
Abstract
Introduction
This paper critically reviews those papers published from January 2002 to September 2009. It focuses on experimental studies that have investigated the gene expression and release of growth factors and cytokines from laser-irradiated human and animal cells in culture, identifying similarities and differences in these studies, and assessing whether a clear conclusion can be reached from the data presented regarding the influence of laser irradiation. The cell types chosen for consideration are those involved in wound repair and soft tissue regeneration, i.e. fibroblasts and macrophages (for granulation tissue formation), endothelial and smooth muscle cells (for angiogenesis), keratinocytes (for skin re-epithelization), and adipocytes (for subcutaneous tissue). The studies reviewed included laser effects on cells cultured under nutritional deficit conditions (“stressed” cells), “wounded” cells, “wounded” cells under hyperglycemic conditions, and tumour cell lines. Studies of stem cells which could be used for wound and soft tissue repair were also included.
The growth factors and cytokines considered in this review are endothelin-1 (ET-1), basic fibroblast growth factor (bFGF), hepatocyte growth factor (HGF), interferon-γ (IFN-γ), insulin-like growth factor-1 (IGF-1), interleukin 1-α (IL-1α), interleukin 1-β (IL-1β), interleukin 6 (IL-6), interleukin 8 (IL-8), interleukin 10 (IL-10), keratinocyte growth factor (KGF), melanocyte stimulating factor (MSH), monocyte chemotactic protein −1 (MCP-1), nerve growth factor (NGF), platelet-derived growth factor (PDGF), stem cell factor (SCF), transforming growth factor-β (TGF-β), tumor necrosis factor-α (TNF-α), and vascular endothelial growth factor (VEGF). The actions of these mediators are summarized in Table 1.
Laser irradiation is characterized by a number of physical parameters such as wavelength, spot size, power, power density, energy, energy density, duration of irradiation, various pulse regimes, number of irradiations, and interval between consecutive irradiations; however, at present, the relevance of these parameters to the putative healing effects of laser irradiation on different injuries and skin conditions remains unclear. Variability in results from several previous investigations may be in part a consequence of the wide range of suitable parameters for laser irradiation and even the inability to measure the possible effects after irradiation with the necessary objectivity. 27,28 The potential effects of variations in the number of irradiations and dosing intervals have been rarely considered. Similarly, the possibility that the photobiomodulatory effects of laser irradiation are dissimilar across different cell types and species has been largely ignored. 29 The current review was informed by the recommendations of Tuner and Hode, 30 who have previously identified important factors to be taken into consideration when examining such studies.
The aim of this paper was therefore to review experimental studies of laser irradiation of human and animal cells in culture to assess the possible photobiomodulatory effects of such irradiation in terms of the expression and release of growth factors and cytokines. The current review also included assessment of: The quality of the study designs and protocols used; The experimental protocols and laser treatments used, and the appropriateness of these to clinical applications in humans; The relevance of irradiation parameters to any observed effects.
Materials and Methods
Original research papers investigating the effects of laser therapy on human and animal cells in culture, published from January 2002 to September 2009, were retrieved and used for this review. Relevant papers were sought and obtained from library sources and the online databases PubMed and Medline using EndNote X1 (Thomson Reuters, Carlsbad, CA).
Search terms were “growth factor,” “cytokine,” “laser therapy,” “laser irradiation,” “laser phototherapy,” and “phototherapy.” Additional secondary sources of information included reference lists from retrieved papers, and pertinent papers identified by hand searches of relevant journals not found in the databases (e.g., Laser Therapy).
We included studies that met the following criteria: Laser (or other monochromatic light source) was investigated as the primary intervention (independent variable) The type of laser and precise wavelength were defined or implied At least one outcome or index of cellular expression or release of growth factors or cytokines was identified as the dependent variable Studies were performed with human and/or animal cells or cell lines in culture Studies were related to wound repair or soft tissue regeneration
Studies excluded from this review were: Articles published prior to 2002
In vivo studies involving whole animals rather than isolated cells or cell lines Studies reported in languages for which no English language translation was available Studies performed with cells from animals other than common laboratory animals Studies involving cells or tumour cells isolated from hard tissues (e.g., osteoblasts, osteoclasts, or osteosarcoma cells) Reviews and meta-analyses Studies involving multisource or multiwavelength arrays, photodynamic therapy, high power laser, or flashlamp pulsed-dye laser Studies for which only an abstract was available
EndNote searches were carried out independently by two of the authors (PP, BR). Then, the articles for inclusion or exclusion were identified independently and confirmed, thereby minimizing bias. For included articles, the following data were extracted and tabulated by two of the authors (PP, BR):
Research method (including controls; minimizing variability in experimental conditions due to scattering of laser light and a reduction in energy when laser light is transmitted through culture medium; culturing of cells to subconfluent monolayer and ensuring that the whole of the cell monolayer was irradiated; and measurement of power density and energy density at the level of the cell monolayer);
Species type (human or animal cells, source, and species if animal in origin);
Description of cells (normal, stressed [ = nutritional deficit], wounded, diabetic, or tumour cells; normal cells are ones which have been grown in the presence of an adequate concentration of serum (usually 10%) to support cellular proliferation and are not cells obtained from wounds or tumour cells.);
Laser treatment parameters;
Experimental outcomes;
Authors' conclusions (results of laser irradiation).
