
Abstract
Introduction
IGF-I is secreted primarily in the liver after stimulation with growth hormone, and it maintains cell growth and development and plays an important role in fracture healing. 7 In addition, IGF-I plays an essential role in the anabolic regulation of bone metabolism. 8,9 IGF-I can regulate Runx2 DNA binding through the sequential activation of the phosphatidylinositol 3 kinase/p21 activated kinase 1 (PI3K/Pak1) and extracellular signal-regulated kinase 1/2 (ERK1/2) signaling cascades. 10 IGF-I receptor inhibitors prevent IGF-I signaling by binding to the IGF-I receptor, specifically blocking the phosphorylation of tyrosine kinase (Y1136) on the β-subunit of the IGF-I receptor. 11
This study determined the effect of LLLT on the expression of IGF-I and Runx2, ERK phosphorylation, and mineralization in mouse osteoblastic MC3T3-E1 cells. Additionally, we examined the effect of interrupting IGF-I and bone morphogenetic protein (BMP) signaling on mineralization in vitro.
Materials and Methods
Cell culture
The mouse osteoblast-like cell line MC3T3-E1 was provided by the RIKEN Bioresource Center (Tsukuba, Japan). The cells were maintained in α-minimum essential medium (α-MEM; Invitrogen, Grand Island, NY), containing 10% (vol/vol) fetal bovine serum (HyClone Laboratories, Logan, UT) and 1% (vol/vol) penicillin–streptomycin solution (50 U/ml penicillin and 50 μg/ml streptomycin; Sigma Chemical, St. Louis, MO), at 37°C, in a humidified atmosphere of 95% air and 5% CO2. The medium was changed twice weekly.
LLLT Procedure
The MC3T3-E1 cells were plated in 100-cm2 cell-culture dishes at a density of 2 × 104 cells/cm2. After a 24-h incubation, the nearly confluent cells were irradiated once according to an experimental model described previously. 4 –6 In brief, a low-level Ga-Al-As diode laser (model Panalas-1000; Matsushita, Osaka, Japan), which has a wavelength of 830 nm (maximum power output, 500 mW), was used. The laser beam was delivered by an optical fiber 0.6 mm in diameter that was defocused at the tip by a concave lens to provide a uniform circle of irradiation, 100 mm in diameter, at the cell-layer level. The power density of the laser beam was measured by using a laser power meter (ORION PD; Ophir, Jerusalem, Israel). We used 2-Hz pulsed irradiation, according to Ueda and Shimizu (2003). 4 The exposure and rest times were the same for each irradiation pulse (50% duty cycle). The total energy corresponding to pulse irradiation (2 Hz) exposures for 5, 10, and 20 min was 0.96, 1.91, and 3.82 J/cm2, respectively. Laser irradiation was performed 1 day after subculture (day 1) on a clean bench. Control dishes were placed on a clean bench for corresponding periods with no irradiation.
Real-time polymerase chain reaction
Total RNA was isolated from cultured MC3T3-E1 cells by using an RNeasy Mini Kit (Qiagen, Valencia, CA). Aliquots containing equal amounts of total RNA were subjected to real-time PCR. First-strand cDNA synthesis was carried out by using 1 μg of DNase-treated total RNA in 20 μl of a solution containing first-strand buffer, 50 ng random primers, 10 mM dNTP mixture, 1 mM DTT, and 0.5 U reverse transcriptase, at 42°C for 40 min. The cDNA mixtures were diluted fivefold in sterile distilled water, and 2-μl aliquots were subjected to real-time PCR by using SYBR Green I dye. The real-time PCR was performed in 25 μl of a solution containing 1 × SYBR Premix Ex Taq (TaKaRa, Tokyo, Japan), and 0.2 μM specific sense and antisense primers, as shown in Table 1.
