
Abstract
Introduction
Human fibroblasts exposed to 8-methoxypsoralen plus ultraviolet-A irradiation (PUVA) may show stress-induced senescence. 10 Therefore, PUVA is a suitable tool for the analysis of senescence-related mechanisms in vitro. Cell senescence may show various robust markers, 11 including morphological changes, increased staining of senescence-associated β-galactosidase (SA-β-gal), 12 telomere shortening, 13 and induction of reactive oxygen species (ROS). 14
This study was designed to evaluate whether IPL exposure attenuates PUVA-induced cell senescence and to explore the mechanism of IPL-induced skin rejuvenation.
Methods
Cell culture
Fibroblasts were harvested from the foreskin of human donors 6 years of age, and were grown at 37°C in 5% CO2 and 95% air, in Dulbecco's Modified Eagle's Medium (DMEM; GIBCO-BRL, Eggenstein, Germany) supplemented with 10% fetal calf serum (FCS; Biochrom AG Biotechnologie, Berlin, Germany).
PUVA treatment
Crystalline 8-methoxypsoralen (8-MOP; Sigma-Aldrich, Steinheim, Germany) was dissolved in dimethyl sulfoxide (DMSO) at a concentration of 1 mg/ml. Based on preliminary tests and the literature, 15 50 ng/ml of 8-MOP was added to the medium, and the cells were cultured for 16–18 h. Before UVA irradiation, the culture medium was changed to pre-warmed phosphate buffered saline (PBS) containing the same concentration of 8-MOP. The cells were irradiated using a high-intensity UVA lamp at wavelengths of 340–400 nm (UVA1-S; Shanghai SIGMA High-Tech Co. Ltd., Shanghai, China).
IPL irradiation
After treatment, both PUVA-treated fibroblasts and normal fibroblasts were immediately exposed to IPL once a day for 5 days, and the medium was changed every day. Cells were treated with the IPL device (Lovely II Laser AFT Workstation; Lumenis Ltd., Yokneam, Israel) using a 570-nm cutoff filter with a pulse interval of 12 ms and fluences of 15 J/cm2.
Senescence-associated β-galactosidase staining
Five days after PUVA, PUVA+IPL and IPL treatment, the fibroblasts were trypsinized and seeded in 35-mm culture dishes (Corning, Lowell, MA). 24 h after the seeding, SA-β-gal staining was performed according to the manufacturer's instructions (Senescence-Associated β-Galactosidase Staining Kit; Cell Signaling Technology, Danvers, MA). At least three independent experiments were performed. The stained cells were photographed, positive fibroblasts were counted, and the results were expressed as mean±SD of the percentage of SA-β-gal-positive fibroblasts in the total number of fibroblasts.
MTT cell viability
On the first, third, and fifth days after PUVA, PUVA+IPL, and IPL treatment, viability of the fibroblasts was monitored using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT; Sigma, China) to quantify living and metabolically active fibroblasts. Cell viability was assessed using a tetrazolium dye that is reduced to a blue formazan only by living cells. The amount of dye formed was quantified using a microplate spectrophotometer (ELISA plate reader). The amount of formazan dye produced was confirmed by visual observation to be directly proportional to the number of viable target cells. The cell viability was calculated as the percentage absorbance relative to control cultures.
Real-time polymerase chain reaction (PCR) for telomere length
Genomic DNA was extracted directly from cell samples via standard procedures. Fibroblast telomere length was measured by quantitative PCR using the method described by Cawthon. 16 Briefly, for each sample, two PCRs were performed: the first one was to amplify the telomeric DNA (T), and the second one was to amplify a single-copy control gene(S) (36B4, acidic ribosomal phosphoprotein P0). The amount of telomeric DNA was divided by the amount of control-gene DNA, producing a relative measurement of the telomere length of the sample. All PCRs were performed on the Prism 7700 Sequence Detection System (ABI; Applied Biosytems, Inc., Foster City, CA). ABI SDS v.1.7 software was then used to generate the standard curve for each plate and to determine the dilution factors of standards corresponding to the T and S amounts in each sample.
