
Abstract
Introduction
Beyond this, increasing interest is seen in the clinical use of stem cells for transplantation to sites of injury (e.g., spinal cord injury), 2,3 and in treating neurodegenerative disorders (e.g., Parkinson disease). 4 LLLT may be used to increase the numbers of stem cells in culture or to induce differentiation into specific cell types or both. Furthermore, it may be possible to inhibit the growth of tumors by using laser light. Previous reviews summarized relevant work up to 2003 and 2004. 5,6 Both of these reviews included in vivo studies of laser effects on wound healing; the review by Reddy et al. 5 also included studies on pain management. Despite such previous comprehensive reviews, at the present time no therapeutic window for laser dosimetry and mechanism of action has been definitively determined at the level of individual cell types (e.g., diabetic fibroblast cells in culture). 7
This article extends previous reports in this area by critically reviewing those articles published from January 2002 to September 2009. It focuses on experimental studies that investigated laser irradiation of human and animal cells in culture, identifying similarities and differences in these studies, and whether a clear conclusion can be reached from the data presented regarding the influence of laser irradiation on these cells. The cell types chosen for consideration in this article are those involved in wound repair and soft-tissue regeneration [i.e., fibroblasts and macrophages (for granulation tissue formation), endothelial and smooth muscle cells (for angiogenesis), keratinocytes (for skin reepithelialization), and adipocytes (for subcutaneous tissue)]. The studies reviewed include laser effects on cells cultured under nutritional-deficit conditions (cells “stressed”), hyperglycemic conditions, or collected from diabetes patients or experimental animals, as well as cells “wounded” or harvested from wounds. Studies using stem cells that could be used for wound and soft-tissue repair also have been included; human adipose-derived stem cells have been shown to differentiate in the presence of appropriate stimulants into bone, cartilage, fat, or muscle. 8
Laser irradiation is characterized by a number of physical parameters, principally including wavelength, spot size, power, power density, energy, energy density, duration of irradiation, frequency of irradiation, and interval between consecutive irradiations. However, at present, the relevance of these parameters to the putative healing effects of laser irradiation on different injuries and skin conditions remains unclear. Results from several previous investigations are remarkably contradictory, which may be in part a consequence of the plethora of possible combinations of irradiation parameters, and the inability to measure the possible effects after irradiation with the necessary objectivity. 9,10 The potential effects of variation of irradiation frequency and dosing intervals have been rarely considered. Similarly, the possibility that the photobiomodulatory effects of laser irradiation are dissimilar across different cell types and species has been largely ignored as a potential explanation for the conflicting results reported in the literature after LLLT treatment. 11 The current review was informed by the recommendations of Tuner and Hode, 12 who previously identified important factors to be taken into consideration when examining such studies.
It is important to recognize that cell cultures and tissues have different optical properties: cell cultures provide a low scattering medium in which laser light may have almost the same effect as incoherent light [i.e., color-filtered light from a light bulb or a light-emitting diode (LED) of the same wavelength]. In contrast, tissue is a highly scattering medium, and thus laser light provides results superior to those of incoherent light of the same parameters. 12
The aim of this article was therefore to review experimental studies of laser irradiation of human and animal cells in culture, to assess the possible photobiomodulatory effects of such irradiation in terms of cell viability and proliferation. The cells chosen were those involved in wound or soft-tissue repair or cell lines relating to soft tissues. The current review also included assessment of (a) the quality of the study designs and protocols used; (b) the experimental protocols and laser treatments used, and the appropriateness of these to clinical applications in humans; and (c) the relevance of irradiation parameters to any observed effects.
Methods
A systematic review of the relevant literature in the area was performed. Original research articles investigating the effects of laser therapy on human and animal cells in culture, and published from January 2002 to September 2009, were retrieved and used for this review. Relevant articles were sought and obtained from library sources and the online databases PubMed and Medline by using EndNote X1 (Thomson Corporation).
Search terms were “cell proliferation,” “laser therapy,” “laser irradiation,” “laser phototherapy,” and “phototherapy.” Additional secondary sources of information included reference lists from retrieved articles, and pertinent articles identified by hand searches of relevant journals not found from the databases (e.g., Laser Therapy).
We included studies that met the following criteria: Laser (or other monochromatic light source) was investigated as the primary intervention (independent variable); The type of laser and precise wavelength were defined or implied; At least one outcome or index of cell proliferation or differentiation was reported; Studies performed with human and/or animal cells or cell lines in culture; and Studies relating to wound repair or soft-tissue regeneration.
Studies excluded from this study were the following: Articles published prior to 2002;
In vivo studies involving whole animals, not isolated cells or cell lines; Studies reported in languages for which no English-language translation was available; Studies performed with cells from animals other than common laboratory animals; Studies involving cells or tumor cells isolated from hard tissues (e.g., osteoblasts, osteoclasts, osteosarcoma cells), skeletal muscle cells, nerve cells; Reviews and meta-analyses; Studies involving multisource/multiwavelength arrays, photodynamic therapy, high-power laser, flashlamp pulsed-dye laser; Studies involving confocal laser microscopy, laser-scanning cytometry, laser-capture microdissection, laser light scattering, matrix-assisted laser desorption/ionization/time-of-flight mass spectrometry; and Studies for which only an abstract was available.
EndNote searches were carried out independently by two of the authors (PP, TYC), and the findings were confirmed by a third person (BR). Articles for inclusion and exclusion were identified independently, and confirmed, thereby minimizing bias.