The primers were designed by using Primer3 software (Whitehead Institute for Biomedical Research, Cambridge, MA). PCR was carried out in a thermal cycler (Smart Cycler II system; Cepheid, Sunnyvale, CA), and the data were analyzed by using Smart Cycler software (v. 2.0). The real-time PCR conditions consisted of 35 cycles of 95°C for 5 s and 60°C for 20 s. The specificity of the PCR products was verified by conducting melting-curve analyses between 60°C and 95°C. All real-time PCR was performed in triplicate, and the data of two different experiments were combined. The levels of mRNA expression were calculated and normalized to the level of GAPDH mRNA at each time point.
Extracting protein from cultured cells
To obtain whole-cell extracts, MC3T3-E1 cells that had been cultured with or without laser irradiation were rinsed with phosphate-buffered saline (PBS), and then exposed to a lysis buffer, consisting of 50 mM Tris-HCl, 0.1% Triton X-100, 0.1 mM EDTA, and 1 mM phenylmethylsulfonyl fluoride (PMSF). Cells in the lysis buffer were sonicated 3 times for 10 s each time. Aliquots that contained equal amounts of protein were subjected to sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE).
SDS-PAGE and Western blotting
At the indicated times, conditioned medium was collected, and aliquots containing equal amounts of protein were then analyzed with SDS-PAGE. Samples were run on 5% to 20% gradient cross-linked polyacrylamide gels and transferred to polyvinylidene fluoride membranes, by using a semidry transfer unit. After the transfer, the membranes were probed with anti-IGF-I, anti-Runx2, anti-p-ERK, or anti-β-tubulin antibodies as an internal standard (Santa Cruz Biotechnology, Santa Cruz, CA; dilution, 1:500) followed by a biotin-conjugated secondary antibody (dilution, 1:10,000). Then the membranes were treated with horseradish peroxidase–conjugated streptavidin. The immunoreactive proteins were visualized by using a chemiluminescence kit (Amersham Life Science, Buckinghamshire, UK) according to the manufacturer's protocol.
Effects of PPP on LLLT-induced IGF-I and Runx2 expression and ERK phosphorylation
The MC3T3-E1 cells were plated in 100-cm2 cell-culture dishes at a density of 2 × 104 cells/cm2. After incubation for 23 h, exogenous 100 ng/ml cyclolignan picropodophyllin (PPP; Calbiochem, Darmstadt, Germany), an IGF-I–receptor inhibitor, was added and incubated for 60 min, and then LLLT was performed. The cells were incubated for a further 5, 10, 20, 30, or 60 min before measuring the phosphorylation of ERK and for 48 h before measuring the IGF-I and Runx2 expression. The total RNA and whole-cell extracts were isolated as described earlier, and equal amounts of RNA and protein were subjected to real-time PCR and Western blotting, respectively.
Determining the calcium content in mineralized nodules
MC3T3-E1 cells were cultured for 48 h after laser irradiation with or without noggin or PPP. The conditioned medium was filtered through a 0.22-μm membrane (Millipore, Billerica, MA) and then kept at −20°C until required. Aliquots of conditioned medium, which contained equal amounts of protein, were diluted with α-MEM supplemented with 10% FBS and used in this experiment at dilutions of 1:1. The MC3T3-E1 cells were placed in 96-well tissue-culture plates at a density of 2 × 103 cells/cm2 and cultured in diluted conditioned medium containing 10 mM β-glycerol phosphate and 50 μg/ml ascorbic acid for up to 14 days. The culture medium was changed every second or third day. Cell condition and nodule formation were checked routinely with phase-contrast microscopy (Nikon, Tokyo, Japan).
After 14 days, the medium was discarded, 300 μl of 0.5 M HCl was added to each well, and the cells were left overnight to decalcify the mineralized nodules. The calcium content was quantified by using a Calcium C-Test kit (Wako Fine Chemicals, Osaka, Japan), following the kit protocol.
Statistical analysis
All data are presented as mean ± standard deviation (SD). Statistical significance was determined by using the Mann-Whitney U test or Kruskal–Wallis test. P < 0.05 was deemed to indicate statistical significance.