Measurement of intracellular ROS with flow cytometry
The fluorescent probe 2′,7′-dichlorodihydrofluorescein diacetate (DCFH2-DA; Image-iT ™ LIVE Green Reactive Oxygen Species Detection Kit I36007, Invitrogen Ltd., Paisley, UK) was used to assess intracellular oxidative stress. Briefly, on the sixth day after the PUVA and IPL treatments, the fibroblasts were trypsinized and collected in 1 ml of PBS, incubated with 5 μg/ml of DCFH2-DA for 30 min at 37°C, and then washed three times with PBS. This process was followed by flow cytometric analysis (FACScan; Becton, Dickinson, and Company, Milan, Italy) using Cell Quest 3.2 software (Becton, Dickinson, and Company). The results of the flow cytometry experiments were expressed as the mean of fluorescence intensity.
Statistical analysis
The results are expressed as mean±SD from at least three independent experiments. Statistical analysis was performed using the Statistical Package for Social Sciences (SPSS) version 13.0 (SPSS Inc., Chicago, IL). One-way and two-way analyses of variance (ANOVA) were used to determine the statistical significance of differences. Probability values <0.05 were considered significant.
Results
PUVA increased SA-β-gal activity and IPL decreased PUVA-induced SA-β-gal activity
SA-β-gal activity was detected 1 day after PUVA treatment, and a steady increase in the numbers of SA-β-gal-positive fibroblasts was observed over a period of 5 days (data not shown). Five days after PUVA treatment, 81% of the fibroblasts showed de novo expression of SA-β-gal (Fig. 1). Five days after treatment, the SA-β-gal-positive cell percentages in the PUVA+IPL group, IPL group, and control group were 72%, 10%, and 11%, respectively. After PUVA treatment, IPL reduced the percentage of SA-β-gal-positive fibroblasts (p<0.05). IPL irradiation alone without earlier PUVA exposure did not affect the activity of SA-β-gal (p>0.05 in comparison with the control group) (Fig. 2).
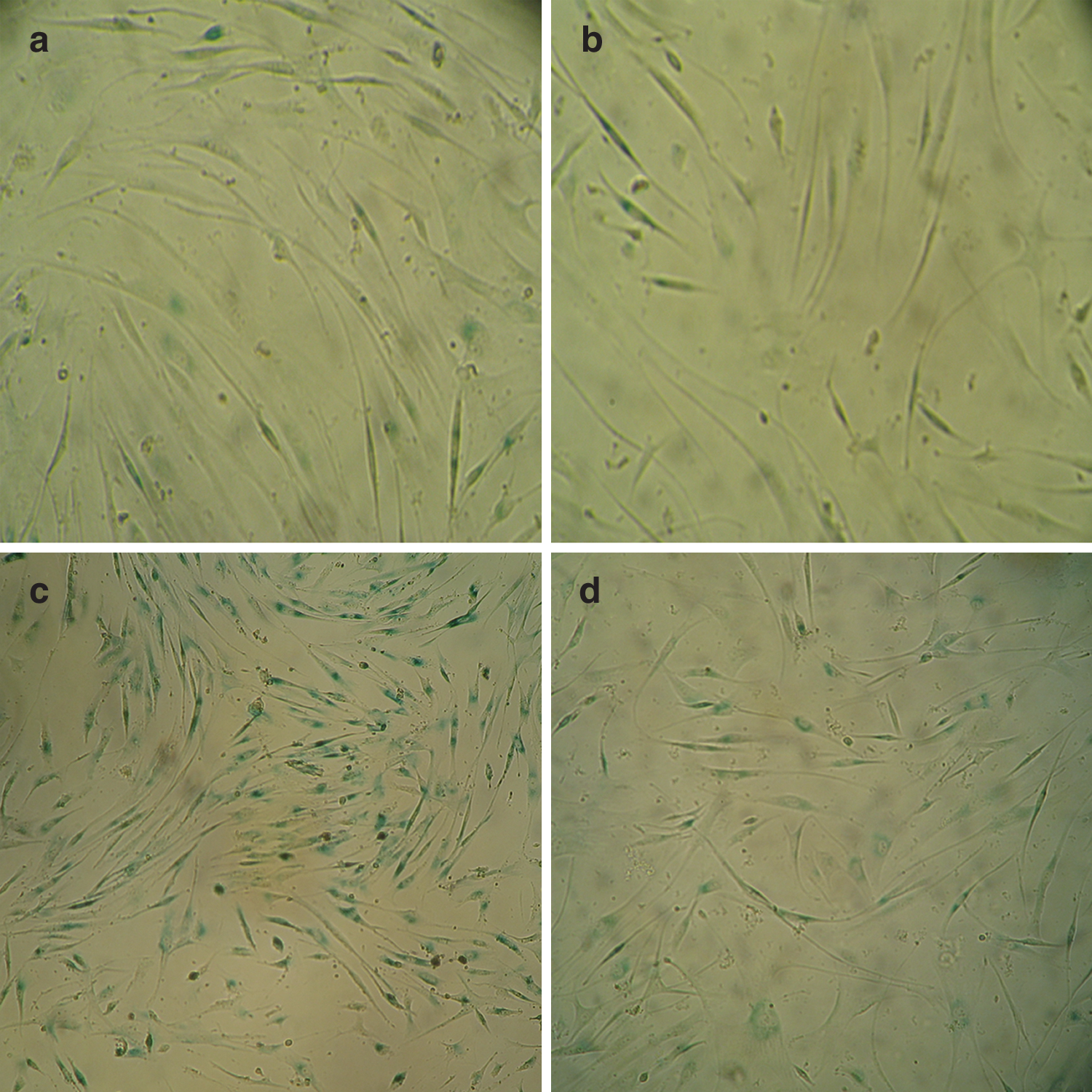
PUVA treatment induced the expression of SA-β-gal.

IPL decreased PUVA-induced SA-β-gal activity. SA-β-gal expression was detected as described in the material and methods. The number of SA-β-gal-positive fibroblasts is shown as mean±SD of at least three independent experiments performed in triplicate. Supplementation with IPL reduced the percentage of SA-β-gal-positive fibroblasts, in comparison with the percentage in the PUVA group (p<0.05). Five consecutive days of IPL irradiation alone did not increase the activity of SA-β-gal, in comparison with the control cells (p>0.05). *p<0.05 as compared with the control group; #p<0.05 as compared with the PUVA group.
IPL treatment did not increase the viability of fibroblasts with PUVA-induced senescence
