
Abstract
Introduction
Adequate muscle functioning is intimately correlated to functions of the mitochondrial energy production system. 19 Alterations in this muscular activity may be monitored by the activities of sucinate dehydrogenase (SDH) 20 and nicotinamide adenine dinucleotide diaphorase (NADH), 21 as these enzymes demonstrate mitochondrial early structural alterations. 22
The laser irradiation restores mitochondrial functions in vivo (0.33–1.34 J/cm2, 810 nm; GaAlAs), 23 increases mitochondrial NADH (660 nm; 0.6, 1.2, 2.4, and 4.8 J/cm2 and 632.8 nm; 460 J/m2), 24,25 and normalizes SDH activity in inflamed tissue. 26 In contrast, the irradiation (5 J/cm2; 400–500 nm) decreased SDH activity in fibroblasts. 27 Despite all controversy, there is still evidence of interaction among laser, cells, and tissues. Therefore, some studies suggest that the laser mechanism of action is to arrange the intermediary steps of electron flux 28 and to alter mitochondrial membrane potential, 29 or that light on photoreceptors is only a trigger for cell metabolism regeneration. 30 –32 However, light signal transduction pathways are still obscure.
The remodeling of skeletal muscles requires the activity of a family of enzymes known as the matrix metalloproteinase (MMPs). 33 These proteins are zinc metalloenzymes, with the following properties: (1) They are proteinases that degrade at least one component of the extracellular matrix (ECM); (2) They contain a zinc ion and are inhibited by chelating agents; (3) They are synthesized in a latent form that requires activation by proteolysis; and (4) They are inhibited by the tissue inhibitors of metalloproteinases. The members of the family are split into groups based on their structure and substrate specificity: collagenases, gelatinases, stromelysins, and the membrane-type MMPs (MT-MMPs). 34
MMP-2 and MMP-9 have been described in the current literature, considering their capacity in degrade the major components of the basal lamina and its presence in regenerating skeletal muscle and pathological states including muscular dystrophy. 33
Some studies reveal increased expression of MMP-9 after photodynamic therapy (635 nm, 37 J/cm2), 35 and in the association of LLLT (810 nm, 100 mW, 54 J) to orthodontic movements in rats. 36 The HeNe laser (632.8 nm; 0.54 J) reduced gingival inflammation and the MMP-8 levels. 37 The cells of the aorta smooth muscle irradiated in pigs with diode laser (1 and 2 J/cm2, 780 nm) increased activity of MMP-2 and expression of MMP-1. 38
Although the laser effectiveness is evident, further studies are needed to establish safe and effective conditions for this therapy, which favor comparison of biological effects. This study aims to evaluate the effect of LLLT on masseter muscle of Wistar rats by analysis of the activity of NADH, SDH, and MMP.
Methods
Animals
Thirty male Wistar rats (weighing 350 g) were randomly divided in six groups (n=5) receiving different density energy irradiations. Animals were raised in the Central Animal House of the Ribeirao Preto Campus of the University of Sao Paulo and kept in temperature controlled rooms (24–25°C), with dark–light cycles of 12 h. They were fed with ration and had water ad libitum. The study protocol was approved by the local Ethics Committee (Protocol no. 08.1.204.53.5) according to international laws.
Laser irradiation
Twin-Laser MmOptics (780 nm, IR, GaAlAs), belonging to the Electromyography Laboratory of the Department of Morphology, Stomatology and Physiology of the Faculty of Dentistry of Ribeirao Preto, University of Sao Paulo, were used.
Previously anesthetized animals (ketamine 75 mg/kg+xylazine 10 mg/kg) had the area of the left masseter shaved, and underwent to 10 laser irradiations with different energy densities: 0.5, 1, 2.5, 5, and 20 J/cm2 (Table 1), and other fixed parameters: 5 mW, 780 nm, spot area 0.04 cm2 (manufacturer information) (Table 2) on alternate days. The left masseter muscle was irradiated in two points. The control group (group I, 0 J/cm2) was submitted to the same stress as the experimental animals but, it was not irradiated.