Results
Effect of different doses of LLLT on IGF-I mRNA expression
We examined IGF-I mRNA expression by using real-time PCR as a marker of bone formation. At an irradiation dose of 0.96 J/cm2, the expression of IGF-I mRNA was increased at 24 and 48 h, compared with controls; at a dose of 1.91 J/cm2, it was increased at 9, 12, 24, and 48 h; whereas at 3.82 J/cm2, it was increased at 12, 24, and 48 h, compared with controls. In particular, with 1.91 J/cm2 (irradiation for 10 min), the IGF-I expression was highest in all dose groups, and it increased continuously, for up to 48 h of cell culture (Fig. 1).

Effect of different doses of LLLT on IGF-I mRNA expression. MC3T3-E1 cells were exposed to different doses of LLLT (0, 0.96, 1.91, or 3.82 J/cm2). The gene expression of IGF-I was measured for ≤ 48 h by using real-time PCR. Each bar indicates the mean ± SD of three separate experiments. *p < 0.05, LLLT vs. controls.
Effect of LLLT on IGF-I protein expression
We determined that the optimal irradiation dose inducing the highest IGF-I expression was 1.91 J/cm2. Therefore, we examined the expression of IGF-I protein with Western blot analysis at a dose of 1.91 J/cm2. The expression of IGF-I protein increased in a time-dependent manner from 24 to 48 h after LLLT (Fig. 2).
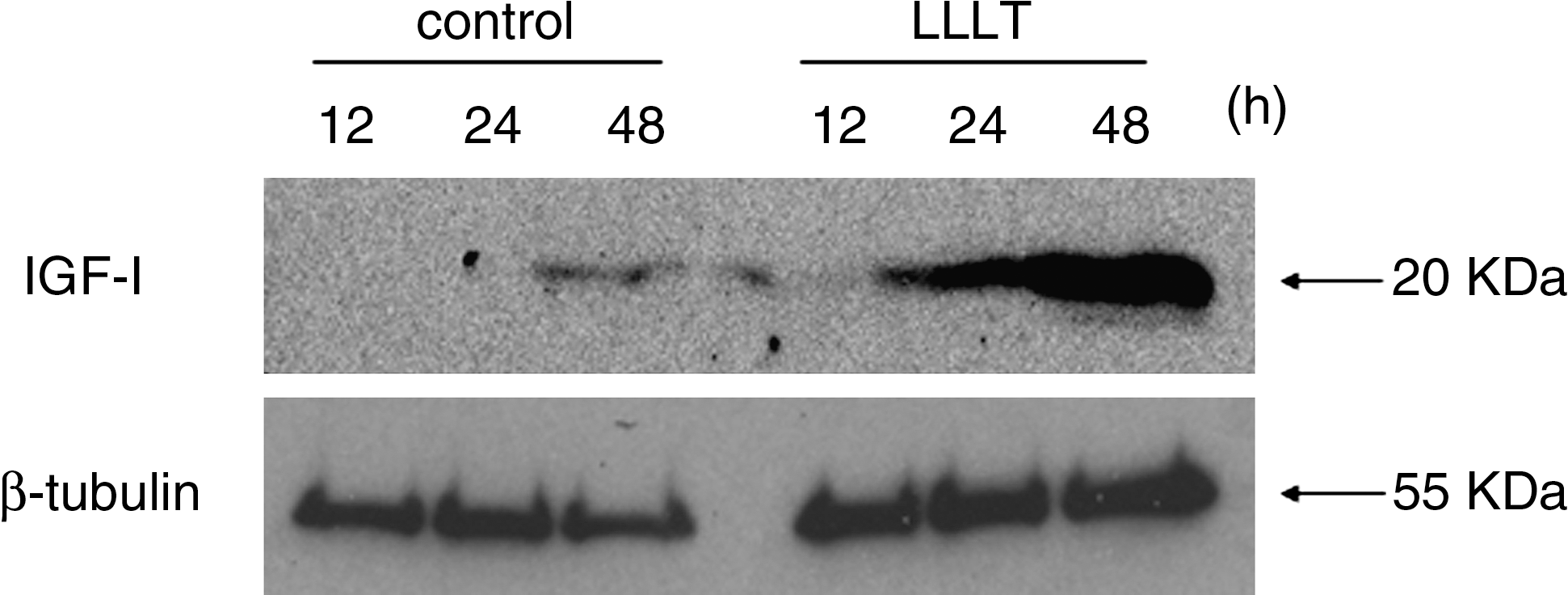
Effect of LLLT on IGF-I protein expression after the cells were exposed to LLLT (1.91 J/cm2). Protein expression of IGF-I was determined by using Western blotting at 12–48 h.