PUVA induced a marked time-dependent decrease in cell viability; the cell viability diminution was 90% on the first day, 77% on the third day, and 40% on the fifth day. This indicates that PUVA induced a long-term arrest of growth. However, supplementary IPL exposure did not immediately improve cell viability (p>0.05). IPL irradiation alone without earlier PUVA exposure dramatically increased cell viability; the viability increased by 182% on the first day, 188% on the third day, and 194% on the fifth day (Fig. 3).
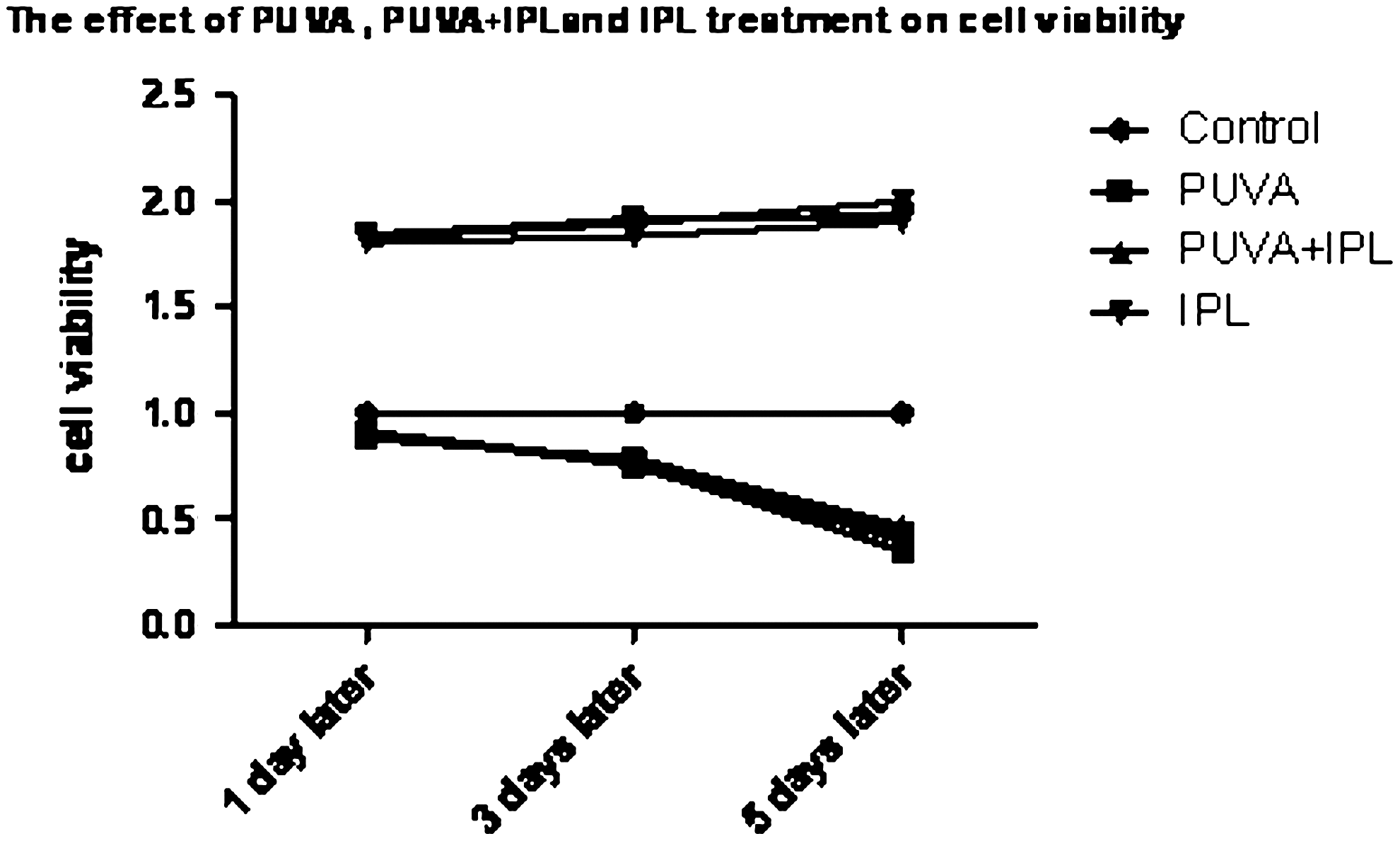
IPL treatment did not increase the viability of fibroblasts with PUVA-induced senescence. Viability was calculated as the percentage of light absorbance relative to the control group and is shown as mean±SD of at least three independent experiments performed in triplicate. PUVA induced a marked time-dependent decrease in cell viability, however, supplementary IPL exposure did not immediately improve cell viability (p>0.05). IPL irradiation alone without earlier PUVA exposure dramatically increased cell viability as compared with control (p<0.05).
IPL provided protection against PUVA-induced shortening of telomere length
The telomeres of cells in the PUVA treatment group were clearly shorter than those in the control group (p<0.05). Immediate supplementation with IPL irradiation protected fibroblasts against PUVA-induced shortening of telomere length (p<0.05). Five consecutive days of IPL irradiation alone had no effect on telomere length, in comparison with the length in the control group (p>0.05) (Fig. 4).