For included articles, the following data were extracted and tabulated by two of the authors (PP, TYC) and confirmed by a third person (BR):
research method: including controls; minimizing variability in experimental conditions because of scattering of laser light and a reduction in energy when laser light is transmitted through culture medium; culturing of cells to a subconfluent monolayer, and ensuring that the whole of the cell monolayer is irradiated; measurement of power density and energy density at the level of the cell monolayer;
species type: human or animal cells; source and, for the latter, the species;
description of cells: normal*, stressed (nutritional deficit), wounded, diabetic, tumor cells, other cell lines; group sizes, numbers of replicates;
laser-treatment parameters;
experimental outcomes;
authors' conclusion: results of laser irradiation.
[*Normal cells are those that have been grown in the presence of an adequate concentration of serum (usually 10%) to support cellular proliferation and are not cells obtained from wounds or tumor cells.]
Studies were then critically reviewed in terms of study design, methods (PP and TYC), and appropriateness of irradiation parameters (GDB); comments on these study characteristics also were summarized and tabulated (as bold text within tables).
Results
The human and animal cell types irradiated in the searched studies have been limited to those cells involved in wound or soft-tissue repair or cell lines relating to soft tissues and are summarized in Table 1. Cell types included normal cells; cells incubated in culture medium deficient in FBS (i.e., serum starved = nutritional deficit, with cells being “stressed”); cells incubated in culture medium with a high concentration of glucose (i.e., hyperglycemia) or obtained from diabetic animals (“diabetic cells”); cells incubated in culture medium to confluence and scratched with a needle (i.e., “wounded cells”); cells incubated in culture medium to confluence, medium replaced with one containing a high concentration of glucose, and cells scratched with a needle (i.e., “diabetic wounded cells”); and cell lines, mostly carcinoma.
NF, not found in searched studies from January 2002 to September 2009.
Cell types included are those involved in wound and soft-tissue repair; cell lines, mostly carcinoma.
Results from the literature search are summarized in Fig. 1; in total, 46 publications were included in this review 1,7,13 –56 and are summarized in Tables 2 –13.
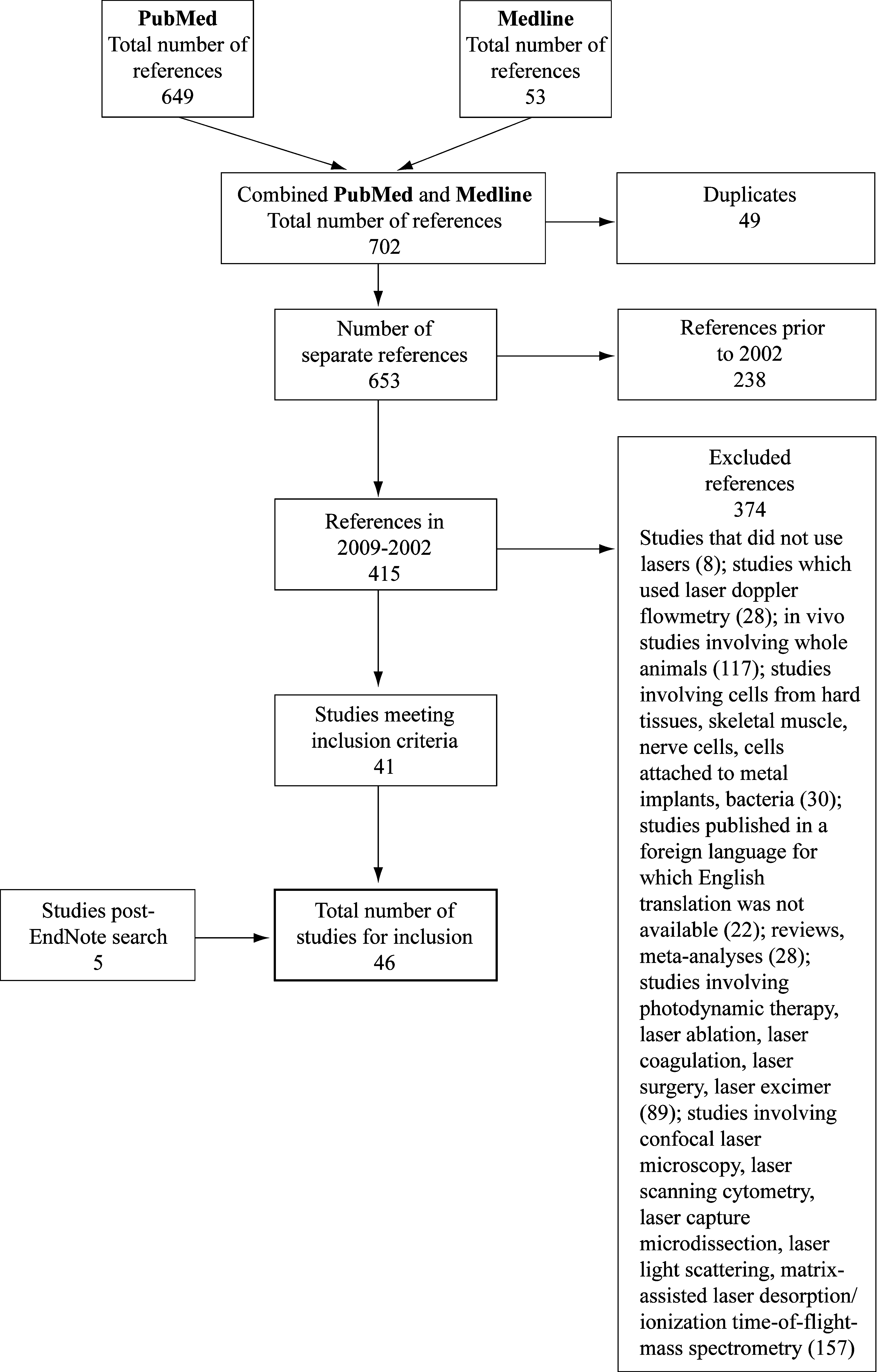
Flowchart of literature search: Studies with human and animal cells.