Studies were then critically reviewed in terms of study design, methodology (PP and TYC), and appropriateness of irradiation parameters (GDB). Comments on these study characteristics were also summarized and tabulated (as bold text within Tables 3 –10, and in the row headed: Comments on study design & findings).
Results
Searched studies were limited to those which investigated human or animal cells involved in wound or soft tissue repair or cell lines relating to soft tissues. They are summarized in Table 2. Cell types included: Normal cells Cells incubated in culture medium deficient in fetal bovine serum (FBS) (i.e., serum starved = nutritional deficit, with cells referred to as “stressed”) Cells incubated in culture medium to confluence and scratched with a needle (i.e., “wounded cells”) Cells incubated in culture medium to confluence, medium replaced with one containing a high concentration of glucose, and cells scratched with a needle (i.e., “diabetic wounded cells”) Cell lines, carcinoma
NF, not found in searched studies from January 2002 to September 2009.
Cell types included are those involved in wound and soft tissue repair cell lines, mostly carcinoma-based.
Results from the literature search are summarized in Fig. 1. In total, 17 publications 1,31 –46 were included in this review (n = 14, human; n = 3, animal) and are summarized in Tables 3 –10.
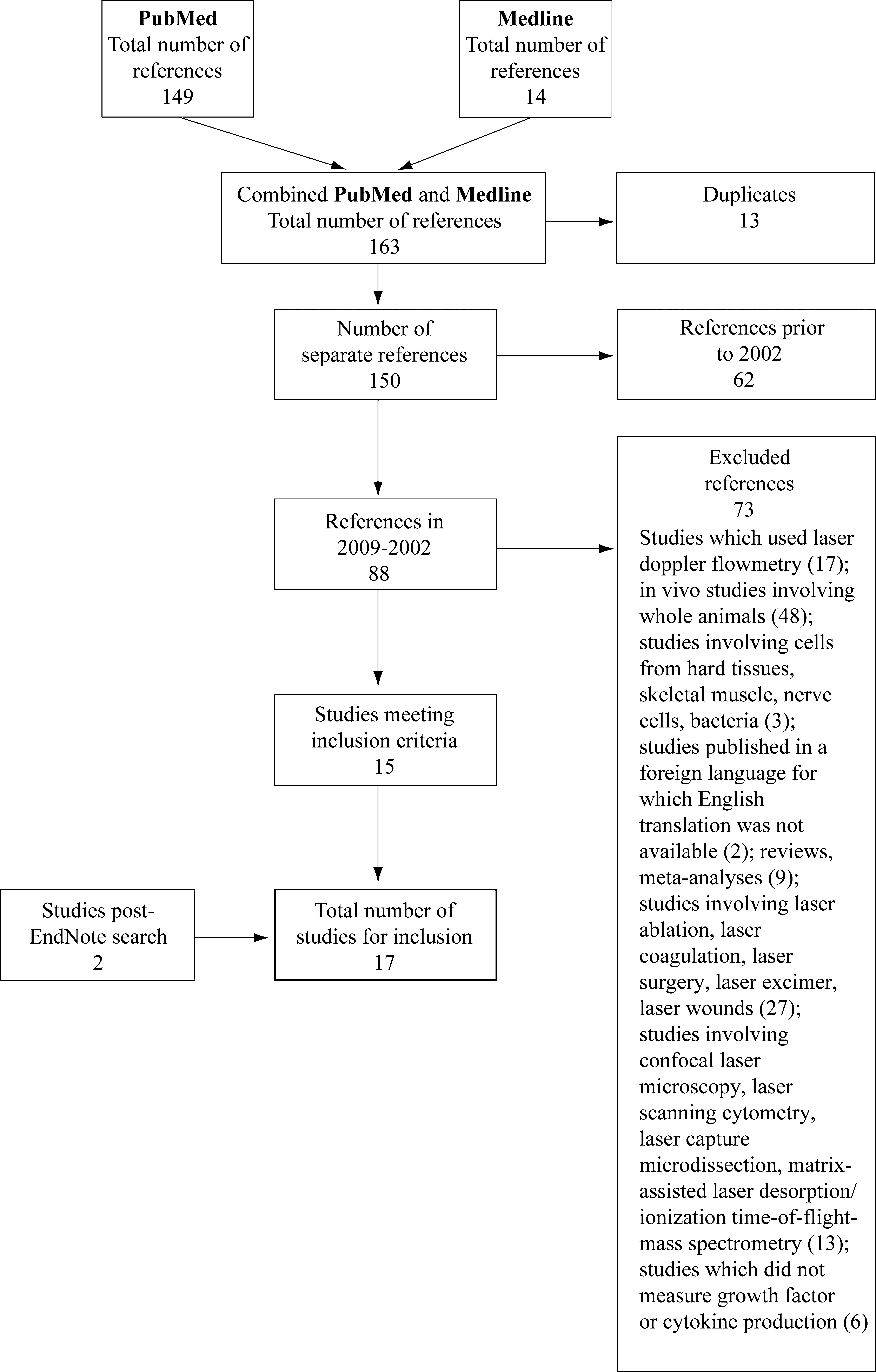
Flowchart of literature search method: studies with human and animal cells.
