
Abstract
Introduction
Photodynamic therapy (PDT) is a very powerful modality of cancer treatment with an interesting history. The activation of the photochemical properties of substances called photosensitizers results in a cytotoxic and vasculotoxic chain of events for their photoproducts. 3
Photodynamic therapy (PDT) is presently a widely accepted modality for the treatment of a variety of solid tumors. 4,5 The therapeutic action occurs during the absorption of light by the photosensitizer, which leads to the subsequent conversion of chemical reaction products that act upon the cells to induce necrosis and apoptosis. 6 The effects of photodynamic therapy depend on the simultaneous presence of the drug, excitation light, and oxygen. Oxygen is essential for promoting the production of highly reactive singlet oxygen species, which destroy cellular substrates and lead to cell death. 7
Porphyrins and their analogues are frequently adopted as photosensitizers. Their tetrapyrrolic macrocycle exhibits absorption in the red region (600–800 nm) of the visible spectrum, which is endowed with an especially high penetration power into human tissue and is not absorbed by the endogenous cell constituents, thereby minimizing the risk of general photodamaging effects. Photofrin® (porfimer sodium) is a complex of porphyrin oligomers that has been approved for specific clinical applications in several countries in Europe, America, and Asia and has been under investigation for malignant and nonmalignant diseases. 8,9 The corresponding photosensitizer used in Brazil is called Photogem® and is produced in Russia. It is described chemically, photophysically, and by diagnostic and therapeutic features as being identical to Photofrin. 10
PDT using Photogem, a hematoporphyrin derivative-type photosensitizer, has been used clinically in the field of gynecology for the treatment of dysplasia and carcinoma in situ of the uterine cervix; 11,12 the use of this therapy aims to estimate the photosensitizer distribution, determining the tumor boundaries before photodynamic treatment. 13
Several studies from in vitro culture systems and in vivo animal models have been proposed to explain the three morphologically distinguished forms of programmed cell death (autophagic cell death, necrosis, and apoptosis), but little is known about the kinetics of the intracellular process after PDT. 14,15 The molecular nature of the photo-oxidized targets has a profound influence on the signaling pathways and mode of cell death initiated following PDT. In general, photoactive compounds localized in the mitochondria promote apoptosis, within a certain threshold of oxidative stress, while PDT with photosensitizers targeting either the plasma membrane or lysosomes can either delay or block the apoptotic program, predisposing the cells to necrosis. 11
The purpose of this study was to investigate the effects of PDT using Photogem in five different glioma cell lines (U87, U138, U252, U343, and T98G) and the role of apoptosis during this process.
Materials and methods
Chemicals
Photogem® (PG) was purchased from the Moscow Institute of High Chemical Technology (Moscow, Russia) as a freeze-dried porfimer. It was stored at stock solution in phosphate-buffered saline (PBS, Invitrogen, Carlsbad, CA) with a solution concentration of 5 mg/ml and was kept at −20°C until use. Storage, dilution, and incubation were performed under experimental conditions that prevented the exposure of Photogem® to light.
All chemicals and additives for the cell culture were purchased from Gibco Invitrogen (products for culture, Karlsruhe, Germany) unless otherwise specified.
Cell culture
U87 (glioblastoma cell line with a p14ARF/p16 deletion and a PTEN mutation), U138 and U251 (glioblastoma cell lines with TP53 and PTEN mutations and a p14ARF/p16 deletion), U343 (anaplastic astrocytoma cell line with a p14ARF/p16 deletion and a PTEN mutation), and T98G (glioblastoma cell line with TP53 and PTEN mutations and a p14ARF/p16 deletion) cell lines were originally obtained from the American Type Culture Collection (ATCC, Manassas, VA). Cells were cultured and maintained at 37°C with 5% carbon dioxide (CO2) in Dulbecco's modified Eagle medium (DMEM) containing 10% (v/v) fetal bovine serum (FBS), 100 IU/mL penicillin, 100 μg/mL streptomycin/neomycin, and 1% nonessential amino acid (Invitrogen).