Fluence rate or flux; **fluence.
In this and subsequent tables, the concentration of FBS and stimulators in culture medium has been given but not of antibiotics or other additives.
The outcomes reported in the retrieved studies included cell proliferation, including measurement of cell numbers and cell viability; membrane integrity; DNA damage; differentiation of stem cells into specific cell types (e.g., mesenchymal stem cells into myoblasts).
The following laser parameters were recorded from the searched studies: wavelength, power, power density (irradiance), energy, energy density (radiant exposure), spot area or irradiated area, method of irradiation, number of irradiations, duration of irradiation, interval between irradiations, and time after final irradiation for outcome measurements. These parameters are summarized in Tables 2 –13. Comments on study design and findings are presented in a row under each respective study.
The main findings are summarized in the following sections.
Application of laser light to cells in searched studies
Considerable variation was found in how irradiation was performed; this included the following: Irradiating cells from above or below, with a constant distance between the laser source and cell monolayer in dishes, wells, or plates; Sheltering adjacent wells from scattered laser light with a conical application tip for the laser diode, using an aluminum foil lid with an appropriate-sized hole for the laser beam to pass through, filling adjacent wells with a colored solution, leaving adjacent wells unfilled, and positioning dishes under the laser beam on a black surface or inside a black mask; Minimizing reflection of laser light by blackening inner walls and bottoms of wells, and removing the plastic lids covering the plates or dishes; Minimizing absorption of laser light when irradiating cells from above by removing the plastic lids; and Ensuring precise irradiation over an entire culture well by use of a transparent grid composed of a number of squares or circles and placed at the bottom of the plate.
The laser-beam diameter was adjusted so that the entire area of the dish, plate, or well was irradiated. This was achieved by using an optical lens system to expand or clip the beam. Irradiation of cells from above was often performed by using an optical fiber, whereas in some studies, irradiation from below was carried out by using a contact mode with the tip of the laser diode positioned against the bottom of the dish or plate. Only for some of the studies was power output reported to be measured at the level of the cells. This had been done when irradiating from above by using a power meter placed at the level of the cells or with a fiberoptic detection system placed below the tray during irradiation. However, it is not clear in these studies whether measurement was performed with culture medium present in the wells. Irradiating cells from below minimizes the absorption or reflection of laser light through the culture medium. Brondon et al. 38 reported attenuation of <5% of the full output when irradiating cells through the bottom of the culture plates with laser light of 670 nm. Some studies also measured the irradiated area and power at the level of the cell monolayer. In many studies, the laser irradiation was carried out in the dark or partial darkness to minimize the effect of ambient light. Phenol red indicator was often omitted from the culture medium on account of its having an estrogen-like effect. 57
Irradiation parameters
A wide range of irradiation parameters (and combinations of such parameters) were used in the studies reviewed; relevant parameters were not always provided, and in a number of instances, errors in calculation of dosages were made (see Tables 2 –13). Furthermore, with the exception of wavelength and–particularly–energy density (radiant exposure; see later), in many cases, little evidence was found of a systematic approach to the investigation of the relevance of parameters to light-induced effects (i.e., manipulation of a single parameter while controlling others). This, coupled with the almost exclusively positive outcomes reported in these studies, confounds identification of simple relations between irradiation parameters and observed effects.
Wavelength
Studies used a wide range of wavelengths from 532 (green) to 2,940 nm (infrared); however, the majority of studies used red-light sources (632.4–670 nm; 32 of 46, including those studies that used different wavelengths), which reflects clinical practice and recommendations for treatment of open wounds. With the exception of two studies that used a Q-switched Nd:YAG laser (532 nm, green) and a diode system (808 nm; infrared), respectively, which caused reduction of cell fluence in a dose-dependent manner, 21,37 all other wavelengths investigated (including green, red, and infrared wavelengths) demonstrated the ability to stimulate cellular proliferation in a variety of cell types. A number of studies systematically compared wavelengths (i.e, by controlling other irradiation parameters) and provided evidence of wavelength-dependent stimulatory effects in favor of red wavelengths. 29 –31,35,41,42,44,47
Power
Where reported, laser-power output varied widely, from 0.6 to 7,500 mW (7.5 W), with higher powers typically associated with the use of infrared sources (e.g, Nd:YAG laser 1,064 nm in four studies that used 1,000 mW). This notwithstanding, 12 (26%) of 46 studies failed to report details on this parameter, and of these, only four provided sufficient detail to allow its estimation; additionally, limited evidence was reported of regular checking of power outputs of devices used, and, in two cases, calculation of power based on other reported parameters did not match reported values (see Tables 2 –13).
Only one study systematically investigated the relevance of this parameter to observed effects: this found that irradiation with red light (660 nm) by using 20 mW produced superior stimulatory effects on dental-pulp stem cells compared with irradiation at 40 mW (3 J/cm2). 16 However, given that other relevant parameters were standardized (i.e., spot size and energy density), this may better reflect an irradiance-specific effect.
Irradiance
Irradiance (or more simply, “power density”) is calculated from power output (mW) and spot size or area of irradiation (specified in square centimeters). The latter is usually based on a mixture of measurement, manufacturer's specification, or estimation (e.g, from beam characteristics or surface of well plate). Thus, although power output can be measured with some degree of accuracy, spot size represents a potential area for error (or at least assumptions), which may in turn affect accuracy in the calculation of irradiance. Reported spot size or area of irradiation varied widely in the reviewed studies, from 0.008 to 78.5 cm2, largely reflecting the range of experimental setups used (e.g., configuration of cell-well plates). For irradiance, a wide variation in reported values was found (from 1.16 to 428.57 mW/cm2); however, 26 (57%) of 46 studies did not report this parameter. This notwithstanding, a further 18 studies provided sufficient information to allow calculation of irradiance: taking these into account, irradiance ranged from 0.14 to 1,714 mW/cm2.