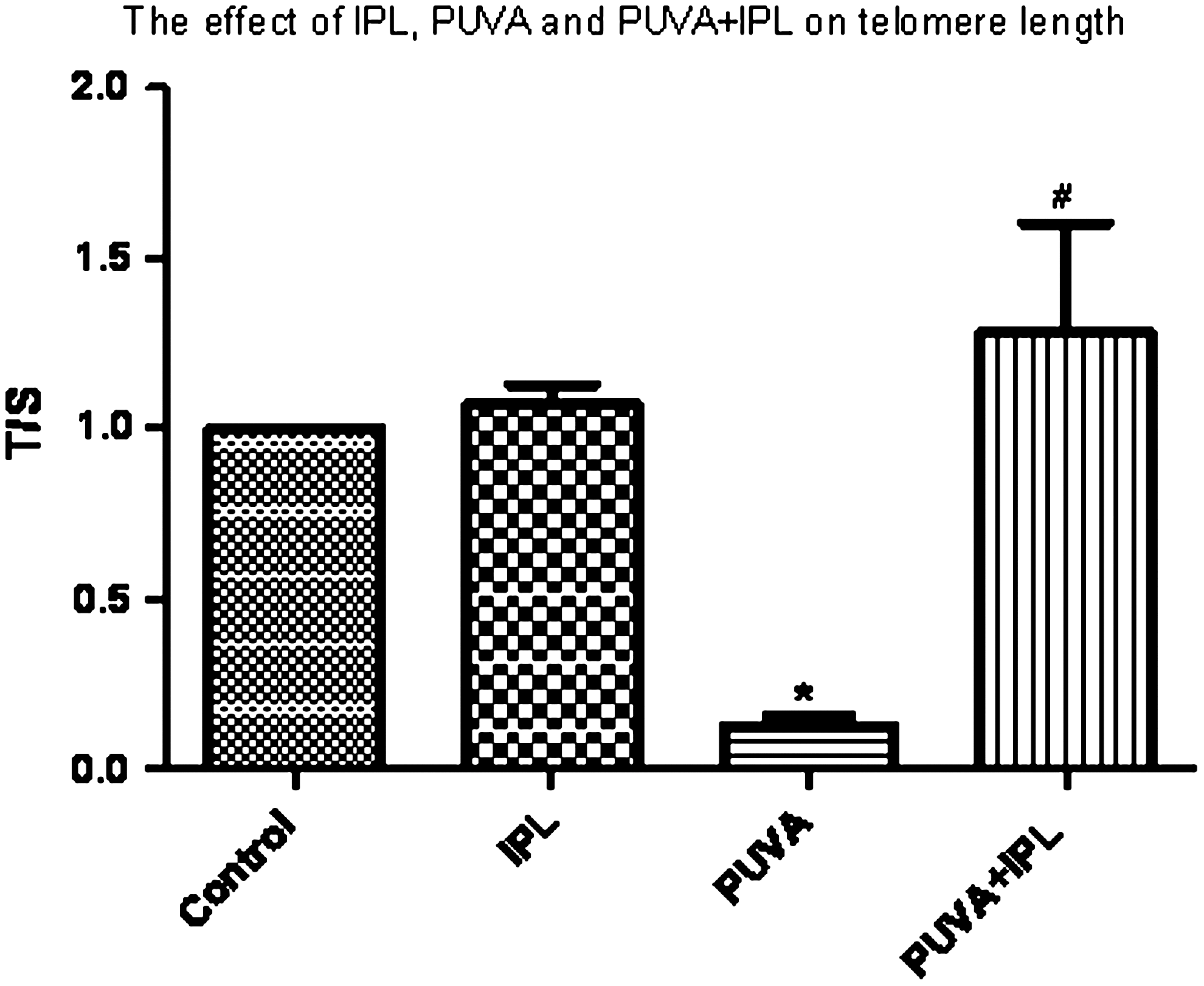
IPL provided protection against PUVA-induced shortening of telomere length. Our strategy for determining relative telomere lengths by quantitative PCR was to measure, for each DNA sample, the factor by which the sample differed from a reference DNA sample in its ratio of telomere repeat copy number (T) to single copy gene copy number (S). This ratio should be proportional to the average telomere length. PUVA treatment shortened telomere length compared to control (p<0.05), whereas immediate supplementation with IPL irradiation protected the fibroblasts against the PUVA-induced telomere shortening (p<0.05). Five consecutive days of IPL irradiation alone had no effect on telomere length in comparison with the control (p>0.05). *p<0.05 in comparison with the control group; #p<0.05 in comparison with the PUVA group.
IPL exposure protected fibroblasts against PUVA-induced release of ROS
ROS production was detected 5 days later after exposure of fibroblasts to PUVA, PUVA+IPL, and IPL. PUVA-treated fibroblasts had high levels of ROS, whereas the PUVA-induced ROS production in cells exposed to IPL was significantly lower than that in the PUVA group (p<0.05). Five consecutive days of IPL irradiation alone decreased the level of ROS in comparison with the levels in the control group (p<0.05) (Fig. 5).
