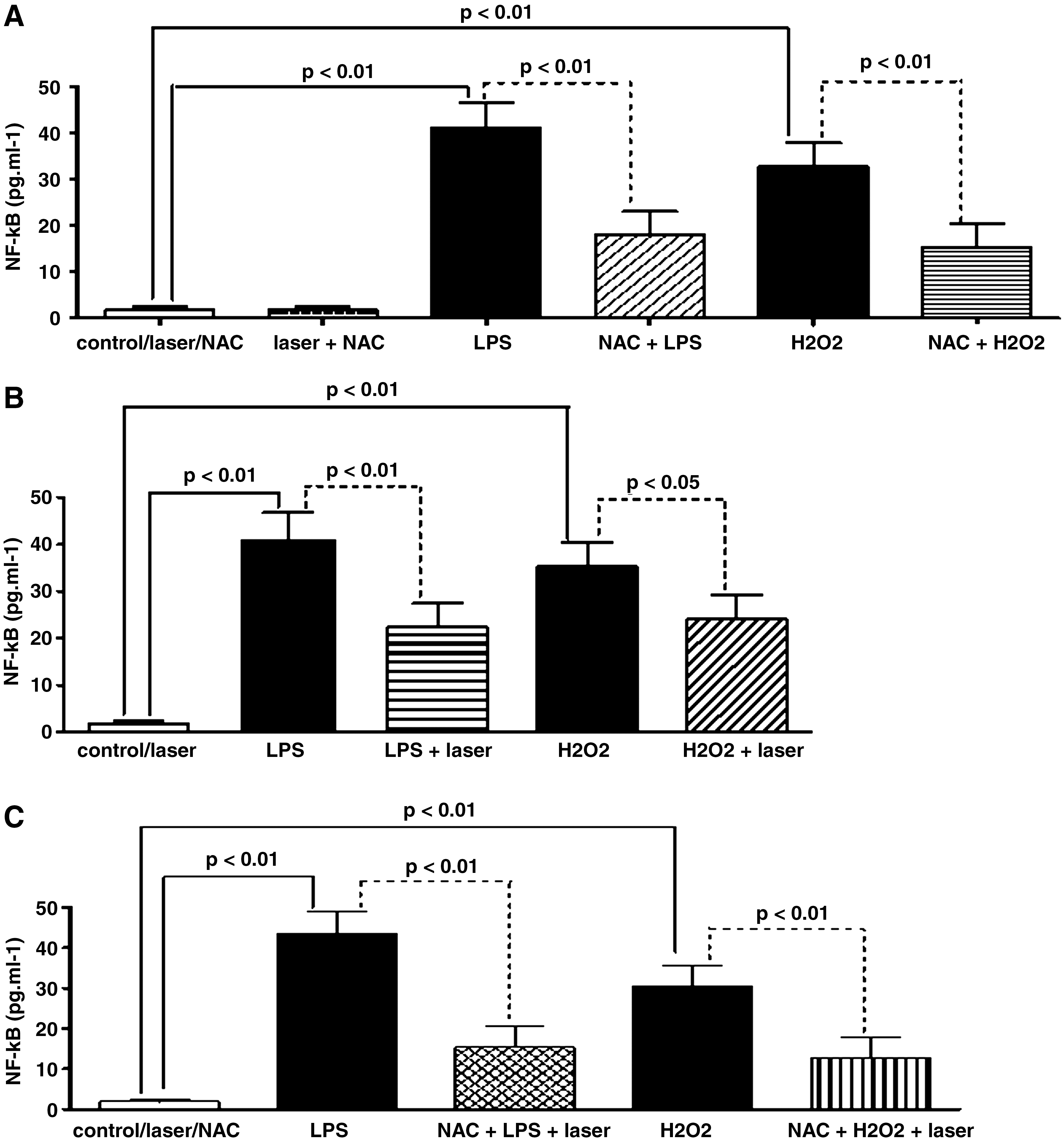
Effect of LLLT or NAC, or NAC associated to LLLT on NF-κB protein after incubation of AM with LPS or H2O2. (
Discussion
The present study investigated if the regulation of MIP-2 mRNA expression in rat AM by the antioxidant agent NAC can be enhanced by LLLT through a mechanism in which the NF-κB signaling would be important. The question addressed herein was based on studies conducted by Shi et al. 6 and Zhang et al. 32 The first reported that the expression of MIP-2 in rat AM is regulated by oxidative stress induced by LPS, whereas the second demonstrated that LLLT induces apoptosis and tyrosine kinase activation by a cellular mechanism mediated by ROS generation.
Medical applications of LLLT are now widespread; nevertheless, the underlying mechanisms involved are not fully understood. 33 ROS are considered the key secondary messengers produced by LLLT. On the contrary, Fujimaki et al. 34 showed evidence that the laser attenuates the production of ROS derived from human neutrophils. Wu and Cheng-Yi 35 demonstrated immediate generation of mitochondrial ROS following laser irradiation, reaching its maximum level 60 min after irradiation. Actually, the effects of low-level laser on ROS are still very controversial.