The relevance of irradiance to stimulatory effects was specifically investigated in two studies: in the first, higher irradiances (20–65 mW/cm2) were associated with greater stimulatory effects at a wavelength of 670 nm. 17 A second study (660 nm) using higher irradiances found an inverse relation, with a lower irradiance providing a superior stimulatory effect (143 versus 429 mW/cm2). 24 Taken together, these findings are suggestive of a putative irradiance “window” for stimulatory effects.
Energy
Energy is calculated from power output (mW) and irradiation time (specified in seconds). Time of irradiation ranged from 2 to 13,793 s (>3 h and 48 min), reflecting not only the range of dosages investigated but the variation in power outputs used in different studies; it is important to note that the higher irradiation times used in these studies are somewhat artificial in terms of clinical practice. Although energy was reported in only three articles (0.12–23.4 J), this parameter could be calculated for 33 of the remaining studies. Whereas energies varied widely (0.018–1,257 J), the relevance of these was not specifically investigated by any study.
Radiant exposure
Radiant exposure (or energy density) is generally considered the most appropriate means of specifying dosage, at least in experimental studies. It is calculated from irradiance and time of irradiation, and specified in joules per square centimeter. This parameter was reported in all but one of the studies reviewed, 23 and varied between 0.04 and 20.79 J/cm2. Although differences in experimental setup and calculation of this parameter preclude identification of a definitive figure (or window) of radiant exposure for stimulatory effects, consistent evidence from those studies compared radiant exposures of a dosage-dependent effect on measured parameters of cellular proliferation: this included a threshold for effect, increasing stimulation effect with increasing dosage, and a plateau of effect or inhibition at the highest dosages. 1,3,5,8,19 –21,25,28,29,33 –36,43,45,49 –53,55
Irradiation regimen
Considerable variation was found in the number of irradiations used and the interval between successive irradiations. The former ranged between one and seven doses, often with two or three doses being given on the same day, whereas the time interval between successive irradiations varied between 6 and 72 h. This notwithstanding, only one study directly compared different irradiation regimens (different intervals between two standardized dosages): this found superior results in terms of cell viability and wound closure with irradiation at 30 min and 72 h, versus 30 min and 24 h (632.8 nm; 5 J/cm2). 7
Measurement of Outcomes
Time after final irradiation
The times at which outcomes were measured after irradiation (or the final irradiation if several irradiations were applied) varied from 0 to 144 h.
Cell proliferation
Data from the searched studies were critically analyzed to determine whether specific cell types of human and animal origin were influenced in a similar way by laser light of a particular wavelength, and with a similar power density and energy density. Unless stated otherwise, cells were irradiated once. Proliferation rates of irradiated cells were compared with those of nonirradiated cells (controls).
Stem cells
Human and animal studies (Tables 2 and 8) were not able to be directly compared, as dissimilar cell types had been irradiated. A significantly increased proliferation occurred in response to irradiation at 635 nm with 5 J/cm2 for human adipose-derived stem cells at 24 and 48 h, cultured in the presence of EGF, but in the absence of EGF, the increase was not significant. For human embryonic stem cells, irradiation at 830 nm with 5 and 8 J/cm2 resulted in a significant increase in differentiated colonies. A significant increase in rat bone marrow–derived mesenchymal stem cells and ventricle-derived cardiac stem cells occurred after irradiation at 804 nm, with 1 and 3 J/cm2 ≤ 2 and 4 weeks after treatment. Irradiation of rat bone marrow–derived mesenchymal stem cells at 635 nm with 0.5, 1, 2, and 5 J/cm2 significantly stimulated proliferation, with 0.5 J/cm2 being the optimal energy density. The latter suggests that proliferation of these animal stem cells is more responsive to low energy density at 635 nm rather than at 804 nm. Furthermore, 5-azacytidine induction of myogenic differentiation was markedly increased by irradiation at 635 nm with 5 J/cm2.
Stressed stem cells
Human dental pulp stem cells grown under nutritional-deficit conditions and irradiated at 660 nm with 3 J/cm2 (two irradiations with a 6-h interval, possibly giving a cumulative effect equivalent to a single irradiation of 6 J/cm2) had significantly more viable cells than did controls grown under the same nutritional conditions at 24 h after the first irradiation. Owing to human stem cells of different origin being irradiated, no inferences can be made regarding whether stressed stem cells are more responsive to laser irradiation than are nonstressed cells. No animal studies were found for 2002 through 2009.
Endothelial cells
Power and energy density–specific effects were found for human umbilical vein endothelial cells irradiated at 670 nm; proliferation was increased in a dose-dependent manner when irradiated at 2, 4, and 8 J/cm2 with a power density of 20 mW/cm2 (Table 3). Irradiation at 8 J/cm2 with 20 and 65 mW/cm2 increased cell proliferation, but no significant effect on proliferation was seen at 8 J/cm2 with 10 mW/cm2. Mouse aortic endothelial cells showed increased proliferation after exposure to all chosen wavelengths from 625 to 675 nm at 10 J/cm2, with maximum stimulation at 655 nm, and did not increase with longer visible red wavelengths (Table 9). This was similar to that reported previously for bovine aortic endothelial cells. 58
Stressed endothelial cells
Human umbilical vein endothelial cells grown in a nutritionally deficit medium showed increased cell proliferation at 96 and 120 h after irradiation at 632.5 nm with 0.26 J/cm2. These findings suggest that these cells might be more responsive to laser light than normal human endothelial cells. Bovine aortic endothelial cells grown in a low-glucose medium had an increased cell proliferation at 48 h after irradiation at 632.5 nm with 0.54 and 1.08 J/cm2. These findings suggested that these cells might be more responsive to laser light than were normal human and bovine endothelial cells.
