
Abstract
Introduction
Expression of staphylococcal virulence factors is regulated by several systems. An example is the agr regulon (accessory gene regulator). It controls genes expression via the quorum sensing mechanism, a specific bacterial system that gives information about cells density in a given environment. The Agr system is one of the best-known quorum sensing staphylococcal mechanisms, which plays an important role in the process of infection in a human organism. Studies indicate that the activity of this system correlates with the degree of the strains' virulence. 3 Although the agr locus is conserved among staphylococcus species, it bears a hypervariable, polymorphic fragment, which is used to classify strains into one of four different groups by the use of the PCR method. 4 The hypervariable region is situated in the fragment encoding signaling receptor AgrC, auto-inducing peptide AgrD, and protein AgrB, which are responsible for extracellular transport of the auto-inducing peptide. 5,6
One of the S. aureus mechanisms of resistance to methicillin is based on production of penicillin-binding protein (PBP2a) encoded by mecA gene. 7,8 The mecA gene is situated on the mobile genetic element, integrated into bacterial chromosomes and called the Staphylococcal Cassette Chromosome mec (SCCmec). 7 This element can be transferred between bacterial cells by means of horizontal transfer. 9 Apart from the mecA gene, SCC also harbor regulatory genes, insertion sequences, and genes encoding ccr recombinases that are responsible for cleavage and integration of SCCmec into the bacterial chromosome. 9 Mec and ccr complexes were used to create four classes and these classes allowed eight different types of SCCmec to be defined. 10 Those types, listed as SCCmec I–SCCmec VIII, differ in their size and genetic element content. As the SCCmec types VI, VII, and VIII are rarely isolated, epidemiological studies mainly analyze types I–V. 8,11 –15
The search for gene polymorphisms and determination of genetic profiles of particular strains can have an important diagnostic value. Previously, scientists have shown that the differences in expression of various bacterial virulence factors connected with the agr group correlate with varied pathogenicities and progression of disease. In the case of SCCmec, the type may give information about the strain's origin, as a majority of nosocomial strains belong to SCCmec I, II, or III and, in most cases, CA MRSA strains (Community Acquired MRSA) carries SCCmec IV. 11,13 It was also shown that virulence factors associated with SCCmec types play a role in the progression of bacteremia caused by S. aureus. Published data show that strains bearing SCCmec II are the cause of highest morbidity among patients with staphylococcal infections. 16
Easily developing multidrug resistance of S. aureus strains shows a great need for new, alternative therapeutic methods. 7 One of those methods may be photodynamic inactivation (PDI). It is based on the action of compounds called photosensitizers (PS), which selectively accumulate in certain tissues or cells and after illumination with specific light source and can generate cytotoxic reactive oxygen species (ROS). 17 Some research shows that the response of S. aureus to PDI varies among strains. The reduction of colony units may range from 0 to 5 log10, and therefore, it is possible to state that some strains are less sensitive to photoinactivation. 18 The mechanisms responsible for such discrepancies in response to PDI are still unknown, even though the correlation between biofilm production and response of S. aureus strains to PDI was previously determined; and it has also been shown that MRSA strains are more resistant to this method than MSSA strains. 19,20 Therefore, studying the genetic profile and seeking its correlation with the response to PDI seems to be of significant value. The genetic profile would not only enable further research focused on responses to PDI, but would also provide useful diagnostic information about effective therapy.
Materials and Methods
Study design
1. Identification of the species. All 99 bacterial isolates were identified as S. aureus species.
2. Photodynamic inactivation. Each strain (99 isolates) was cultured and incubated with defined dose of photosensitizer (protoporphyrin diarginate) and next irradiated with a red light at a dose of 12 J/cm2. After an 18 h incubation, the Colony Forming Units were counted and the results were analyzed statistically.
3. Determination of the agr group. The agr polymorphism was determined for all 99 S. aureus strains with the use of the Multiplex PCR reaction.