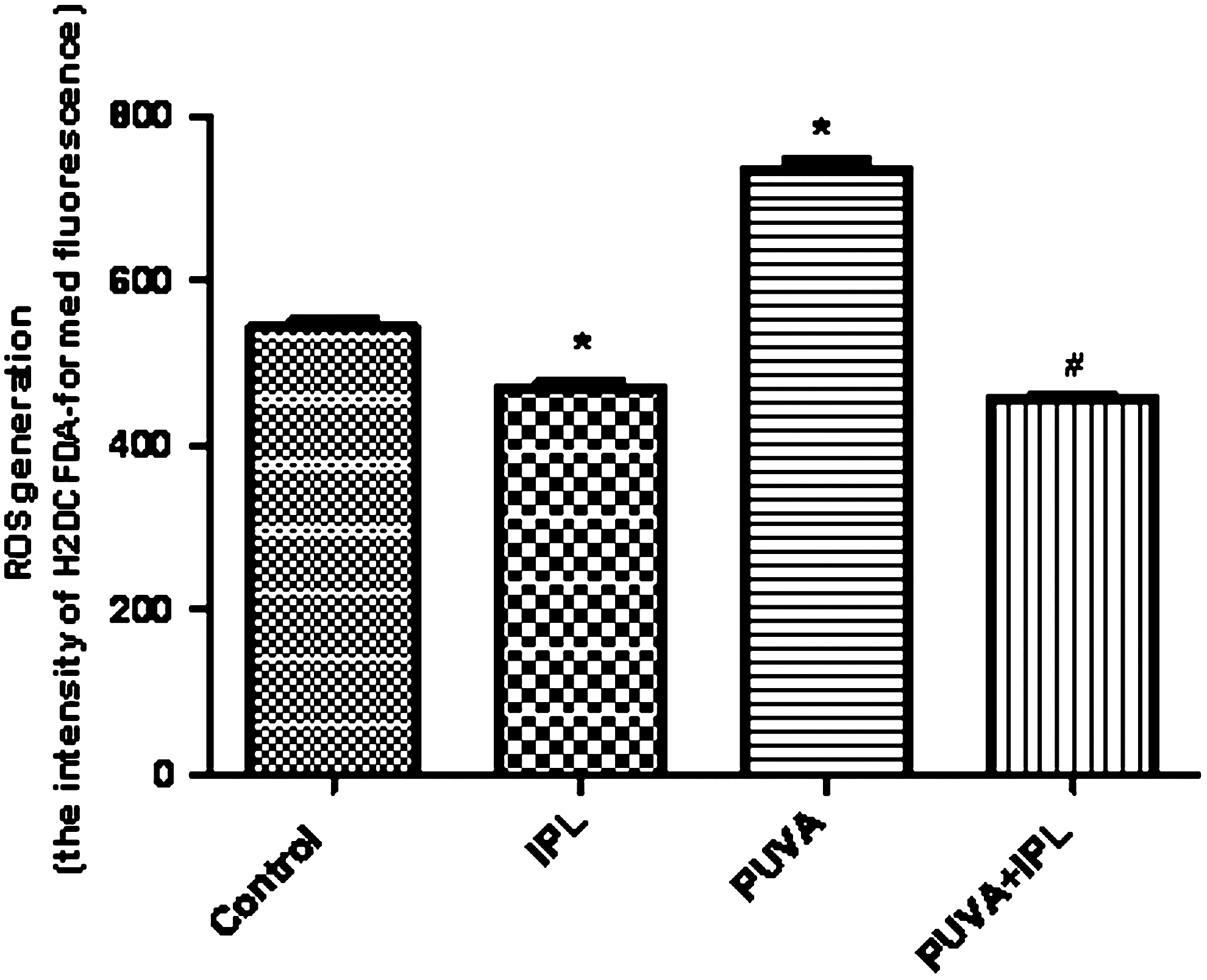
IPL exposure protected fibroblasts against PUVA-induced release of ROS. Intracellular ROS levels were assessed by the intensity of H2DCFDA-fluorescence as determined by FACS analysis. ROS generation was measured 5 days after exposure of the fibroblasts to IPL, PUVA, and PUVA+IPL. PUVA induced ROS production, in comparison with the findings for the control group (p<0.05), whereas PUVA+IPL significantly reduced ROS level, in comparison with the findings for the PUVA group (p<0.05). Results are expressed as means±SD of at least three independent experiments performed in triplicate. *p<0.05 as compared with the control fibroblast; #p<0.05 as compared with the PUVA-treated fibroblast.