Smooth muscle cells
No human or animal studies were found in the search carried out for 2002 through 2009.
Stressed smooth muscle cells
No human studies were found in the search carried out for 2002 through 2009. Pig aortic smooth muscle cells grown under nutritionally deficit conditions and irradiated at 780 nm with 1 and 2 J/cm2 showed increased proliferation with 2 J/cm2 at 24, 48, and 72 h (Table 10).
Keratinocytes
No human or animal studies were found in the search carried out for 2002 through 2009.
Monocytes
Irradiation of human peripheral blood monocytes at 632.8 nm with 2.55 and 5.10 J/cm2 significantly increased proliferation, with 2.55 J/cm2 giving a significantly higher stimulation than 5.10 J/cm2. Proliferation of cells irradiated at this wavelength with 7.64 J/cm2 was not significantly increased. PHA increased the proliferation of laser-irradiated cells (Table 4).
No animal studies were found in the search carried out for 2002 through 2009.
Fibroblasts
Irradiation of human skin fibroblasts at 632.8 nm with 8, 10, and 16 J/cm2 showed a dose-dependent effect, with increasing energy density causing a greater increase in numbers of cells; cell proliferation was highest at 16 J/cm2 and decreased at the lower energy densities (Table 5). Human periodontal ligament fibroblasts irradiated at 809 nm with 1.96, 3.92, and 7.84 J/cm2 also showed an increased proliferation. Mouse skin fibroblasts showed increased proliferation when irradiated at chosen wavelengths between 625 and 675 nm at 10 J/cm2. The fibroblast proliferation increased at longer wavelengths, with a maximum at 665 and 675 nm (different from mouse endothelial cells); exposure to 810 nm was inhibitory (Table 11). At 24 h after the third irradiation, chicken 8-day-old embryo fibroblasts gave the higher cell proliferation when irradiated once per day for 3 consecutive days at 570 nm (green LED) with 0.1 J/cm2 and at 660 nm (red LED) with 0.53 J/cm2, with that brought about at 570 nm being higher than that at 660 nm. This indicated a marked difference between the responses of mouse skin fibroblasts and chicken embryo fibroblasts, with the latter showing more similarity to rat mesenchymal stem cells regarding their response to laser light at low energy density.
Stressed fibroblasts
Proliferation of human gingival fibroblasts grown in nutritional-deficit conditions was significantly increased after irradiation at 660 nm with 2 J/cm2; fibroblasts were irradiated twice with a 12-h interval (Table 5). The highest cell growth was found by using the least power density (143 mW/cm2), and only cultures in this group reached confluence in 6 days after irradiation. Cultures irradiated with the highest power density (429 mW/cm2) had a significant reduction in cell numbers on the sixth day after irradiation. It would appear that stressed fibroblasts were more responsive to laser irradiation at 2 J/cm2 than were normal fibroblasts, for which much higher energy densities were necessary.
In a previous study, it was shown that human gingival fibroblasts grown in nutritional-deficit conditions and irradiated with 2 J/cm2 at 670, 692, 780, and 786 nm had similar or higher cell growth than did normal cells grown in ideal conditions, and that irradiation at 780 nm (50 mW) induced higher proliferation than at the lower wavelength, 670 nm (10 mW), by using the same energy density. 59
A further study with human gingival fibroblasts grown in nutritional-deficit conditions and irradiated with 3 J/cm2 at 904 nm showed that procollagen was not altered by the irradiation, but a significant reduction in protein synthesis was found. A previous study had shown that irradiation of 3 J/cm2 was able to induce cell proliferation. Furthermore, irradiation of human gingival fibroblasts grown in nutritional-deficit conditions at 2,940 nm with 1.68, 2.35, and 3.37 J/cm2 showed that proliferation was increased at 3 days after irradiation.
A study using human dental-pulp fibroblasts grown to confluence and then grown in nutritional-deficit conditions and irradiated at 810 nm showed no significant changes in cellular DNA synthesis measured with [ 3 H]-thymidine uptake at 2 h after irradiation.
However, it is known that confluent cells have minimal or zero proliferation rate.
Rabbit synovial HIG-82 fibroblasts grown in nutritional-deficit conditions and irradiated with 4.8 or 7.2 J/cm2 at 660 nm had significantly increased cell proliferation at 24 h after irradiation. Irradiation of these cells at 1.2 J/cm2 had no effect on proliferation (Table 11). Mouse embryonic NIH 3T3 fibroblasts in nutritional-deficit conditions and irradiated twice with a 6-h interval with 3 or 4 J/cm2 at 904 nm had increased cell growth at 144 h after irradiation, but 5 J/cm2 was without effect. These studies would suggest that stressed fibroblasts are responsive to laser light over a certain dose range.
Diabetic fibroblasts
No human studies were found in the search carried out for 2002 through 2009. Irradiation of skin fibroblasts from STZ-induced diabetic rats at 632.8 nm with 4 J/cm2 at 2 or 4 days significantly increased proliferation (Table 12). Chicken 8-day-old embryo fibroblasts irradiated at 570 nm with 0.3 J/cm2 and maintained in a hyperglycemic medium (17 mMglucose) showed a significantly higher proliferation rate.