4. Determination of the type of SCCmec element. The determination of the type of SCCmec element was carried out for all MRSA strains with the use of Multiplex PCR reaction.
5. Looking for any statistically relevant correlation between genetic profiles of studied S. aureus strains and their response to photoinactivation.
Bacterial isolates
A total of 99 clinical S. aureus strains (46 MRSA and 53 MSSA) isolated in the Provincial Hospital in Gdansk were used. The isolates were characterized by Gram-staining and ability to produce coagulase and clumping factor using Slidex Staph Plus (BioMerieux, Marcy l'Etoile, France). Additionally, the species was identified using the biochemical identification system ID 32 Staph (BioMerieux) and confirmed by PCR-based detection of the nuc gene characteristic for S. aureus species. 21 Resistance to methicillin was determined using the disc-diffusion method, as well as by using the latex test detecting PBP2a protein (Staphytect Plus, Oxoid, Lenexa, KS). Among these 99 strains, 57.5% were isolated from the patients hospitalized in the Orthopedic Ward, 16.25% in the Surgery Ward, 12.5% in the Intensive Care Unit (ICU), 7.5% in the Dermatologic Ward, 3.75% in the Pediatric Ward, 1.25% in the Internal Medicine Ward, and 1.25% in the Laryngological Ward. 6.25% (5 isolates) of strains were isolated from surveillance culture, 77.5% (62 isolates) from patients with local infections, 8.75% (7 isolates) from those with bacteremia and generalized infections, and 7.5% (6 isolates) from infections connected with endoprosthesis. The Local Ethics Committee accepted the protocol of this study.
PCR amplification of the staphylococcal nuc gene
To amplify the thermostable nuclease gene (nuc) fragment, the primers listed in Table 1 were used. Each amplification was performed in a final volume of 20 μl that consisted of 200 μM of each dNTP, 20 pmol of each primer, and 1 unit of Taq DNA polymerase (Fermentas, Lithuania). The applied thermal program was as follows: initial denaturation at 94°C for 4 min, followed by 36 cycles of 1 min at 94°C, 0.5 min at 55°C, and 1.5 min at 72°C with a final step at 72°C for 3.5 min. The PCR product was analyzed by electrophoresis on 1% agarose gels. 22
PDI studies
All bacterial isolates were submitted to photodynamic inactivation (PDI). The bacterial culture was grown overnight (16 h) at 37°C in nutrient trypcase soy broth (BioMerieux) and then diluted with fresh broth to an appropriate density (OD600 = 0.05) (107/ml bacterial cells). Such S. aureus culture (stationary phase) was incubated with protoporphyrin diarginate in the dark at 37°C for 15 min at concentration of 25 μM. Afterwards, the cells were transferred into a 96-well microtiter plate (100 μl per well) and illuminated with appropriate light at room temperature for 60 min (12 J/cm2). After the illumination, aliquots of 10 μl were taken to determine the colony forming units (CFU). The contents of the wells were mixed before sampling. The aliquots were serially diluted 10-fold in PBS to give dilutions of 10−1 to 10−3 times the original concentrations and were streaked horizontally on square trypcase soy agar plates. Plates were incubated at 37°C overnight. As a control, the second microtiter plate was prepared in similar way. Overnight culture (16 h) at 37°C in nutrient trypcase soy broth (BioMerieux) was diluted with fresh broth to an appropriate density (OD600 = 0.05) (107/ml bacterial cells). Such S. aureus culture was incubated with protoporphyrin diarginate in the dark at 37°C for 15 min at concentration of 25 μM. Next, the cells were transferred into a 96-well microtiter plate (100 μl per well) and incubated in darkness at room temperature for 60 min. Thus, controls consisted of bacteria untreated with photosensitizers or light but kept in 96-well plates for the duration of the illumination, and bacteria exposed to light without photosensitizers. After the illumination time, the aliquots (10 μl) were taken from each well to determine the colony-forming units. The aliquots were serially diluted 10-fold in PBS (0.13 mmol/l NaCl, 8.1 mmol/l Na2HPO4, 2.68 mmol/l KCl, 1.47 mmol/l KH2PO4) to give dilutions of 10−1, 10−2, and 10−3 times the original concentrations. The aliquots (10 μl) of each of the dilutions were plated on trypcase soy agar (BioMerieux). After 18 h of incubation at 37°C in the darkness, the colony forming units were counted and results were statistically analyzed. Each experiment was performed three times. Survival fractions were expressed as ratios of CFU of bacteria treated with light and photosensitizer to CFU of untreated bacteria.