Preparation of samples for NADH and SDH activity determinations
The animals were anesthetized (ketamine 75 mg/kg+xylazine 10 mg/kg) and decapitated for immediate dissection of the left masseter muscle 48 h after last irradiation. The muscle fragments were frozen in isopentane cooled by liquid nitrogen, and kept at −80°C until used. Transversal serial cuts (10 μm) were obtained in Leica cryostat microtome CM1850 and stained to demonstrate NADH and SDH, which represent the oxidative metabolism pattern of the muscle tissue. Thirty random microscopic fields were captured for each group (15 for NADH and 15 for SDH) at a magnification of 20 x, by using a Leica DM4000B photomicroscope (Germany) equipped with a Leica EC3 digital camera (Leica, Wetzlar, Germany).
To determine fiber type areas (light, intermediary, and dark), according to the point counting method (stereology), digital photomicrographs were used where an 80 points test-system was generated in ImageJ software (free from
All quantitative data are presented as mean±standard error. ANOVA was used for the statistical tests and the confidence level was 95% (p=0.05).
Qualitative analysis of proteinases by substrate embedded enzymography
Sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) substrate embedded enzymography (zymography) was used to detect enzymes with gelatinase activity. Assays were performed as previously reported. 39
Rat masseter muscle (1:4 w/w dilution) was homogenized in 50 mM Tris-HCl buffer (pH 7.4) containing 10 mM CaCl2 and 1% of protease inhibitor cocktail, in a Turrax TE-102 (Tecnal, Piracicaba, Brazil). Whole homogenates were centrifuged at 12,000 g for 10 min at 4°C. The Lowry method was used to measure protein levels in muscle homogenates. 40 Supernatant aliquots (50 μL) were mixed with 10 μL of 100 mM Tris-HCl buffer (pH 7.4) containing 4% SDS, 20% glycerol, and 0.001% bromophenol blue. Twenty microliters of the mixture (25 μg protein) were used for electrophoresis. Proteolytic activity was qualitatively analyzed by comparing controls and irradiated animals.
Results
Activity of NADH (Fig. 1, Table 3) revealed three types of fiber in the superficial bundle of rat masseter muscle: light, intermediary, and dark, according to the staining intensity. Area estimation of different fibers showed groups irradiated with the smaller density energies: 0 J/cm2 (group I), 0.5 J/cm2 (group II) and 1.0 J/cm2 (group III), having the highest areas of light fibers (low-oxidative capacity) (49.73±1.16, 51.27±1.13, and 52.93±0.90) and showing statistical differences (p<0.05) in relation to other groups. Groups receiving highest energy densities: 2.5 J/cm2 (group IV), 5 J/cm2 (group V), and 20 J/cm2 (group VI) (30±1.26, 33.47±2.15, and 31.67±1.78), showed increased areas of intermediary fibers (p<0.05), which were proportional to the decreased areas of the light fibers (Fig. 2). This difference indicates increased metabolic activity in the groups irradiated with highest energy densities, although the 20 J/cm2 group was not the one showing larger areas in intermediary fibers. In relation to the dark fiber areas, only group VI (20 J/cm2) had a significant increase (p<0.05) (2.27±0.47), but the area was very small.

Determination of NADH activity. Images (original magnification × 20) of serial sections (10 μm) of masseter muscle after irradiation with different energy densities. Light fibers with lower oxidative capacity (long arrow) and intermediary fibers (arrow) with greater oxidative capacity are indicated for the six groups.

Bar graph showing areas of muscle fiber types revealed by the NADH reaction in the six groups. Groups IV, V, and VI show an increase in areas of intermediary fibers corresponding to the decreased areas of light fibers, thereby revealing an increase in oxidative activity. No conclusion could be drawn from the very small areas of dark fibers.
SDH activity (Fig. 3, Table 4) also revealed, in the rat masseter muscle superficial fiber bundle, three types of fibers (light, intermediary, and dark) according to stain intensity. Estimated area of light fibers (low-oxidative capacity) was not statistically different among the six groups, although a tendency to lower areas was detected in the highest density energies such as that seen in group V (5 J/cm2) and group VI (20 J/cm2). Discrete differences were observed in the intermediate fiber areas in all groups. Therefore, groups I, II, III, IV, and VI were similar, but group V (5 J/cm2) revealed an area slightly larger (p<0.05) (Fig. 4). A tendency to increased metabolic activity was detected in the three groups receiving the highest energy densities; group IV showed an area in intermediary fibers of 29.73±1.25; group VI showed an area of 30.20±1.72; and group V showed an area of 32.20±1.61. Dark fiber areas did not show a metabolic pattern related to energy densities irradiated with the laser, not revealing statistical differences or tendencies.