Effect of LLLT or PPP on the IGF-I and Runx2 expression
To examine the autocrine action of IGF-I, we studied the effects of LLLT and PPP on IGF-I and Runx2 expression by using real-time PCR and Western blot analysis. LLLT significantly stimulated IGF-I protein (Fig. 3a) and gene (Fig. 3b) expression at 48 h, whereas the addition of PPP partly abolished the IGF-I expression stimulated by LLLT.
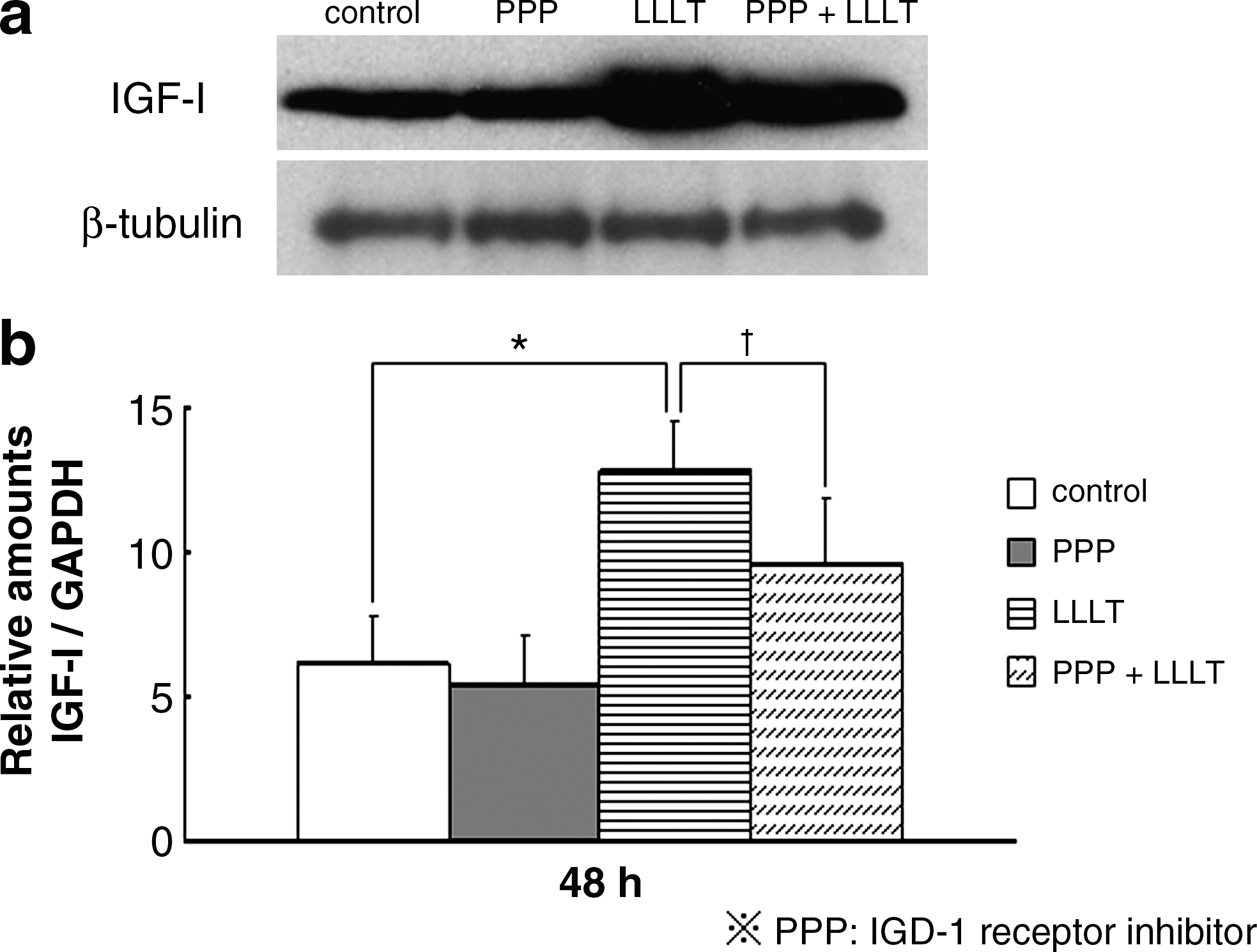
Effect of IGF-I receptor inhibitor (PPP) on the LLLT-induced IGF-I protein (
Additionally, LLLT significantly stimulated Runx2 protein (Fig. 4a) and gene (Fig. 4b) expression, whereas the addition of PPP partly abolished the Runx2 expression stimulated by LLLT.
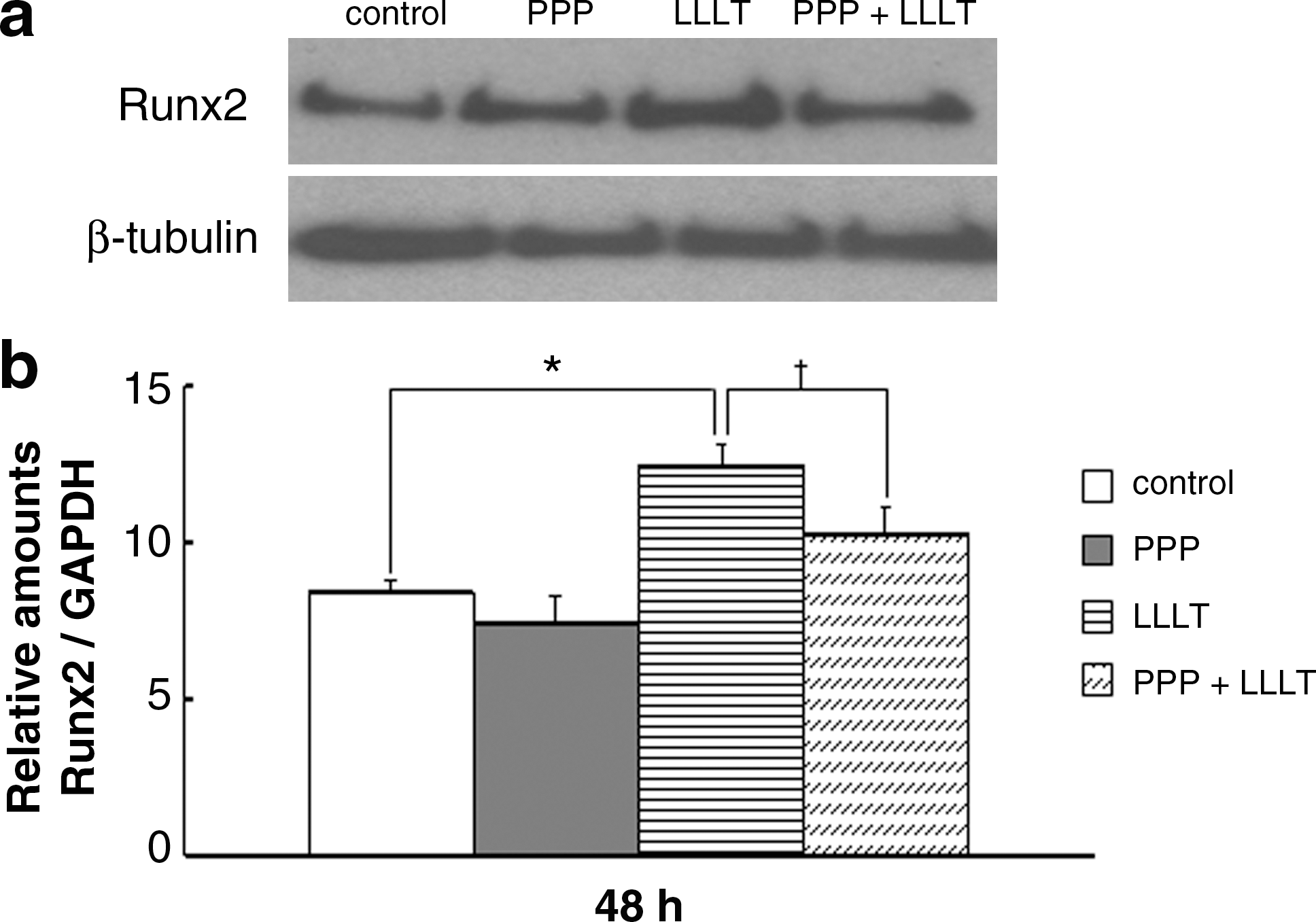
Effect of IGF-I receptor inhibitor (PPP) on the LLLT-induced Runx2 protein (