Photosensitizer
Protoporphyrin diarginate (PPArg2) (λ max 624 nm) delivered by the Institute of Optoelectronics, Military University of Technology (Warsaw, Poland) was used as a sensitizer. PPArg2 was dissolved in distilled water to give a 10 mM stock solution and stored at a −20°C in darkness until use. In Poland the group of Prof. Graczyk sensitized a new class of protoporphyrin IX amino acid derivatives which, due to the attachment of two arginines, became well soluble in aquatic solutions and possess amphiphilic character. Our laboratory has recently demonstrated a high effectiveness of PPArg2-based photoinactivation against multiresistant S. aureus strains (MRSA). 18
Light source
The illumination was carried out using BioStimul Lamp (Biotherapy, Czech Republic). Delivered light energy was determined with the use of a light power meter (model LM1, Carl Zeiss, Germany) and was approximately 0.2 J/cm2 per minute. The BioStimul Lamp emits polarized (96% level of polarization) and monochromatic light (624 ± 18 nm).
Multiplex PCR detection of agr gene group
All bacterial isolates were submitted to determination of the agr gene group. DNA extraction was performed with the usage of the commercially available DNA extracting kit (A&A Biotechnology, Poland), followed by PCR agr gene amplification. The specific primers listed in Table 1 were used. Each amplification was performed in a final volume of 25 μl that consisted of 200 μM of each dNTP, 30 pmol of each primer, and 1 unit of DyNAzyme™II DNA polymerase (Finnzymes, Finland). The thermal step program was as follows: initial denaturation at 94°C for 1 min, followed by 36 cycles of 0.5 min at 94°C, 0.5 min at 55°C, and 1 min at 72°C with a final step at 72°C for 10 min. 23 The PCR product was analyzed by electrophoresis on 1% agarose gels. 23
Multiplex PCR detection of SCCmec type
SCCmec type was determined for all MRSA strains from the studied collection. The specific primers listed in Table 1 were used. Each amplification was performed in a final volume of 35 μl consisting of 400 μM of each dNTP, 30 pmol of each primer, and 0.5 units of Marathon DNA polymerase (A&A Biotechnology). The applied thermal program was as followed: initial denaturation at 95°C for 2 min, followed by 30 cycles of 0.5 min at 95°C, 0.5 min at 58°C, and 1.5 min at 72°C with a final step at 72°C for 4 min. The PCR product was analyzed by electrophoresis on 2% agarose gels. 24
Results
Strain-dependent response of S. aureus strains to photoinactivation
We have determined the response of analyzed S. aureus strains to photodynamic inactivation and observed that the response of the analyzed population to PDI was highly differentiated and ranged from high susceptibility to elevated resistance. Based on calculated log10-unit reduction, we could classify the analyzed strain to one of four groups that reflected the type of response to photodynamic inactivation (Table 2). This classification was previously determined by Grinholc et al. 18
Resistant; bIntermediate resistant; cSensitive; dHighly sensitive.
Agr group determination
Within the agr locus we can specify the hypervariable region. Within this region, the particular polymorphism is used to determine one of four groups of S. aureus strains. To determine the group of the agr gene within S. aureus strains isolated from patients hospitalized in Gdansk, Poland, we have implied Multiplex PCR method. Percentage distribution of particular groups in analyzed population was as follows: agr 1, 59%, agr 2, 15%, agr 3, 25%, and agr 4, 1%. We have observed that in each PDI group, the most prevalent strains were those with agr type 1 (Fig. 1).