Treatment of cells
The experiments were carried out in 25-cm3 flasks (TPP- Techno Plastic Products AG; Transadingen, Switzerland). Different groups of cells (U87, U138, U251, U343, and T98G) were seeded at a density of 1 × 105 cells per flask. After 3 h the medium was removed, and 5 ml of a Photogem-DMEM solution at a final concentration of 5 μg/mL was added to each flask. Cell incubation with fresh culture medium without Photogem® served as the control. After the incubation time, the photosensitizer- containing medium was removed and the cells were irradiated with LED (light emitting diodes) devices (Lat – IFSC: Department of Physics and Science of Materials, Institute of Physics of São Carlos/University of São Paulo – Brazil) at 630 nm. The irradiation was performed at a fluence of 25 mW/cm2 with a light dose of 25 J/cm2 for 17 min. After PDT, the cells were returned to the incubator and kept at 37°C with 5% carbon dioxide, and after 24 h the cells were analyzed.
Cell viability assay
Cell viability was assessed using a trypan blue dye-exclusion method. Briefly, the cells were trypsinized and then resuspended in 1,000 μL of DMEM, and then trypan blue solution (0.4%) was added to the cells in an equal volume. Cells were then counted using a hemacytometer after 15–20 min of incubation at room temperature, and the ratio of viable cells to the total number of cells was calculated and recorded.
RNA isolation and real-time polymerase chain reaction
Total RNA was extracted with Trizol reagent (Applied Biosystems, Carlsbad, CA) according to the manufacturer's instructions. In preparation for the real-time polymerase chain reaction (PCR), reverse transcription of RNA samples was performed using the High-Capacity cDNA kit (Applied Biosystems). Real-time PCR analysis of cDNA was performed at 95–60°C for 45 cycles in the sequence detection system of an ABI Prism 7300 (Applied Biosystems), in accordance with the manufacturer's instructions, using Taq man reaction master mix (Applied Biosystems) and the primer for caspase-3. In each RNA sample, β-actin mRNA was measured as the housekeeping gene.