Discussion
Senescence-associated β-galactosidase (SA-β-gal) activity is widely used as a marker of cellular senescence. 17,18 Briganti et al. 15 found that the SA-β-gal activity increased after PUVA treatment. In our study, we examined SA-β-gal activity after IPL, PUVA and PUVA+IPL irradiation. The results showed that IPL exposure alone has no effect on the activity of SA-β-gal which showed that multiple IPL irradiations may not result in cell senescence. Single PUVA treatment sharply increases the activity of SA-β-gal; however, supplementary IPL exposure abrogates the PUVA-induced increase of SA-β-gal activity. This suggests that IPL may partially rescue the cellular senescence phenotype. To our knowledge, this may be the first report in the literature about the effects of IPL.
Previous studies and our data showed that IPL could increase fibroblast activity 2 –4, 6 –9 in both normal tissue and cultured cells. To clarify the mechanism of the IPL-mediated protection against PUVA-induced senescence in fibroblasts, an MTT assay was performed to evaluate cell viability in our experimental system. The data showed that PUVA induced a time-dependent decrease in cell viability, whereas supplementation with IPL after PUVA treatment did not improve the viability of cultured cells. This result indicated that IPL irradiation may have different effects on normal cells and senescent ones, promoting proliferation in the former but having little effect on the latter.
It is generally accepted that oxidative stress and ROS eventually cause DNA damage, whereby insufficient cellular repair mechanisms may contribute to premature aging. ROS resulted in cell senescence, 19 which could be induced by PUVA. 15 IPL generates high-intensity short flashes of polychromatic light in the visible and near-infrared range (500–1200 nm), depending upon the filters used. Sorg et al. 20 thought that IPL, although filtered for wavelengths <500 nm, could generate oxidative stress in human skin in vivo. However, Applegate et al. 21 showed that infrared light (700–4000 nm) was not capable of inducing frank damage to DNA or oxidative stress proteins in fibroblasts in vitro. Our results showed that IPL exposure generated no reactive oxidative stress in this culture system, which are not consistent with the results of Sorg et al. 20 but agree with those of Applegate et al. 21 This indicated that the small infrared component of our IPL system (570–950 nm) was probably not responsible for the oxidative stress in vitro. This oxidative stress in skin biopsy specimens in the study of Sorg et al. 20 was, therefore, probably caused by a moderate dose of visible light delivered through a very high fluence rate.
The mechanism whereby IPL irradiation alone increases cell viability and decreases ROS levels remains unclear. We assume that IPL generates short flashes of visible light, which are delivered in a short pulse at a moderate dose. This mode of action differs from that of UVA, and therefore results in a protective effect benefiting the treatment of photo-aging.
Telomeres play a key role in the maintenance of chromosome integrity. Short telomeres are linked to aging-associated diseases. The telomeres shortened by 455 bp over 10 years, on average. 22 Telomere shortening has been implicated in cellular senescence, which may cause certain aging phenotypes. PUVA-treated fibroblasts showed a continuing lack of telomerase activity and accelerated telomere shortening. 23 Our results showed that telomere length of cultured fibroblast cells clearly shortened upon PUVA exposure, whereas supplementation with IPL preserved telomere length quite well. These results again indicated the beneficial effects of IPL on cell senescence. Cattan 24 reported that increased oxidative stress in vivo promotes telomere shortening in CAST/Ei mice. Our data support the idea that IPL decreases the level of PUVA-induced ROS and consequently prevents telomere shortening.
In addition, telomere maintenance is mainly achieved by telomerase, an RNA-dependent DNA polymerase that adds telomeric DNA to telomeres. The modulation of telomerase activity may therefore have important implications in antiaging. Studies showed that nicotine 25 and estrogen 26 reduced endothelial progenitor cell senescence through augmentation of telomerase activity. However, the relationship between IPL irradiation and telomerase activity has not been reported so far, and further study is needed.
Conclusions
PUVA treatment induced cell senescence in cultured fibroblasts, whereas IPL exposure protected cells against PUVA-induced aging by reducing ROS and preventing telomere shortening.
Footnotes
Acknowledgments
This study was funded by the National Natural Science Foundation of China (30872269).
Author Disclosure Statement
No conflicting financial interests exist.