Effects of LLLT and PPP on the phosphorylation of ERK
We examined the effects of LLLT and PPP on the phosphorylation of ERK-1 (p-ERK) by using Western blot analysis. p-ERK was not seen without LLLT (Fig. 5a, b), whereas it was significantly stimulated 5 to 60 min after LLLT (Fig. 5c). However, the stimulation of p-ERK by LLLT was partially abolished by the addition of PPP (Fig. 5d).
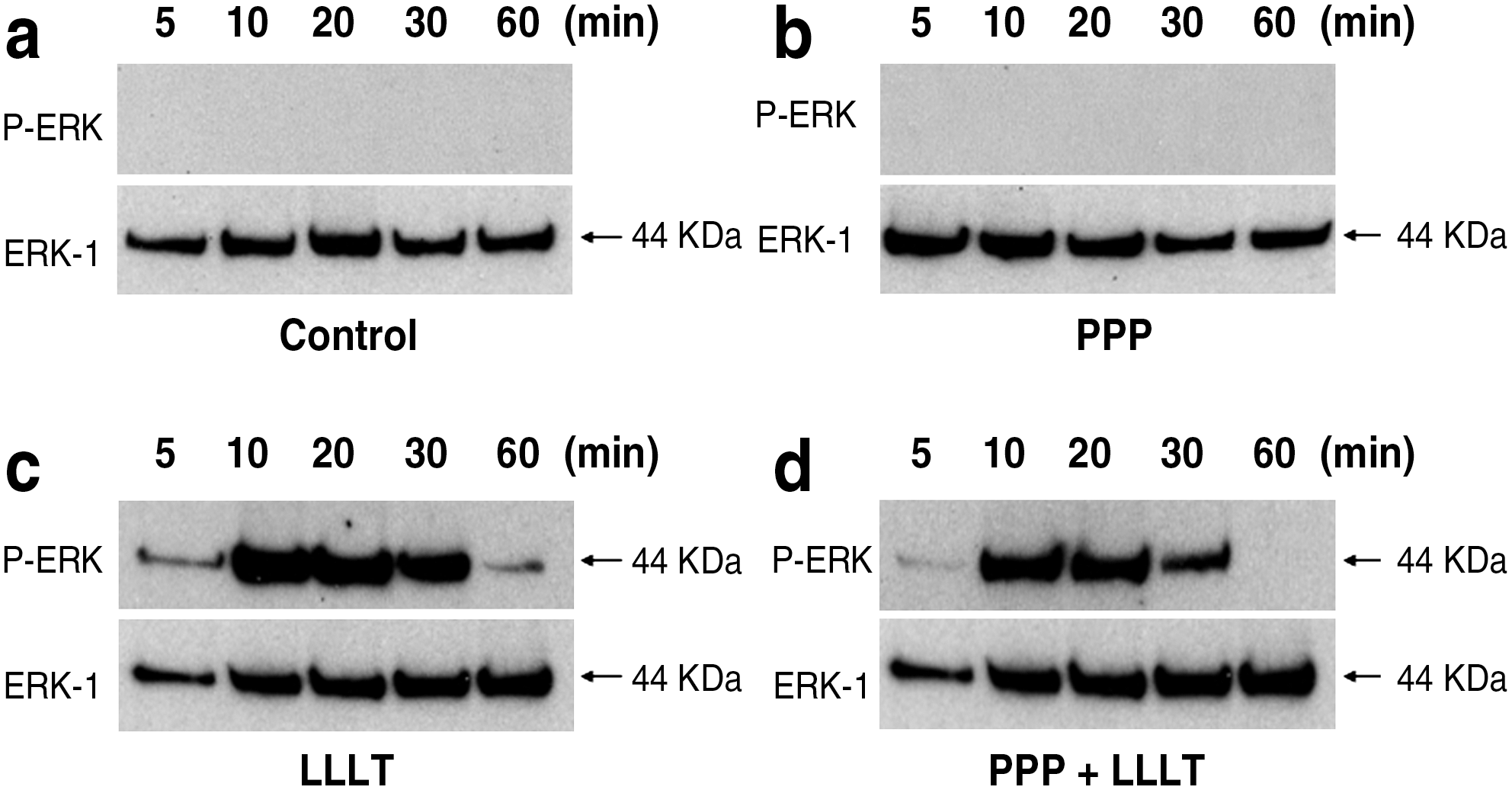
Effect of the IGF-I receptor inhibitor PPP on the LLLT-induced phosphorylation of ERK. P-ERK was not seen without LLLT (
Calcium content of mineralized nodules
When the conditioned medium from the cells subjected to LLLT at 1.91 J/cm2 was added to the MC3T3-E1 culture, the calcium content in the mineralized nodules increased significantly (twofold higher), compared with the