Wounded fibroblasts
Human skin fibroblasts wounded before irradiation and treated with 5 J/cm2 at 632.8 nm on day 1 and again on day 4 had increased cell proliferation at 1 h after the final exposure (Table 6). Complete closure occurred in wounded fetal fibroblast cells irradiated at 5 J/cm2 compared with nonirradiated cells. Irradiation with 5 J/cm2 at 632.8 nm resulted in a stimulatory effect that was more effective than that at 830 or 1,064 nm, indicating that these cells were more responsive to laser stimulation at lower wavelengths. This is an important difference compared with human stressed fibroblasts, which were stimulated more at the higher wavelength. Wounded cells exposed to 5 or 16 J/cm2 at 1,064 nm showed an increase in alkaline phosphatase enzyme activity when compared with normal cells exposed to the same dose. Significant differences were found for alkaline phosphatase enzyme activity for 5 and 16 J/cm2 between the different wavelengths, providing evidence that dose and wavelength influence the stimulatory or inhibitory effect of laser irradiation. One possible explanation for cells being less responsive with 1,064 nm is that less laser energy is absorbed by the cell monolayer (irradiation at this wavelength has a greater penetration depth). Two further studies involving irradiation of human wounded skin fibroblasts with 5 J/cm2 at 632.8 nm showed an increase in cell viability after 1 h and an increase in cell proliferation after 24 h. A duration of between 1 and 3 h after irradiation was sufficient in assessing the direct effect of laser irradiation on cells (cell viability, ATP mitochondrial activity, membrane integrity, DNA damage), but a longer duration of 24 h was required to demonstrate the indirect effect of laser on cells (cell proliferation, protein expression). Furthermore, a decrease in cell proliferation occurred after one dose of 2.5 or 16 J/cm2, two doses of 5 or 16 J/cm2, and three doses of 5 or 16 J/cm2 at 632.8 nm, indicating a bioinhibitory effect dependent on dose (2.5, 5, or 16 J/cm2) or number of doses. Two studies showed that irradiation of these cells with 10 and 16 J/cm2 at 632.8 nm caused a decrease in cell viability and cell proliferation, with a significant amount of damage to the cell membrane and DNA.
No animal studies were found in the search carried out for 2002 through 2009.
Diabetic wounded fibroblasts
Human skin fibroblasts grown in a hyperglycemic medium (additional 17 mM
No animal studies were found in the search carried out for 2002 through 2009.
Cell lines
The number of human melanoma A2058 cells was found to be increased after 3 days for groups irradiated at 632.8 nm with 1 and 2 J/cm2 (Table 7). For human epithelial cell line HEp-2, irradiation at 670 nm with 5% FBS at doses of 0.04 to 4.8 J/cm2 resulted in a significantly increased proliferation, whereas irradiation at this wavelength with 10% FBS did not cause any significant change in proliferation. Irradiation of the cells at 635 nm under both nutritional states did not stimulate proliferation. Furthermore, when cells were irradiated at 670 nm with 5 J/cm2 treatment for 50 J/cm2 total dosage, the highest rate of cellular proliferation was produced by the two treatments per day regimen at 6 days after treatment. The four treatments per day regimen resulted in a significant decrease in proliferation after a moderate early phase increase at 48 h from the start of irradiation; the one treatment per day regimen had a minimal effect on cellular proliferation. Similar findings were obtained for mouse subcutaneous connective tissue cell line L-929, irradiated at 670 nm with 5 J/cm2 treatment for 50 J/cm2 total dosage (Table 13). Thus, the empiric use of a single treatment per day dosing frequency as a treatment strategy for all cell lines and tissues might explain conflicting reports on the effects of laser irradiation (i.e, both positive and negative effects) and thus why the efficacy of LLLT remains controversial. Identification of the most appropriate treatment frequency for a particular cell line or tissue is crucial to achieve the desired result, whether photobiostimulation or photobioinhibition.
Human epithelial adenocarcinoma (HeLa line) and lymphoblast (TK6 line) were irradiated at 808 nm continuous-wave laser and 905-nm pulsed-wave laser with energy densities ranging from 1 to 60 J/cm2. HeLa cell proliferation was not affected by irradiation with each dose of continuous-wave diode, 808 nm, but was stimulated after irradiation with pulsed wave of 905 nm and with simultaneous irradiation with 808 and 905 nm. At day 2 after irradiation, an increase in cell number was evident after 30 J/cm2 of combined laser. TK6 cell proliferation was only slightly inhibited after irradiation with continuous, pulsed, or combined laser light, which was more evident at day 5–7 from irradiation with a pulsed diode. A higher proliferation of human laryngeal epithelial carcinoma line was brought about by irradiation at 809 nm with energy densities of 1.96, 3.92, and 7.84 J/cm2. Inhibition of human hepatoma cell lines HepG2 and J-5 proliferation occurred after irradiation at 808 nm, with maximal effect achieved with 90- and 120-s irradiation for HepG2 and J-5 cells (corresponding to 5.85 and 7.8 J/cm2), respectively.
Cell growth of mouse connective tissue cell line L-929 was shown to be stimulated to a greater extent when irradiated at 904 nm with 0.05 J/cm2 than at 6 J/cm2. This growth occurred at 24, 48, and 72 h after irradiation with more growth between 24 and 48 h. When these cells were irradiated with an LED at 670 nm at 5 J/cm2/treatment for 50 J/cm2 total energy dose, the treatment frequency that produced the highest rate of cellular proliferation was the two treatments per day regimen at 144 h after treatment.
It has been suggested that the different cellular effects induced by laser irradiation could be due to the redox potential of target cells, which is associated with stimulation of cell functions if it shifts toward oxidation, and with inhibition, if it moves toward reduction. 60
Stressed cell lines
A dose-dependent effect of irradiation at 632.8 nm on proliferation of human lung adenocarcinoma cells (ASTC-a-1) cultured in nutritional-deficit conditions was shown with energy densities of 0.5, 0.65, and 0.80 J/cm2, with the highest energy density significantly stimulating cell proliferation (Table 7). Proliferation of monkey kidney epithelial cells (Vero cell line) grown in nutritional-deficit conditions was increased when irradiated 3 times at 660 or 780 nm (second and third irradiation with 6-h interval); the growth of cultures irradiated 1 or 2 times was not increased compared with nonirradiated cultures (Table 13). The improvement in cell growth after laser treatment appeared to be independent of the wavelength used, although a trend for a more-pronounced increase in growth rate was noted when 780 nm was used.