Fluence rate or flux, bfluence.
Fluence rate or flux; bfluence.
Fluence rate or flux; bfluence.
Fluence rate or flux; bfluence.
Fluence rate or flux; bfluence.
Fluence rate or flux; bfluence.
Fluence rate or flux; bfluence.
Fluence rate or flux; bfluence.
Outcomes reported in the retrieved studies included: Gene expression and production of pro-inflammatory and anti-inflammatory cytokines including IL-1α, IL-1β, IL-6, IL-8, IL-10, TGF-β, TNF-α, and IFN-γ Gene expression and production of growth factors including bFGF, IGF-1, SCF, HGF, KGF, and IGF
The following laser parameters were recorded from the searched studies: wavelength, power, power density (irradiance), energy, energy density (radiant exposure), spot area or irradiated area, method of irradiation, number of irradiations, duration of irradiation, interval between irradiations, and time after final irradiation for outcome measurements. These parameters are summarized in Tables 3 –10.
Summaries of major findings
Application of laser light to cells in the reviewed studies
There was considerable variation in the method of laser irradiation in the reviewed studies. This is consistent with the findings of our previous review
47
and included the following issues: Irradiation of cells from above or below with a constant distance between the laser beam and cell monolayer in dishes, wells, or plates Sheltering of adjacent wells from scattered laser light, for example by using a foil lid Minimizing reflection of laser light by using black backgrounds for irradiated areas Minimizing absorption of laser light when irradiating cells from above by replacing colored culture medium with phosphate buffered saline, PBS
In some cases, the laser beam diameter was adjusted so that the entire area of the dish, plate, or well was irradiated. This was achieved using an optical lens system to expand or clip the beam. Irradiation of cells from above was often performed using an optical fiber, while in other studies irradiation from below was carried out using a contact mode, with the tip of the laser diode positioned against the bottom of the dish or plate. In many studies, laser irradiation was carried out in the dark or partial darkness to minimize the effect of ambient light. Phenol red indicator was often omitted from the culture medium on account of it having an estrogen-like effect. 48,49
Irradiation parameters
Irradiation parameters reported in the reviewed papers varied widely, and in some cases there was incomplete reporting of relevant parameters: five of the sixteen studies (5/16) did not provide sufficient information to allow calculation of missing parameters (see Tables 3 –10). Such variation in parameters (or lack of specification of key details) limits comparison ability between studies.
(i) Wavelength (nm)
Wavelength was reported in all of the reviewed studies. While wavelengths in the visible red range (632.8–685 nm) were most commonly investigated (12/16; either in isolation or in comparison with other wavelengths), wavelengths ranged from 532 nm (green) to 1064 nm (infrared). The predominance of the use of red wavelengths is in keeping with current clinical practice for laser applications in wound healing and repair. While reported findings were, regardless of wavelength employed, uniformly positive with respect to photobiomodulatory effects, several studies which compared wavelengths in human fibroblasts reported wavelength-dependent differences in cellular responses (see Tables 4 and 5). 33,35,38,40
(ii) Power (mW)
All but four (25%) of the reviewed studies reported the power output of the laser device employed. In the remaining studies, specified power output varied widely from 7 to 1000 mW (1 W), with the use of higher power outputs associated with infrared Nd:YAG and Ti-Sa laser systems. The potential relevance of this parameter to observed laser-mediated effects was not specifically investigated in any of the reviewed studies, nor is the relevance clear from the reported findings.