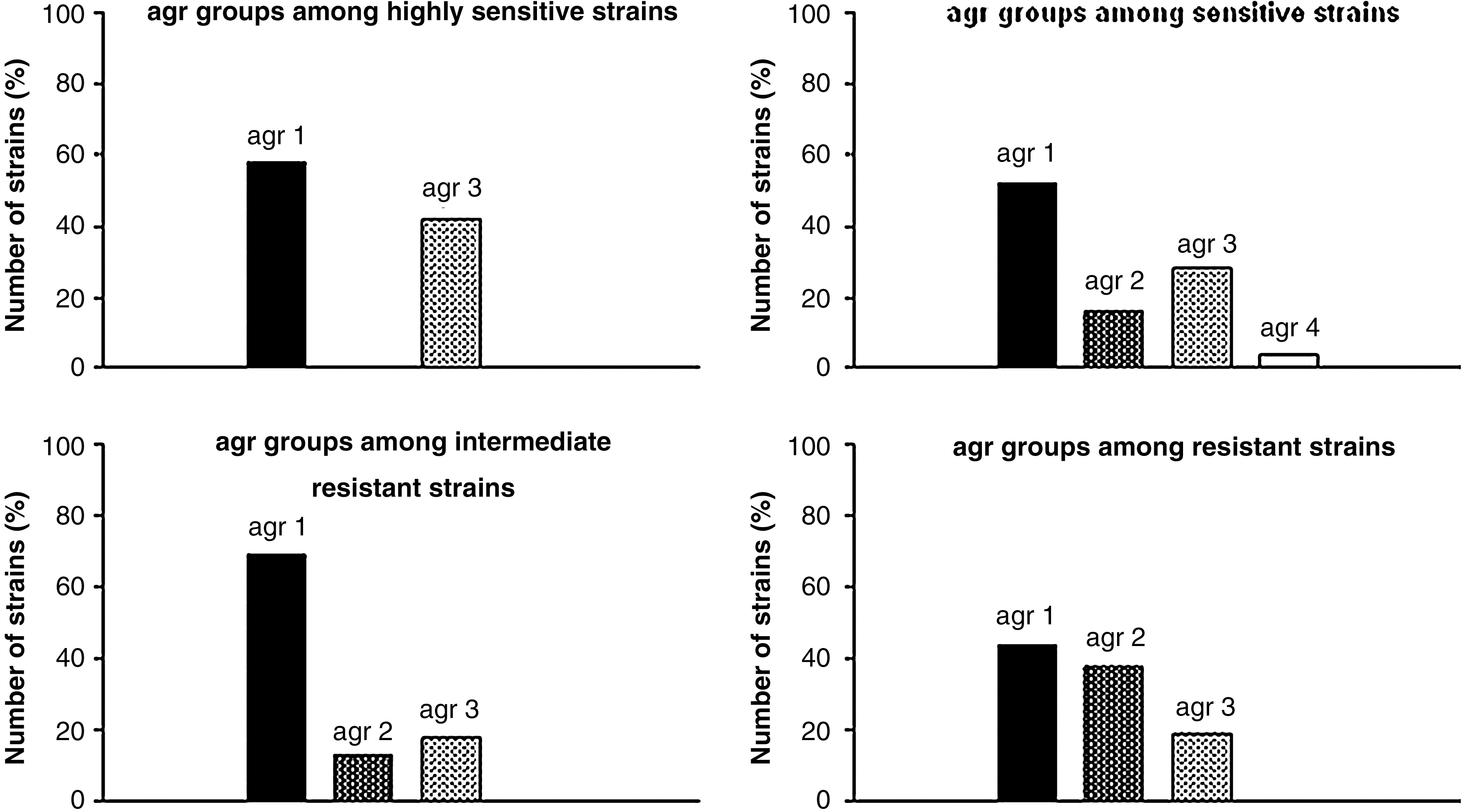
Percentage of strains with certain agr group and type of response of those strains to photodynamic inactivation.