Statistical analysis
Primary data are presented with standard deviations. Differences between means were evaluated by one-way ANOVA and were followed by Bonferroni correction when there were multiple outcome measures (three or more groups), although there was possible concern that the results might be perceived as being a fishing expedition. p Values smaller than 0.05 (p < 0.05) were considered to be significant. All analyses were performed using the Graph Pad Prism Software, version 4.0 (Graph Pad Software, Inc., San Diego, CA).
Results
Cell viability assay
The proportion of viable cells in the U87 and U138 cell lines was reduced when the cells were treated with PDT (Photogem®) (p < 0.05). The T98G, U251, and U343 cell lines showed no significant differences between the controls and the PDT-with-Photogem groups (Fig. 1).
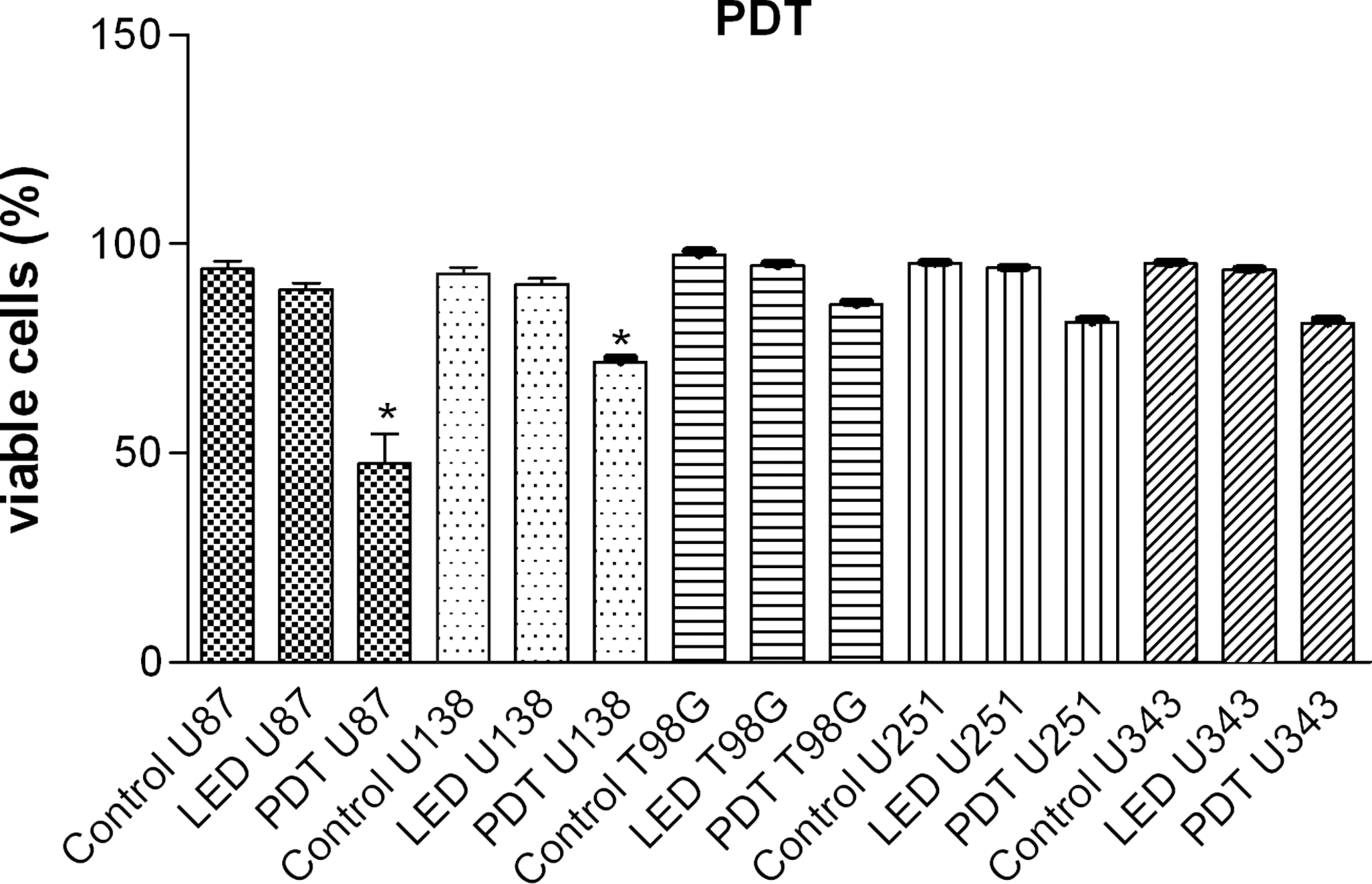
Photogem PDT-induced cytotoxicity. Glioma cell lines (U87, U138, T98G, U251, and U343) were pretreated with 5 μg/ml of Photogem for 4 h and then exposed to LED. Cytotoxicity was determined by the trypan blue dye-exclusion method at 24 h after Photogem-PDT (*p < 0.05, Bonferroni Multiple Comparison Test). The T98G, U251, and U343 cell lines showed no significant differences between the controls and the PDT-with-Photogem groups.
Caspase-3 mRNA levels in glioblastoma cell lines after PDT
The expression of caspase-3 was higher in the U87 and U138 cells irradiated with PDT (Photogem®) (p < 0.05) (Fig. 2), compared with unirradiated cells. No significant differences were found in the T98G, U251, and U343 cell lines.