controls (Fig. 6a–c). PPP or noggin (a BMP-receptor antagonist) partly inhibited the LLLT-induced change in calcium content in the mineralized nodules (Fig. 6a, b). Moreover, the simultaneous addition of PPP and noggin inhibited most of the LLLT-induced change in calcium content to the control level (Fig. 6c).

Inhibitory effect of PPP and noggin on the increased calcium content of mineralized nodules after LLLT. The cells were cultured with LLLT conditioned medium containing PPP (
Discussion
Previous studies demonstrated that LLLT increases the expression of markers of bone phenotypes in vitro, such as type I collagen, 12,13 ALP activity, 14 and bone sialoprotein. 12 We also reported that LLLT stimulated bone formation in the midpalatal suture during rapid palatal expansion in rats, 15 bone nodule formation through increased ALP activity, 4 and osteocalcin expression in vitro. 5 Furthermore, these phenomena occurred through IGF-I production. 6 However, the signal transduction downstream from IGF-I triggered by LLLT has not been elucidated. Therefore, we prepared an in vitro model to investigate the effects of LLLT on the IGF-I–related signal transduction in osteoblasts.
First, we examined the effect of different doses of LLLT on IGF-I mRNA expression. The results indicated that the expression was highest with LLLT at 1.91 J/cm2. The IGF-I protein expression with LLLT was similar to the IGF-I mRNA expression. Furthermore, the LLLT stimulation of IGF-I was partly abolished by the addition of PPP, suggesting that the increased IGF-I in response to LLLT is partly regulated in an autocrine manner.