SCCmec group determination
Staphylococcal Cassette Chromosome mec (SCCmec) is a mobile genetic element integrated into the bacterial chromosome. Genes present within this structure encode proteins responsible for methicillin resistance of MRSA strains. Based on different combinations of genes present in the SCCmec region, it is possible to determine eight groups that differ in the size and presence of genetic elements. In epidemiological research, the first five are usually present while the last three are rarely found. In this work, we used the Multiplex PCR method to analyze SCCmec groups within 46 MRSA strains isolated from patients hospitalized in Gdansk. Strains with SCCmec type I accounted for 40% of the analyzed population, type II: 13%, type III: 19%, type IV: 15%, and type V: 13%. As the distribution of all five SCCmec types in each PDI group (among MRSA strains) is quite even, it is not possible to state that a correlation between any type of response to PDI and SCCmec group type is present (Fig. 2). Nevertheless, we have observed a few relations between analyzed factors. Obtained data show that in analyzed population, the strains with SCCmec type IV were not susceptible to photodynamic inactivation. This information suggests that the presence of SCCmec type IV could be related to PDI resistance. Additionally, most of the analyzed SCCmec type V strains showed high susceptibility to photodynamic inactivation. Moreover, within the studied population most of the strains with intermediate resistance to PDI (57%) were characterized with SCCmec type I.

Percentage of methicillin-resistant S. aureus strains (MRSA) with certain SCCmec group and type of response of those strains to photodynamic inactivation.
Determination of genetic profiles containing agr and SCCmec types
The next step was to determine the genetic profile of analyzed strains. This profile was comprised of agr and SCCmec type, which together provided a genetic pattern. Based on these groups, we could determine 11 different patterns within 46 analyzed MRSA strains, each pattern comprising of one of four agr groups and one of five SCCmec groups (Table 3).
agr/SCCmec determined genetic profile vs. PDI effectiveness
Obtained data show that the distribution of agr/SCCmec patterns among susceptible to as well as resistant to photodynamic inactivation strains is quite even and therefore it is not possible to determine one pattern characteristic to specific type of response to PDI (Table 3). Most of the strains with intermediate resistance to PDI strains (66.6%) could be described as pattern I (agr1/SCCmecI). However, this pattern is not strictly specific to such a PDI response. Thus, no conclusion stating the predictive value of this pattern could be made. Nevertheless, the presence of pattern IV (agr1/SCCmecIV) could suggest elevated resistance of the strain to photoinactivation, as all strains characterized with this profile were intermediate or highly resistant to photodynamic inactivation (Table 3).
Moreover, all strains that were highly susceptible to PDI could be characterized with one of three patterns (I, III, or V) and all of these patterns consisted of agr group 1 (Table 3). Considering together the agr and SCCmec elements, reveals the fact that among highly sensitive strains there is no strain with agr group different than agr 1. Because the number of S. aureus samples with agr groups 2 and 3 is not numerous enough, we could not draw statistically significant conclusions, however, such an observation is worth being underlined. On this basis, one could conclude that strains described with a pattern different than agr 1 reveal elevated resistance to photodynamic therapy.
Discussion
The emergence of CA-MRSA (community acquired MRSA) and VRSA strains (vancomycin-resistant Staphylococcus aureus) has become a serious therapeutic problem, thus studying mechanisms responsible for S. aureus pathogenicity could play a key role in contemporary medicine. 25 Moreover, the development of alternative therapies and diagnostics is required.