(iii) Irradiance (mW/cm2)
In low level laser therapy applications, irradiance (or ‘power density’) is specified in mW/cm2, and is derived from power (mW) and spot size or area of irradiation (cm2). Over one third of the reviewed studies (6/16) did not specify this parameter, and three of these did not provide sufficient information to allow this parameter to be calculated. Where specified (or calculable), irradiance ranged from 2 to 1000 mW/cm2 (1 W/cm2); despite such variation, as for power (above), there appeared to be no clear relationship between irradiance and observed effects.
(iv) Energy (J)
Energy (specified in J) may be considered a basic form of dosage, and is derived from power (mW) and time of irradiation (s). Time of irradiation (where specified: 12/16) ranged widely from 3 to 7,729 s (i.e., over 2 h); such variation reflects in part the range of energies or dosages used, and the reciprocity with laser power outputs (i.e., longer times are required with lower output powers to achieve a specified dosage). Energy as a parameter was not specified in any of the review studies, nor was its relevance specifically investigated (e.g. 40 ; however, cf. radiant exposure, below), but where this could be derived from other specified parameters (10/16), its value ranged widely, from 0.01 to 1260 J. This is probably indicative of its being less useful as a viable parameter.
(v) Radiant Exposure (J/cm2)
Radiant exposure (J/cm2) is generally considered and reported as the most appropriate means of specifying dosage, at least in the case of experimental studies. It is derived from time (s) and irradiance (mW/cm2), which is in turn derived from power (mW) and area of irradiation (cm2). Radiant exposures were reported in all papers, and ranged from 0.1 to 16 J/cm2 (see Tables 3 –10); this represents an interesting contrast with other parameters, and energy in particular, for which values ranged by factors of up to 1,000.
This was the parameter which was most commonly investigated in studies: i.e., to assess the ‘dosage-dependency’ of observed effects. Such studies consistently found evidence of dose-dependent effects, 32,35,40 –42,45 including dosage thresholds for mediated effects, 41 and possible reversal of effects at higher dosages (as predicted by the Arndt-Schultz Law). 40
(vi) Irradiation Regime
Considerable variation was found in the duration of irradiation, the number of irradiations, and the interval between successive irradiations. The number of irradiations varied between one and six doses – in some studies two or three doses were given on the same day – and the time interval between successive irradiations varied between 6 and 72 h. Some studies reported differences in responses with repeated doses, providing evidence of cumulative effects of laser irradiation. 1,34,39
Measurements of outcomes
Time after Final Irradiation
The times at which outcomes were measured following irradiation (or the final irradiation if several irradiations were applied) varied from 15 min to 120 h after irradiation (or the final irradiation if multiple irradiations took place).
Synthesis and Release of Growth Factors and Cytokines
Data from the searched studies were critically analyzed to determine whether specific cell types of human and animal origin were influenced in a similar way by laser light of a particular wavelength, and with a similar power density and energy density. Unless stated otherwise, cells were irradiated once. Responses of irradiated cells were compared with those of non-irradiated cells (controls).
a) Stem cells (Table 8)
No human studies were found in the current search.
Laser irradiation of rat bone marrow mesenchymal cells at 635 nm with 5 J/cm2 significantly stimulated the secretion of VEGF and NGF at 24 h after irradiation.
b) Stressed smooth muscle cells (Table 9)
No human studies were found in the current search. Pig aortic smooth muscle cells irradiated at 780 nm with 2 J/cm2 and cultured in nutritional deficient medium had a decreased gene expression of IL-1β within the first 30 min after irradiation.
c) Stressed keratinocytes (Tables 3 and 7)
For human keratinocytes (HaCaT cell line) irradiated at 780 nm with 2 J/cm2, gene expression of IL-1α peaked at 2 h after irradiation. IL-6 expression was activated immediately after the end of irradiation, then peaked at 2 h, and was repressed at 6 h; control levels of IL-6 reappeared at 10 h. KGF was upregulated at 2 h.
Pro-inflammatory IL-1β gene expression was inhibited in a gradual fashion by irradiation. There was a sevenfold increase in upregulation of expression of IL-6 in sublethally irradiated keratinocytes using 532 nm at a dose of 1.6 J/cm2. Irradiation of human foreskin keratinocytes at 632.8 nm with 0.5, 1.0, and 1.5 J/cm2 showed a dose-response effect for the total amount of bFGF released by the cells into the culture medium (including during irradiation and 30 min after irradiation), with the greatest stimulation occurring with 1.5 J/cm2. The total amount of NGF released by the cells into the culture medium at 24 h also showed a dose-response effect with the greatest stimulation occurring with 1.5 J/cm2. Laser treatment showed no significant stimulatory effects on SCF, HGF, and ET-1 release by the cells. No animal studies were found in the current search.
d) Stressed monocyte/macrophages (Table 10)
No human studies were found in the current search.