IGF-I is a polypeptide hormone and potent chemotactic factor that may play a major role in recruiting osteoblasts during bone formation. 7,16 In osteoblast-like MC3T3-E1 cells, IGF-I stimulates the activity of ALP and the signal for osteoblast mitogenesis and survival through various signals, such as extracellular signal–regulated kinase (ERK) and Runt-related transcription factor 2 (Runx2), and some other pathways in MC3T3-E1 cells. 10,17,18 Moreover, Runx2 is a master switch for osteoblastic differentiation, and the expression of Runx2 is critical for bone formation. 19,20 Therefore, Runx2 and ERK appear to be target genes of the IGF-I signaling pathway and act as important regulators of the process of osteoblast differentiation. To confirm the downstream signaling of LLLT-induced IGF-I, we examined the effect of the optimal dose of LLLT on the expression of Runx2 and the phosphorylation of ERK-1. Both were increased significantly by LLLT, suggesting that IGF-I stimulated by LLLT accelerates downstream signals such as Runx2 and ERK phosphorylation.
The IGF-I receptor consists of two identical extracellular α-subunits that are responsible for ligand binding, and two identical β-subunits with a transmembrane domain that serves as an intracellular tyrosine kinase. 21 PPP prevents IGF-I signaling by binding to the IGF-I receptor, specifically blocking the phosphorylation of tyrosine kinase (Y1136) on the β-subunit of the IGF-I receptor. 11 IGF-I binds to the IGF-I receptor, and its signal is mediated by the phosphorylation of receptor-regulated ERK. 22 We found that LLLT clearly stimulated downstream IGF-I signaling, such as Runx2 and ERK phosphorylation, and that the addition of PPP partly inhibited the LLLT-induced Runx2 expression and phosphorylation of ERK. LLLT treatment of osteoblasts led to increased IGF-I and Runx2 expression after 24 and 48 h, respectively, whereas ERK phosphorylation was detected 5 min after LLLT. A similar phenomenon has been described: ERK phosphorylation was detected within minutes of treating an osteoblast cell line with fibroblast growth factor-2, whereas Runx2 phosphorylation did not occur for several hours. The time lag between ERK phosphorylation and Runx2 expression suggests that several steps exist between these events and may include the activation of other kinases. 23
Recently, we reported that LLLT induced mineralization through increased bone morphogenetic protein (BMP) production by MC3T3-E1 cells. 24 Because MC3T3-E1 cells produced both IGF-I and BMP in an autocrine manner, 25 we examined the role of stimulation of these factors by LLLT on the calcium content of mineralized nodules. As expected, the calcium content of the cell culture was increased by adding LLLT-conditioned medium. In contrast, the increase in calcium content was only around one-half with the addition of PPP-treated conditioned medium or noggin-treated conditioned medium, whereas simultaneous PPP and noggin treatment inhibited most of the LLLT-induced change in calcium content of the mineralized nodules. These results suggest that IGF-I and BMP are similarly effective at stimulating the calcium content of mineralized nodules in MC3T3-E1 cells, and that IGF-I, BMP, and their downstream signals, such as Runx2 and ERK phosphorylation, were responsible for a large part of the bone nodule formation stimulated by LLLT.
Conclusion
This study demonstrated that LLLT stimulates in vitro mineralization through increased IGF-I and BMP expression through Runx2 expression and ERK phosphorylation, and that IGF-I and BMP were similarly effective at increasing the mineralization of bone nodules. Therefore, IGF-I and BMPs are likely major mediators of bone formation by LLLT.
Footnotes
Acknowledgments
This study was supported by a grant from the Ministry of Education, Culture, Sports, Science, and Technology of Japan to promote multidisciplinary research projects (19592369), by a grant from the Dental Center at Nihon University School of Dentistry, and by a special research grant from the Promotion and Mutual Aid Corporation for Private Schools of Japan.
Author Disclosure Statement
No conflicting financial interests exist.