Determination of SDH activity. Images (original magnification ×20) of serial sections (10 μm) of masseter muscle after irradiation with different energy densities. Light fibers with lower oxidative capacity (long arrow) and intermediary fibers (arrow) with greater oxidative capacity are indicated for the six groups.
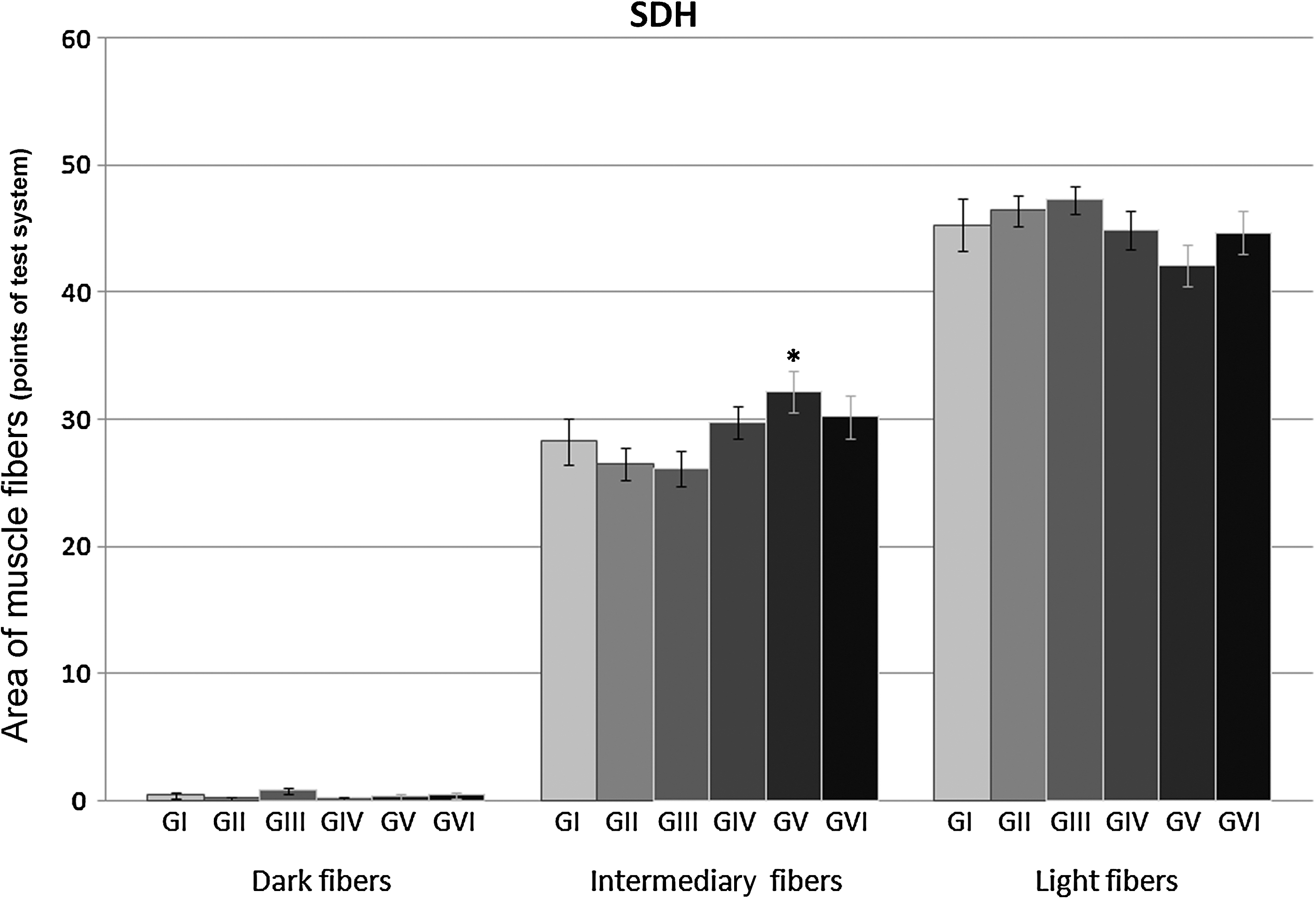
Bar graph showing areas of muscle fiber types revealed by the SDH reaction in the six groups. Only group V shows a significant increase in the intermediary fiber area, revealing an increase in oxidative activity. No conclusion could be drawn from the very small areas corresponding to dark fibers.