Mouse macrophages (RAW 264.7 cell line) activated by the addition of lipopolysaccharide (LPS) (0, 0.1, and 1 μg/ml) to the culture medium immediately before irradiation at 780 nm with 2.2 J/cm2 had significantly reduced gene expression of MCP-1, IL-1α, IL-1β, IL-6, and IL-10 at 2 h when stimulated by 1 μg/ml LPS. Irradiation did not influence TNF-α gene expression; irradiation also did not influence gene expression when cells were not stimulated with LPS or when they were stimulated with 0.1 μg/ml LPS. Irradiation reduced LPS-induced secretion of MCP-1 over non-irradiated cells at 12 h.
e) Fibroblasts (Table 4)
Human gingival fibroblasts irradiated at 660 or 780 nm with 3 or 5 J/cm2 did not alter KGF production, but that of bFGF was significantly increased in groups irradiated at 780 nm. Irradiating human gingival fibroblasts once at 685 nm with 2 J/cm2 caused a significant increase in bFGF and IGF-1 release, but not of insulin growth factor binding protein-3 (IGFBP3). Irradiating the cells twice with a 24 h interval gave significant increases in all of the measured parameters.
No animal studies were found in the current search.
f) Stressed fibroblasts (Tables 4 and 7)
For human foreskin fibroblasts irradiated at 632.8 nm with 0.5, 1.0, and 1.5 J/cm2, a dose-response effect was found for the total amount of bFGF released by the cells into the culture medium (including during irradiation and 30 min after irradiation), with the greatest stimulation occurring with 1.5 J/cm2. Laser treatment showed no stimulatory effect on NGF release by stressed fibroblasts and thereby differed from findings with irradiated stressed keratinocytes (see above). Laser treatment showed no significant stimulatory effects on SCF, HGF, and ET-1 release by stressed fibroblasts. Irradiation of human foreskin or skin fibroblasts with Q-switched laser, 532 nm, at a sublethal dose of 0.8 J/cm2 stimulated SCF, HGF, and bFGF gene expression in five out of six cell lineages. A second study showed upregulation of expression of these growth factors in sublethally irradiated fibroblasts using the 532 nm laser at a dose of 0.8 J/cm2. No animal studies were found in the current search.
g) Wounded fibroblasts (Table 5)
Wounded human skin fibroblasts exposed to 5 or 16 J/cm2 (one irradiation on day 1 and again on day 4) using 632.8, 830, or 1064 nm showed an increase in release of IL-6 when compared with normal cells irradiated with the same dose and wavelength at 1 h after the final exposure. There were no significant differences between the different wavelengths, indicating that release of IL-6 was dependent on dose administered and not on wavelength. Wounded skin fibroblasts exposed to 16 J/cm2 using 632.8 nm and wounded fibroblasts exposed to 5 J/cm2 using 1064 nm showed an increase in release of IL-6 at 1 h after the final exposure (one irradiation on day 1 and again on day 4) compared with wounded non-irradiated cells. Also, wounded skin fibroblasts exposed to 16 J/cm2 using 1064 nm showed a decrease in bFGF release when compared with wounded non-irradiated cells (irradiated on day 1 and day 4). Interestingly, wounded skin fibroblasts exposed to a single irradiation of 16 J/cm2 at 1064 nm showed an increase in bFGF release when compared with wounded, non-irradiated cells after 1 h. Wounded skin fibroblasts exposed to a single irradiation dose of 5 J/cm2 at 830 nm showed an increase in bFGF release after 24 h when compared with cells after 1 h. Wounded skin fibroblasts showed an increase in IL-6 release after a duration of 3 h when exposed to a single dose of 5 J/cm2 on one day, while an increase in IL-6 release was observed after a duration of 1 h when exposed to a single dose of 5 J/cm2 on two consecutive days. There was a significant increase in IL-6 release after cells were exposed to 5 J/cm2 on two consecutive days with a duration of 3 h when compared with cells exposed on one day with a duration of 3 h. This supports previous evidence that laser irradiation has a cumulative effect. No animal studies were found in the current search.
h) Diabetic wounded fibroblasts (Table 5)
Human skin fibroblasts grown in medium containing an additional 17 mM of D-glucose, wounded and irradiated with 5 J/cm2 at 632.8 or 830 nm showed a significant increase in bFGF release compared with diabetic wounded non-irradiated cells, as well as diabetic wounded cells irradiated with 16 J/cm2 at 632.8 or 830 nm. Diabetic wounded cells irradiated with 5 or 16 J/cm2 at 1064 nm showed no significant change in bFGF release, and thereby differed from irradiated wounded cells. No animal studies were found in the current search.
i) Stressed cell lines (Table 6)
Human melanoma cell line A2058 irradiated with 0.5, 1, or 2 J/cm2 at 632.8 nm had an increased release of IL-8 and TGF-β1 and was dose responsive, with 1 or 2 J/cm2 eliciting the greatest response. No animal studies were found in the current search.