It was based on the hypothesis that LLLT acts as an antioxidant that the AM were exposed directly to H2O2. It is important to emphasize that H2O2 is commonly used in in vitro systems to generate oxidative stress. When exposed to H2O2 and irradiated with LLLT 1 h later, the AM presented a reduction in the MIP-2 mRNA expression similar to that obtained with DMSO. Interestingly, when the AM were pretreated with NAC 1 h before H2O2, the effect of LLLT on MIP-2 mRNA expression was significantly potentiated. It was observed that in the presence of H2O2, the synergism of LLLT on the effect of NAC on MIP-2 expression was more pronounced. The same effect of LLLT was found when the AM were exposed to LPS. Those findings let suppose that LLLT should enhance the effect of NAC on the MIP-2 expression by the reduction of intracellular ROS generation.
It is for that particular reason that the present study investigated the effect of LLLT on intracellular ROS generation in AM activated by LPS. Those assays evidenced the antioxidant effect of LLLT, since the intracellular ROS generation was markedly reduced when the AM were irradiated. In our experiment, the AM bathed with H2O2 served as a positive control of ROS generation. Experiments on the intracellular ROS generation were done only with LPS as the stimulus of AM, since we were focusing on the effect of LLLT from the point view of the disease. In the present study, the investigation of the effect of LLLT on ROS generation is important because the AM release diverse inflammatory mediators when stimulated with LPS; therefore, the LLLT may reduce the MIP-2 mRNA expression by indirect mechanism without involvement of ROS. However, the confirmation that LLLT acts directly on intracellular ROS generation strongly suggests that the laser would be an antioxidant agent in ALI induced by LPS. Interestingly, when the AM were pretreated with NAC and irradiated, the reduction of intracellular ROS generation was potentiated. Those results seem to corroborate that LLLT increases the NAC effect on MIP-2 mRNA expression, at least partially, due to the reduction of intracellular ROS generation. In an experimental model of rat skeletal muscle, Rizzi et al. 36 showed that LLLT reduced the inflammatory response provoked by trauma, and was able to block the effects of ROS release and the activation of NF-κB.
Regarding the effect of LLLT on mRNA expression of inflammatory mediators, Albertini et al. 37 in an acute inflammation model induced by carrageenan in rat paw demonstrated that laser therapy reduced the COX-2 mRNA expression. Moreover, in the same experimental model, the anti-inflammatory effect of LLLT diminished the mRNA expression to TNF-α, IL-1β, and IL-6. 38 In lung inflammation models, we reported that the effect of LLLT on mRNA expression attenuated the neutrophils infiltration into lung and the pulmonary microvascular leakage by a remarkable reduction of IL-1β mRNA expression in lung of rats systemically inflamed with LPS. 23 Furthermore, we observed that LLLT reduces the anti-apoptotic factors mRNA expression of neutrophils from mice inflamed systemically with LPS through a mechanism dependent on the NF-κB signaling. 39
It is well established that in cellular signaling via NF-κB, this protein is required for the transcription of many pro-inflammatory molecules, including cytokines such as IL-1β, TNF-α, IL-6, and IL-8. 35,40 –42 An excessive activation of NF-κB results in ALI. ROS have been implicated in the activation of the transcription factor NF-κB. 43,44 Thus, based on the hypothesis of Shi et al. 6 that considers ROS as a common signal to regulate the gene expression of most chemokines through the factor transcription NF-κB, we investigated if the ability of LLLT in reducing the MIP-2 mRNA expression and intracellular ROS production could be mediated by a cellular signaling involving the NF-κB. Our results demonstrated that the pretreatment with BMS205820, an inhibitor of NF-κB, completely inhibited the effect of the laser, as well as partially reducing the NAC effect on MIP-2 mRNA expression of AM activated by LPS or H2O2. These findings suggest that the effect of LLLT on MIP-2 mRNA expression can be limited by NF-κB inhibitor agents. The synergism between LLLT and NAC on MIP-2 expression was not blocked by the NF-κB but only reduced to NAC values. In this context, the increased intracellular ROS generation after LPS was not changed by LLLT when the AM were pretreated with BMS 205820. This finding seems to suggest that the role of NF-κB is very important in the effect of LLLT, since pretreatment with BMS 205820 reduced the synergistic effect between LLLT and NAC.
Finally, the effect of LLLT on NF-κB protein reinforces the idea that LLLT can act on MIP-2 mRNA expression through this transcription factor. It confirms the hypothesis that the signaling pathway involving NF-κB is fundamental for LLLT efficacy in the experimental model presented herein.
Conclusion
Results obtained in the present study strongly suggest that LLLT can potentiate the NAC effect on MIP-2 expression, modulating the ROS generation through a cellular mechanism that seems to be significantly influenced by NF-κB signaling.
Footnotes
Author Disclosure Statement
No competing financial interests exist.