Figure 5 shows an increased intensity in proteinase activity bands for the groups exposed to LLLT. For MMP-2, which is expressed in the control group (group I), it only detected a significant increase in the group treated with the highest energy density (group VI); however, for MMP-9, which is not detected in the control group, it observed a slight but detectable increase in its activity in all irradiated groups (II, III, IV, and VI).
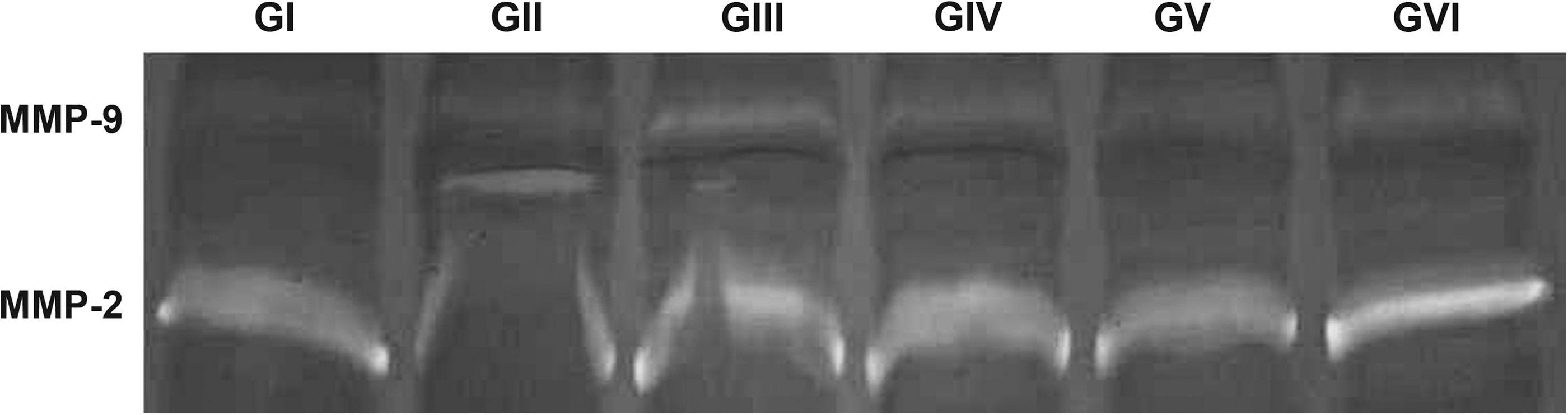
Enzymatic activity (SDS-PAGE and zymography) of extracts of masseter muscle dissected from animals irradiated by laser light in different dosages. Different patterns of qualitative activity/expression are observed among the six groups. The data are representative of separate experiments, five animals per group.
Discussion
In this investigation, Wistar rat masseter muscle was irradiated with different energy densities (0, 0.5, 1.0, 2.5, 5.0, and 20 J/cm2) of low-level laser irradiation, favoring comparison of biological effects. Muscle biomodulation was suggested when areas of light, intermediary, and dark fibers were compared among the different experimental groups. Color intensity differences in methods of enzyme activity determination detected area increase of intermediate fibers in groups irradiated with the highest energy densities, in the same proportion as the decreased areas in light fibers. Thus, the oxidative capacity of the rat masseter muscle was biomodulated by LLLT, showing a tendency to dose-dependent response up to 5 J/cm2. Similar dosages (0.6, 1.2, 2.4, and 4.8 J/cm2) were irradiated, in rat liver, increasing the NADH activity. 24 Nevertheless, in this study, the oxidative activity, in group VI (20 J/cm2), suggests that this energy density might inhibit the biomodulatory effect, when compared to group V. Other studies also report that the best results were not observed with the highest dosages in rodent skeletal muscles. 10,41,42 It was observed in cell culture that the lowest dosage used (0.5 J/cm2) was optimal to stimulate proliferation of mesenchymal stem cells in the spinal medulla. 13 The data corroborate these observations, suggesting that low dosages regulate redox in cell metabolism and in high dosages predominate the photodynamic injuries. Light would be the trigger in cell metabolism regulation, therefore explaining the best effects of low dosages and intensities. 30
The increased oxidative metabolism in the rat masseter muscle irradiated with LLLT (2.5–20 J/cm2), shown in this study, contributes to the understanding of the mechanism underlying the improved muscle fatigue induced by laser in rats 41,43 and in humans. 14,15 In contrast, it was reported that low-level laser did not improve muscle fatigue. Gorgey et al. 18 justified the lack of relevant results by the small number of individuals involved. 18 However, it is possible that the energy used (3 and 7 J) in the protocols of Gorgey et al. 18 was the limiting factor, considering that the human skin is ∼1030–1191 μm thick. 44
In this study, the NADH activity was increased in rat masseter muscle irradiated (2.5, 5.0, and 20 J/cm2). A significant increase of NADH activity was also observed in rat liver irradiated in vitro with argon-dye laser (660 nm) 24 and in yeast cultures irradiated with He-Ne laser (460 J/m2, 632.8 nm). 25 Other study suggested that laser increased the efficiency in energetic transfers by NADH in the mitochondrial respiratory chain. 28 These data corroborate our study.