Discussion
The most important finding from the current review is in relation to the effectiveness of laser therapy in modulating the release of growth factors and cytokines from a very wide range of different cell types, and the dependence of this on wavelength, power density, energy density, number of exposures, and interval between exposures. The current results strongly support the case for further controlled studies using human and animal cells including stem cells, cells grown under nutritional deficit conditions (“stressed” cells), cells grown in a medium supplemented with additional glucose (“diabetic” cells), fibroblast cells wounded in vitro (“wounded” cells), and fibroblast cells grown in a medium with additional glucose and wounded in vitro (“diabetic wounded” cells).
This review also highlights a number of important issues relating to the number of studies performed with human and animal cells, the quality of the study designs and protocols used, the types of cells used (particularly in terms of the appropriateness of these to clinical applications in human subjects), and the relevance of irradiation parameters.
Numbers of searched studies with human and animal cells
There was a wide disparity in the number of studies using human, as compared with animal, cells found in the search; a much greater number of studies had used human cells (n = 14) and very few employed animal cells (n = 3).
Research design and reporting of details
There was a lack of specification of key details in many of the reviewed studies (application of laser beam using an optical lens system or optical fiber, laser parameters such as wavelength and power density), and a wide variety of experimental methods were used. Parameter variability included irradiation of cell monolayers from above or below; differences in steps taken to minimize scattering and reflection of laser light, variable levels of influence of ambient light on cells, differences in absorption of laser light through the culture medium; variability in the range of irradiation parameters used, including the number and timing of irradiations, variation in interval between irradiations, and differences in time at which experimental outcomes were measured following laser irradiation. In most of the searched studies, the cells were irradiated from above.
Apart from inadequate reporting of important elements of research design, and errors in calculation of laser irradiation parameters, there were several other prevalent weaknesses in the reviewed studies which may have influenced some of the reported findings (e.g., incorporation in the culture medium of phenol red which has been shown to have an estrogen-like effect 48,49 ; lack of reported measurement of power density at the level of the cell monolayer; laser irradiation of cells not performed in the dark but under ambient lighting).
Types of cells used and influence of laser light
The cell types that have been used for measuring growth factor and cytokine gene expression and release are summarized in Tables 11 and 12. The methods used for determining gene expression involved reverse transcriptase-polymerase chain reaction (RT-PCR) for detection of mRNA expression levels, while release (or production) was determined by measuring the levels of these mediators in the culture medium or PBS. The main findings concerning the influence of laser light are given in Tables 11 and 12, which also include times of irradiation and energy densities.
As studies using both human and animal sources for specific cell types were not found, it was not possible to determine whether particular cell types of human and animal origin responded similarly to laser light. In addition, no studies were found on the release of growth factors and cytokines from human stem cells; the only study was for rat bone marrow-derived mesenchymal stem cells. 44
Low level laser irradiation was shown to modulate the gene expression and release of growth factors such as bFGF, TGF-β, KGF, and IGF-1, and the release of the pro-inflammatory cytokines IL-1β, TNF-α, and INF-γ as well as other cytokines such as IL-1α and IL-6. These findings are similar to those of in vivo studies in which, for example, rat gingival tissue was irradiated with 7.5 J/cm2 at 632.8 nm and shown to cause an inhibition of gene expression of IL-1β and INF-γ, while increasing that of PDGF (platelet-derived growth factor) and TGF-β. 50 IL-1β and TNF-α are expressed by activated monocytes, epithelial cells, and fibroblasts in inflammatory disorders. 51 It was also shown that laser light at 904 nm with 3 J/cm2 could activate TGF-β1 and -β3 complexes in cell-conditioned medium of human adenocarcinoma and fibrosarcoma cell lines. 42 It appears that low level laser irradiation is able to promote an increase in the gene expression and release of growth factors and cytokines that are anti-inflammatory and stimulate the healing process, and also to inhibit the expression of pro-inflammatory cytokines, thereby reducing inflammation.