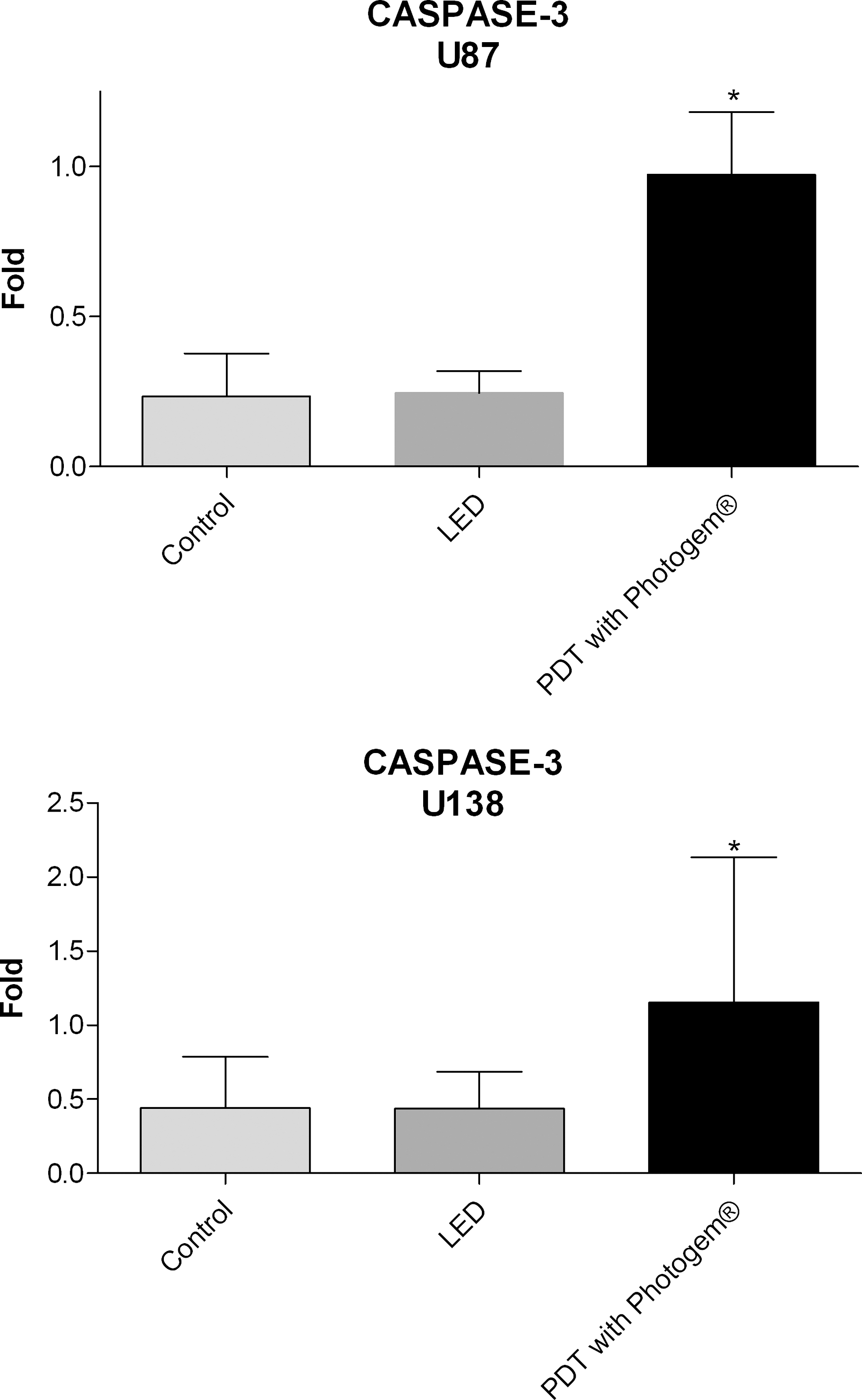
Representation of the average values (±standard deviation) of gene expression for CASPASE 3 in the U87 and U138 cells treated with Photogem-PDT when compared with the LED-only and control cell groups (*p < 0.05, Bonferroni Multiple Comparison Test).
Discussion
Photodynamic therapy is a promising technique for the treatment of many tumors, and it is currently considered to be an efficient and safe adjuvant therapy. In this study, the phototoxic effects of Photogem, a photosensitizing drug, were evaluated in five different glioma cell lines (U87, U138, U252, U343, and T98G); in addition, we evaluated whether PDT using Photogem exerts its activity by inducing apoptosis.
Previous studies have shown that many glioma cell lines are partially or completely resistant to PDT. 16,17 In an attempt to identify a sensitizing agent capable of activating cell-death mechanisms in some glioma cell lines, we investigated the efficacy of PDT with Photogem®. 18 When tested, Photogem had a very limited effect in the T98G, U251, and U343 glioma cell lines, but produced a significant decrease in the viability of U87 and U138 glioma cells. The differential responses in cell lines derived from the same tumor (i.e., glioblastoma) can be explained by the heterogeneity of genetic pathways involved in glioblastoma formation, resulting in different sensitivities of tumors to diverse treatment modalities.