Discussion
This review aimed to evaluate experimental studies of laser irradiation of human and animal cells in culture, to assess the possible photobiomodulatory effects of such irradiation in terms of viability and proliferation of those cells involved in wound or soft-tissue repair, or cell lines relating to soft tissues. In total, 46 eligible studies were included in this review, based on a variety of cell types and outcomes of cellular proliferation. Findings were almost exclusively positive in demonstrating the ability of laser irradiation to modulate (usually stimulate) cellular proliferation; furthermore, evidence suggested that such effects were dependent on irradiation parameters, and particularly wavelength and radiant exposure.
Experimental studies on the effects of low-level laser irradiation on human and animal cells in culture should represent an important prerequisite to clinical trials in humans. Such trials provide a means to test different sets of laser parameters on various cell types to determine an optimal set of parameters that photostimulate or photoinhibit cellular proliferation and also protein expression and release. The most important finding from the current review is in relation to the effectiveness of laser therapy as a modality to increase or decrease the cellular proliferation of a very wide range of different cell types and the dependence of this on wavelength, power density, energy density, number of exposures, and interval between exposures. The review has sought to determine whether similar findings have been reported for specific cell types from human and animal sources. The current results strongly support the case for further controlled studies with human and animal cells (e.g., mouse, rat, monkey), especially stem cells, cells grown under nutritional-deficit conditions (“stressed” cells), cells grown in media supplemented with additional glucose (“diabetic” cells), and cells wounded in vitro.
This review also highlights a number of important issues relating to the quality of the study designs and protocols used, the types of cells used (particularly in terms of the appropriateness of these to clinical applications in humans), and the relevance of irradiation parameters. Systematic reviews have developed principally as a means of integrating evidence from randomized controlled trials to assess the clinical effectiveness of a given intervention; although the principles underpinning systematic reviews of randomized controlled trials apply equally to experimental studies, to date, relatively few such reviews have been published of the effects of low-level laser irradiation on specific cell types. A key aspect of systematic reviews of clinical evidence is assessment of the quality (internal validity) of reviewed studies, by using accepted rating scales. Although such scales have yet to be developed and formally agreed on for the assessment of internal validity in experimental studies, assessment of the quality of studies reviewed here has been informed by consideration of the items typically included in scales for randomized controlled clinical studies.
Research design and reporting of studies
Two salient issues limit comparison between studies: replication of reported benefits, and the formulation of recommended parameters for further studies using human and animal cells. These are the lack of specification of key details in many of the reviewed studies (application of a laser beam by using an optical lens system or optical fiber, laser parameters such as wavelength and power density), and the wide variety in experimental methods used (including irradiation of cell monolayers from above or below; steps taken to minimize scattering and reflection of laser light; the influence of ambient light on cells; absorption of laser light through culture medium; the range of irradiation parameters used, including number and timing of irradiations, interval between irradiations, and the time at which experimental outcomes were measured after laser irradiation).
Apart from inadequate reporting of important elements of research design, and errors in calculation of laser-irradiation parameters, several other prevalent weaknesses in the reviewed studies may have influenced some of the reported findings (e.g., incorporation in the culture medium of phenol red, which has been shown to have an estrogen-like effect; lack of measurement of power density at the level of the cell monolayer; laser irradiation of cells not performed in the dark but under ambient lighting; and absorption of laser energy by colored culture medium).
Types of cells used and influence of laser light
The influence of low-level laser irradiation on a large number of cell types involved in wound and soft-tissue repair has been studied, and, for most of these, has been shown to stimulate cellular proliferation. Specific wavelengths and energy densities have been identified for optimal proliferation of cells such as endothelial cells and fibroblasts.
However, no recent studies were found that used lymphocytes. For the most part, it was not possible to compare the findings using human cells with those of animal cells owing to the considerable variations in experimental protocols. However, important differences were found in the response to laser light of different cell types (e.g., fibroblasts, endothelial cells) and also for the same cell type between different animal species (e.g., mouse skin fibroblasts and chicken embryo fibroblasts).
It is generally held that laser light stimulates cells that are growing slowly at the moment of irradiation, and that the proliferation of fast-growing cells cannot be stimulated by laser irradiation. Thus, if cells are fully functional at the moment of irradiation or are growing in a serum-rich environment (usually 10% FBS), no cells exist for laser irradiation to stimulate, and no therapeutic benefit will be observed. 61
Consequently, for cells to respond to laser light, they must be grown in nutritional-deficient conditions (“stressed”) or in the presence of high concentrations of glucose (“diabetic”), both of which markedly decrease cell growth; or to be harvested from patients or animals with diabetes. A further possibility is to use cells (e.g., fibroblasts) that have been grown to confluence and thereby have zero or minimal growth, and to wound these cells by scratching with a sharp pointed instrument, such as the tip of a glass pipette; or alternatively, to use cells harvested from wounds. When wounded or scratched, cell monolayers respond to the disruption of cell–cell contacts with an increased concentration of growth factors at the wound margin and by healing the wound through a combination of cellular proliferation and migration. A large number of studies were found in which “stressed,” “diabetic,” “wounded,” or “diabetic wounded” cells had been irradiated, and the indication was that these cells were more responsive to laser light than were normal cells. Furthermore, laser irradiation of cells in the presence of stimulators (e.g., EGF for human adipose–derived stem cells, PHA for monocytes) resulted in a significant increase in cell proliferation in comparison to cells irradiated in the absence of the stimulator, or cultured with the stimulator but not laser irradiated.