Laser irradiation was found to have specific effects on various cell types. For example, low level laser treatment, continuous wave, with 0.5, 1.0, and 1.5 J/cm2 at 632.8 nm did not stimulate NGF release by stressed human fibroblasts, whereas NGF release was stimulated by irradiation of stressed human keratinocytes. 32 Also, there were no significant stimulatory effects on SCF, HGF, and ET-1 release by stressed human fibroblasts or stressed human keratinocytes. 32 NGF can act as a melanocyte survival factor. Sublethal laser irradiation of stressed human fibroblasts utilizing high-frequency pulsing of radiation with 0.8 J/cm2 at 532 nm stimulated bFGF, SCF, and HGF gene expression and production. 35,43 Using the same irradiation for stressed human keratinocytes, the gene expression of IL-6 was increased more than sevenfold while that of SCF and ET-1 was not significantly increased and HGF was undetectable. 43
Melanocytes are stimulated by the growth factors bFGF, ET-1, HGF, and SCF, and inhibited by other growth factors and cytokines including TNF-α, TGF-β1, IL-1α, IL-1β, and IL-6. 7 Thus, three melanogenic stimulating paracrine factors are released from fibroblasts, whereas keratinocytes release an inhibitor of melanogenesis (IL-6). The phenomenon of rebound hyperpigmentation following the use of the Nd:YAG laser has been thought to be due to direct melanocyte stimulation, but increased pigmentation through fibroblast stimulation could also be the primary event. 43
It is generally held that laser light stimulates cells that are growing slowly at the moment of irradiation, and that the proliferation of fast growing cells cannot be stimulated by laser irradiation. Thus, if cells are fully functional at the moment of irradiation, or are growing in a serum-rich environment (usually 10% FBS), there is limited potential for laser-mediated stimulation, and no therapeutic benefit will be observed. 52 Consequently, for cells to be capable of responding to laser light, they need to be grown in nutritionally deficient conditions (“stressed”) or in the presence of high concentrations of glucose (“diabetic”), both of which markedly decrease cell growth. Alternatively, they have been harvested from diabetic patients or animals. Another experimental approach is to use cells (e.g., fibroblasts) that have been grown to confluence and thereby have zero or minimal growth, and wound these cells by scratching with a sharp pointed instrument such as the tip of a glass pipette. When wounded or scratched, cell monolayers respond to the disruption of cell-cell contacts with an increased concentration of growth factors at the wound margin, and by healing the wound through a combination of cellular proliferation and migration. It was noteworthy that wounded skin fibroblasts exposed to a single irradiation of 16 J/cm2 at 1064 nm showed an increase in bFGF release when compared with wounded non-irradiated cells after 1 h; 37 by contrast, wounded skin fibroblasts exposed on day 1 and again on day 4–16 J/cm2 using 1064 nm showed a decrease in bFGF release when compared with wounded non-irradiated cells. 36
Times for measuring outcomes following laser irradiation of cells
It has been suggested from studies with human skin fibroblasts that a duration of between 1 and 3 h post-irradiation is sufficient when measuring the direct effects of laser irradiation on cells (cell viability, ATP mitochondrial activity, membrane integrity, and DNA damage), while a longer duration of 24 h is required to demonstrate indirect effects (cell proliferation and protein expression). 37,39 This notwithstanding, in many of the searched studies, growth factor and cytokine expression and release were measured in the period 30 min to 6 h following irradiation.
Mechanisms for laser-stimulated expression and release of growth factors and cytokines
Low-power laser irradiation increases the gene expression and release of several growth factors and cytokines. Before upregulation of KGF in keratinocytes 31 and release of TGF-β1 from melanoma cells, 41 an immediate increase was found in mitochondrial membrane potential level after laser irradiation. Accompanied by increased gene expression of these growth factors, the cells exhibited cell proliferation and differentiation induced by low-level laser irradiation. In addition, following irradiation of keratinocytes, the gene expression of IL-1α and IL-6 is transiently upregulated. 31 Irradiation of melanoma cell line A2058 cells induces an immediate increase in mitochondrial membrane potential, ATP, and cAMP via enhanced cytochrome c oxidase activity, and results in delayed effects on IL-8 release. 41 It is possible that these cytokines could promote proliferation of keratinocytes and melanoma cells, and migration of keratinocytes induced by laser light. These findings show a connection between the cytochrome c/mitochondrial membrane potential/ATP/cAMP/JNK/AP-1 pathway, cellular responses in terms of gene expression and secretion of growth factors and cytokines, and cell proliferation and migration. 53
Relevance of laser parameters to observed effects
A number of the studies assessed the relevance of specific parameters to observed effects of laser irradiation: these principally focused on wavelength-specific effects, dosage (radiant exposure) dependency, or a combination of these two. Notwithstanding the difficulties of direct comparison of results (given differences in experimental setup, and the plethora of possible combinations of laser parameters), the current findings provide consistent evidence of wavelength and dosage dependency for laser-mediated effects upon growth factor and cytokine release. Further controlled studies are warranted to investigate the putative relevance of other irradiation parameters (e.g., energy or pulsing frequencies), and whether such laboratory findings translate into clinical relevance.