The complexity of cell death induced by PDT has been demonstrated in several cell types, and it may be influenced by variability in the intracellular accumulation of the photosensitizer. Recent studies have shown that PDT may induce nonapoptotic cell death associated with the induction of autophagy. 16 Necrosis is the major cell-death morphology induced by PDT with compounds localized to the plasma membrane. 18 Although a biochemical pathway mediating necrosis following PDT has not yet been identified, certain factors (such as Ca2+ overload and the origin and type of generated reactive oxygen species) may be decisive in promoting a necrotic cell-death pathway. Evidence indicates that PDT can evoke the main cell-death morphologies that have been described, including apoptotic, necrotic, and autophagic cell death. This explains why in some systems the specific inhibition of one death signal is not sufficient to block PDT-mediated cell death. Recent studies point out that while apoptosis is probably the preferred path to cell death, it is not the only one. 11
Karmakar et al. 19 used 5-aminolevulinic acid (5-ALA) as a photosensitizer for PDT to induce apoptosis in human malignant glioblastoma U87MG cells. Wright staining and the ApopTag assay showed the occurrence of apoptosis morphologically and biochemically, respectively. The 5-ALA-PDT caused an increase in the Bax:Bcl-2 ratio and the mitochondrial release of cytochrome c and apoptosis-inducing factor (AIF), with the activation of calpain, caspase-9, and caspase-3 occurring in the course of apoptosis. These results suggested that 5-ALA-PDT induces apoptosis in U87MG cells by the suppression of survival signals and the activation of proteolytic pathways.
Caspase-3 has been identified as a key mediator of apoptosis. Once activated, it is responsible for the proteolytic cleavage of a broad spectrum of cellular targets, thereby leading to cell death. 20 The degradation of PARP (polyADP-ribose polymerase), actin, and SREBP (steroid regulatory element binding protein), which can occur during apoptosis, confirms the activation of caspase-3 and suggests an irreversible phase of the death process by apoptosis in these tumors. 20,21 Ray et al. 21 showed that an increase in caspase-3, together with DNA fragmentation, indicates that apoptosis is a mechanism capable of causing cell death in malignant tumor cells. In our study, we observed that PDT with Photogem resulted in the induction of cell death in U138 and U87 glioma cell lines, and that this cell death was mediated by apoptosis, although these data do not exclude the possibility that other cell-death mechanisms could also be involved.
The absence of activation of the apoptotic pathway in wild-type p53 T98G, U251, and U343 cells is puzzling. Several studies have demonstrated that apoptosis induced by DNA-damaging drugs usually requires the function of the tumor suppressor gene p53, which primarily engages the cell-intrinsic apoptosis signaling pathway. The U138 cell line is mut-p53, and this finding is very important because it shows that even in the absence of a functional p53, the synergistic proapoptotic effects of a photosensitizer agent can occur. 22,23 This observed apoptotic activity may be related to a signaling pathway for apoptosis that is independent of p53. Death receptors, which are members of the TNF receptor family, signal apoptosis independently of the p53 tumor-suppressor gene, as observed by Wang and El-Deiry 24 in the TRAIL (Apo2L) receptor, which is capable of initiating apoptosis through the engagement of its death receptors.
In conclusion, we showed that some glioblastoma cell lines are responsive to PDT with Photogem (5 μg/mL) and that the antitumor activity is mediated, at least partially, by apoptosis. Considering the rapid absorption of Photogem and the excellent tissue permeability of the diode laser, PDT with Photogem is expected to exert higher antitumor activity than other photosensitizers. 25 –29 Our in vitro results provide data relevant to the development of new experiments using PDT as a modality of treatment for gliomas, with the aim of better responses and patient outcomes. Understanding the molecular differences and identifying the cross-talk between cell-death mechanisms will be crucial for the development of new therapeutic modalities using PDT aimed at increasing the efficacy of death-induction in cancer cells in which one or several of these pathways might be inactivated.
Footnotes
Acknowledgments
This study was supported by FAEPA-Fundação de Apoio ao Ensino, Pesquisa e Assistència do Hospital das Clinicas da Faculdade de Medicina de Ribeirão Preto da Universidade de São Paulo.
Author Disclosure Statement
No competing financial interests exist.