More recently the effects of laser irradiation on stem cells have been examined and shown to promote cellular proliferation and myogenic differentiation of rat bone marrow–derived mesenchymal stem cells in the presence of 5-azacytidine. The growth of a number of human and animal cell lines, mostly tumor cells, was found to be modulated by laser irradiation. With regard to the laser studies using cell lines, it is essential that cell identification and verification be performed before each study. A relatively high frequency of cross-contamination was recently reported among cell lines, 62 and about 18–36% of cell lines used in various studies (including the laryngeal carcinoma cell line Hep-2) were incorrectly designated, with most of the problems encountered related to HeLa cells. 63 –65
It has been proposed that laser light is a stimulus able to induce some cells to divide and others to apoptosis. This would explain why laser irradiation of 3 J/cm2 was able to induce cell proliferation as in one study 52 but reduced protein synthesis in a subsequent investigation. 26 The authors stated that more research is indicated to establish the laser parameters that would stimulate protein synthesis; this would be important clinically in applying low-level laser treatment in the later stages of wound healing when protein synthesis is highly required. 26
Times for measuring outcomes after laser irradiation of cells
It was suggested from studies with human skin fibroblasts that a duration of between 1 and 3 h after irradiation is sufficient when measuring the direct effect of laser irradiation on cells (cell viability, ATP mitochondrial activity, membrane integrity, DNA damage), whereas a longer duration of 24 h is required to demonstrate the indirect effect (cell proliferation, protein expression). 31,32 Alkaline phosphatase (ALP) activity has been used in many of the searched studies as an index of cellular proliferation. A negative correlation was reported between ALP expression and cell growth. 66 However, a recent study suggested that this negative correlation depends on the concentration of growth factors in the medium, and also that an increase in ALP may be related to cellular damage. It was therefore advised that the ALP enzyme-activity assay be used in conjunction with other cell-proliferation assays such as neutral red, optical density, or, more specifically, bFGF expression. 67
Molecular mechanisms of laser-stimulated cell proliferation
The stimulation of proliferation of multiple cell types by low-power laser irradiation is mainly through activation of the mitochondrial respiratory chain and initiation of cellular signaling. Studies on the signaling proteins involved in laser-stimulated proliferation of cells, some of which are regulated by mitochondrial signaling, were recently reviewed. 68 Laser irradiation was found to induce phosphorylation of tyrosine protein kinase receptor (TPKR; such as c-Met, receptor of hepatocyte growth factor), previously shown to activate the MAPK/ERK pathway and promote proliferation of cells. 69 Laser irradiation increases endothelial cell proliferation, migration, nitric oxide secretion, and promotes angiogenesis. 18 During this process, low-power laser irradiation increases endothelial nitric oxide synthase (eNOS) protein expression in endothelial cells, which is inhibited by PI3K inhibitor LY294002 and indicates that the activation of the PI3K/Akt pathway is a critical step for the increased expression of eNOS by laser irradiation. 18 A recent study showed that laser irradiation induces an immediate increase in mitochondrial membrane potential, ATP, and cAMP of melanoma cell line A2058 through enhanced cytochrome c oxidase activity. 36 Laser irradiation subsequently promotes phosphorylation of Jun N-terminal kinase (JNK) resulting in activation of the transcription factor activation protein-1 (AP-1). These findings suggest that the signaling pathway cytochrome c oxidase/mitochondrial membrane potential/ATP/cAMP/JNK/AP-1 is involved in the regulation of melanoma cell proliferation induced by laser irradiation. 36 Low-power laser irradiation could induce the production of reactive oxygen species (ROS), which function as key secondary messengers regulating the activity of various protein kinases. Nonreceptor tyrosine kinases, particularly the Src kinases, are targets of ROS and can be activated by oxidative events. Src family kinases play important roles in regulating fundamental cellular processes, including cell proliferation, attachment, migration, and survival. 70
Clinical Relevance and Further Studies
Laser irradiation has the potential to stimulate the proliferation and migration of cell types that are essential for reepithelialization, angiogenesis, and granulation tissue formation and could be used to stimulate wound repair. Moreover, laser-stimulated proliferation of endothelial cells, fibroblasts, and keratinocytes could be used in the manufacture of biologic skin substitutes to treat patients with severe burn injuries. The retardation of proliferation of tumor cells and endothelial cells by laser irradiation could also be a strategy to control tumor growth and spread in patients.
Further studies are necessary to enable a comparison to be made of the effects of laser irradiation on human and animal cells, and also to study the effects of laser light on the proliferation and protein synthesis of stem cells, which are likely to become increasingly important as new treatment modalities for a wide range of human diseases and injuries.
Conclusions
Findings from this literature review consistently demonstrated the ability of laser or monochromatic light to photobiomodulate (typically stimulate) cellular processes in human and animal cells in vitro, and strongly support the case for further controlled research with such cells.
Considerable variation was found in research design, methods, and laser-irradiation parameters, which limited comparison of research findings between studies using the same cell type. Only limited comparisons were possible for specific cell types between human and animal studies. Inadequate reporting of key details was also prevalent, as well as errors in specification or calculation of key irradiation parameters. These issues must be considered in designing future research in this area.
Author Disclosure Statement
No competing financial interests exist.
Footnotes
Acknowledgments
We acknowledge the invaluable support of Ms. Brigid Ryan in the preparation of the manuscript. Ms. Ryan was supported through the Centre for Physiotherapy Research, University of Otago. No financial support has been received in conjunction with the generation of this report. None of the authors had any conflicts of interest.