The SDH activity revealed a large area of light fibers, characterized by low metabolic activity in the superficial bundle of the masseter muscle. However, a discrete area reduction of the light fibers was detected in favor of the intermediate fibers as the energy density applied increased. These data complement NADH results confirming that LLLT modifies the oxidative metabolism in skeletal muscle. There are reports that laser was able to normalize SDH activity in inflamed tissues, 26 in addition to the increasing of SDH activity and mitochondrial respiratory chain in injured muscles (GaAs, 904 nm, 5 J/cm2). 45 These observations suggest that increased muscle metabolic activity is a factor that explains the mechanism involved in laser-induced improvement of muscle fatigue. It is noteworthy to mention that there are reports of total suppression of SDH activity in mouse fibroblasts irradiated in vitro by blue light laser (5 J/cm2). 27 However, the wavelength of blue light laser is close to the range of ultraviolet irradiation, which causes health hazards because of its ionizing ability, unlike the laser used in this study (infrared, 780 nm).
The laser effects on mitochondria such as increased mitochondrial density; 9 amplification of intermembrane and cristae spaces during muscular regeneration; 10 modified microstructure of giant mitochondria in yeast, possibly reflecting a respiration and ATP synthesis activation; increased number of mitochondria and endoplasmatic reticulum associations; 25 and alterations in the potential in mitochondrial membrane, as a stimulus to ATP synthesis, both inside and out in mitochondrial matrix, 29 confirm studies by showing that laser irradiation of myotubes (0.33–1.34 J/cm2) restores mitochondrial function, 23 which may be a reason to use laser therapy in the treatment of mitochondrial dysfunction induced by physical exercise or muscular fatigue.
The present study examined whether exposure to different laser dosages result in differential expression/activity of gelatinases (MMP-2 and MMP-9) in rat masseter muscle. Corroborating reports that described that under basal conditions skeletal muscles in adult mice constitutively express MMP-2 but not MMP-9, 46 the zymography in this study showed the expression/activity only of MMP-2 in the untreated control group (group I). However, under exposure to different laser doses a slight but detectable increase in MMP-9 expression/activity after exposure to all doses used can be observed, whereas an MMP-2 increase compared to the untreated control group was detected only when the highest dosage, 20 J/cm2, was applied. This increase in the expression/activity of MMP-9 after laser irradiation was also observed in other studies. 6,35 Furthermore, different studies have demonstrated that during pathological and regenerative processes, or in response to experimentally injured muscle, MMP-2, and, especially, MMP-9, are upregulated. 47, 48
Conclusions
Considering the described results, it is evident that increased oxidative metabolism reflects the biomodulating capacity of laser energy, and that the induction of MMP-2 and MMP-9 may indicate a matrix remodeling process after exposition to laser. However, when different energy densities are applied in the same clinical conditions and do not show significant differences, it emphasizes the need for more studies to develop satisfactory protocols, which is less costly to both professional and patient. The energy density of 20 J/cm2 (group VI) did not produce the best results in relation to oxidative metabolism, suggesting that this dose was high for this protocol. In addition, the qualitative analysis of MMPs in this study helped to show the importance of quantifying MMPs in the masseter muscle in future studies.
Footnotes
Acknowledgments
We are grateful to FAPESP for financial support for this research.
Author Disclosure Statement
No conflicting financial interests exist.