In some studies, long irradiation times were required to reach the desired dose and could further stress already stressed cells or stress unstressed cells, possibly leading to altered expression or release of these mediators. For example, stressed human keratinocytes, stressed pig smooth muscle cells, and monocytes/macrophages from a stressed mouse cell line had been irradiated for 18 min. However, it cannot be determined from the data summarized in Tables 11 and 12 whether these cells had been further stressed as a result of this long irradiation time. Further studies would be required to investigate this.
Clinical relevance and further studies
The laser parameters found to maximize the release of anti-inflammatory cytokines and growth factors that promote cellular proliferation and migration essential for re-epithelization, angiogenesis, and granulation tissue formation could potentially be used to reduce inflammation and stimulate wound healing in the clinical environment. However caution is needed in extrapolating results in vitro to the clinical, in vivo situation. Melanin in the epidermis and blood in the superficial vasculature of the dermis have great affinity for absorbing laser light at shorter wavelengths (e.g., 532 nm), and thereby preventing any real penetration of light of this wavelength into the dermis, whereas laser light of longer wavelength (1064 nm) can penetrate deeper into the skin. 54,55 Thus, some of the results from experiments using cell types examined in vitro become less relevant at shorter wavelengths in vivo.
Further studies are needed to compare the effects of laser irradiation on the release of growth factors and cytokines by human and animal cells in culture, and also to study the effects of laser light on the proliferation and protein synthesis of stem cells, which are likely to become increasingly important as new treatment modalities for a wide range of human diseases and injuries.
Conclusions
Findings from this literature review consistently demonstrated the ability of laser light to photobiomodulate the release of growth factors and cytokines by human and animal cells in vitro. This was caused by laser light inducing changes in gene expression of some of these mediators, e.g., by increased gene expression of IL-1α and IL-6 at 2 h and inhibition of IL-1β by human keratinocytes following laser irradiation. 31 The stimulation of release of some mediators from irradiated cells, while the release of others was unaltered or inhibited, indicates specific effects brought about by laser light. These findings strongly support the case for further controlled research with such cells.
There was considerable variation in research design, methodology, and laser irradiation parameters which limited comparison of research findings between studies using the same cell type. No comparisons were possible for specific cell types between human and animal studies. Inadequate reporting of key details was also prevalent, as were some errors in specification or calculation of key irradiation parameters. These issues need to be considered in designing future research in this area.
Abbreviations
ATP, adenosine triphosphate; CW, continuous wave; DMEM, Dulbecco's modified Eagle medium; EDTA, ethylenediaminetetraacetic acid; EBSS, Eagle's balanced salt solution; EGF, epidermal growth factor; EMEM, Eagle's minimum essential medium; ET-1, endothelin-1; ELISA, enzyme-linked immunosorbent assay; FBS, fetal bovine serum; bFGF, basic fibroblast growth factor; HBSS, Hank's balanced salt solution; HGF, hepatocyte growth factor; IFN-γ, interferon-γ; IGF-1, insulin-like growth factor-1; IGFBP3, insulin-like growth factor binding protein 3; IL, interleukin; keratinocyte-SFM, keratinocyte serum-free medium; IR, infra-red; KGF, keratinocyte growth factor; LD50, lethal dose 50%; LDH, lactate dehydrogenase; LPS, lipopolysaccharide; MCP-1, monocyte chemotactic protein-1; MMP, metalloproteinase; MSH, melanocyte stimulating factor; NGF, nerve growth factor; NO, nitric oxide; iNOS, inducible nitric oxide synthase; PAR-2, protease-activated receptor-2; PBS, phosphate buffered saline; PDGF, platelet-derived growth factor; RT-PCR, reverse transcriptase-polymerase chain reaction; SCF, stem cell factor; TGF-β, transforming growth factor-β; TNF-α, tumor necrosis factor-α; VEGF, vascular endothelial growth factor.
Footnotes
Acknowledgments
Ms. Ryan was supported through the Centre for Physiotherapy Research, University of Otago, Dunedin, New Zealand. No financial support has been received in conjunction with the generation of this report. None of the authors had any conflicts of interest.
Author Disclosure Statement
No competing financial interests exist.