One of the mechanisms responsible for S. aureus pathogenicity is encoded by locus agr regulating the expression of staphylococcal virulence factors. Agr proteins form a two-component transduction system, based on the quorum sensing mechanism and biofilm formation. 26 –28 The next element of S. aureus pathogenicity is its multidrug resistance. The gene responsible for resistance to β-lactam antibiotics is the mecA gene situated on the mobile genetic element called Staphylococcal Cassette Chromocome mec (SCCmec). 29
The genetic polymorphism and presence of specific gene's groups could be a marker of great diagnostic value. It was reported that differences in expression of virulence factors depend on specific agr groups and correlates with different pathogenicity and model of disease progression. Guangyong et al. showed that most of S. aureus strains causing toxic shock syndrome belong to agr group III. 6 Results of Jarraud et al. show that strains bearing agr IV are often a factor of staphylococcal scalded skin syndrome, and strains characterized with the presence of either agr I or agr II cause endocarditis. 5,30 SCCmec group analysis can provide information concerning the origin of an MRSA strain as this cassette is used as the epidemiological marker. 11,13 It was shown that virulence factors connected with SCCmec play a key role in bacteremia progression. Moreover, strains bearing the SCCmec type II are characterized with the highest mortality among infected patients. 16 The above results suggest that the genetic profile of a strain could be a useful tool for pathogenesis analysis and infection control.
Photodynamic inactivation can be an effective and alternative therapeutic method. However, it is worth noticing that some strains reveal elevated resistance to photoinactivation. Our current results are in compliance with those published previously and show that the response of S. aureus strains to PDI (employing PPArg2 as the photosensitizer) differs and ranges from high susceptibility to elevated resistance. 18 Moreover, out of 99 analyzed strains, 55% showed intermediate or high resistance to PDI. Thus, photodynamic therapy with the use of this particular photosensitizer and light wavelength cannot be considered a 100% effective method for S. aureus infections treatment. Our results indicate that there is a strain-dependent mechanism of resistance to PDI and how to determine this resistance.
The agr locus is one of the most important sets of genes controlling expression of staphylococcal virulence factors. Differences present in the agr hypervariable region can be associated with different onset of infection and response to treatment. 30 Thus, the influence of the agr group on PDI effectiveness could be observed. Our obtained distribution of the agr profile, in particular PDI groups, resembles the dispersion of one of the strains in the entire analyzed population. It proves a lack of correlation between the agr group and the strain's response to PDI. The only exception could be observed within the group of strains highly susceptible to photodynamic inactivation. This group consisted only of strains with agr 1 and agr 3. Moreover, there were no agr 2 isolates highly susceptible to PDI, even though 15% of the whole population was comprised of agr 2 S. aureus strains.
Current results suggest the existence of some relation between SCCmec type and the response to PDI. As SCCmec classification is based on different combinations of mecA and ccr genes encoding PBP2a and ccr recombinases, respectively, it is hard to suggest the mechanism of its influence on photodynamic inactivation. It could be due to the polymorphic differences in SCC cassette's region J. This fragment is comprised of different genes and pseudogenes that could be connected to additional determinants of bacterial resistance to antibiotics different than β-lactams. 31 Our results show the connection between intermediate or high resistance to PDI and the presence of SCCmec type IV. This type of SCCmec element is the smallest structural version of all five analyzed types and it is thought to be the most mobile and the most variable. 32 Currently, eight different subtypes of SCCmec IV are described and the differences among them are observed within the J region. 33 Thus, our results could suggest that those differences within the J region can render S. aureus strains more resistant to PDI. However, further investigations must be performed to determine if the SCCmec element really influences the mechanism of S. aureus response to PDI.
Conclusion
Unfortunately, the mentioned-above relations are not specific enough to determine a diagnostically important pattern, which could enable predictions of strain response to PDI. Nevertheless, we can conclude that the connection between the response of S. aureus strains to photoinactivation and the strain-specific agr/SCCmec pattern could be observed.
Footnotes
Acknowledgments
This work was supported by Grant No. (N N405 165139) from the Ministry of Science and Higher Education (MNiSW), by Grant Mo. BW/B051-5-0289-9 from the University of Gdansk, by the grant for young investigators, The Foundation for the Development of Gdansk University, and by the European Union within the European Social Fund in the framework of the project “InnoDoktorant—Scholarships for PhD students, I edition.”
Author Disclosure Statement
No competing financial interests exist.